锰、铜、锌和铁离子对抑食金球藻生长的影响
2019-05-30石伟杰李欣阳王全颖张永丰杜雨蒙
石伟杰,李欣阳,张 勇,王全颖,张永丰,李 莉,杜雨蒙,杨 超
(国家海洋局 秦皇岛海洋环境监测中心站,河北 秦皇岛 066002 )
近年来,秦皇岛近岸海域由微微型浮游植物引起的褐潮频发。该海域褐潮具有细胞密度极高,暴发面积大,持续时间较长等特点。Zhang等[1]通过对优势种18S rDNA序列监测分析,确定褐潮藻种为抑食金球藻(Aureococcusanophagefferens)。我国是第三个出现抑食金球藻褐潮的国家。褐潮发生期间,海域水体透明度降低,抑食金球藻能适应低光照环境,细胞迅速繁殖,底栖藻类光合作用却受到抑制,严重危害了海域生态系统;另一方面褐潮不仅对扇贝、鱼类及其他海洋生物的重要栖息造成严重的破坏,还能抑制滤食贝类的进食,从而导致其生长受阻,对海域养殖业造成巨大的经济损失[2-4]。随着人们对海洋生态环境的日益重视,抑食金球藻褐潮已成为研究的热点。国内外对抑食金球藻研究主要集中于其生理生化方面及其对生态环境的影响[2-7],关于重金属对抑食金球藻生长影响的报道较少。铁、锰、锌和铜等是秦皇岛海域海水中常见重金属离子,同时也是藻细胞生长过程中必不可少的元素,其中有学者认为铁和锰与赤潮发生有一定的关系[8-9]。本试验通过探讨锰、铜、锌和铁4种重金属离子对抑食金球藻生长综合效应的影响,以期为秦皇岛海域抑食金球藻引发褐潮的研究提供科学依据。
1 材料与方法
1.1 试验藻种
抑食金球藻来自国家海洋环境监测中心,保存于L1培养基中[10],光照条件4000 lx,光暗周期为12L∶12D,培养温度为(25±1) ℃。
1.2 试验方法
分别用分析纯的CuSO4、MnCl2和ZnSO4溶解于去离子水中,配制成0.1 g/L的重金属母液;用FeCl3配制成2.0 g/L的母液。试验时,用消毒的海水将重金属母液稀释至试验所需的离子质量浓度。
1.2.1 4种重金属离子对抑食金球藻生长的影响试验
试验在250 mL的三角烧瓶中进行,CuSO4据其质量浓度设为0(对照组)、5、25、125、625 μg/L试验组,每组设有3个平行,其他营养盐均采用L1培养基[10]。MnCl2和ZnSO4试验也设置同样的质量浓度梯度,按上述方法进行。FeCl3据其质量浓度设为0(对照组),100、500、2500、12 500、62 500 μg/L试验组,也按上述同样的方法进行试验。将处于指数生长期的藻种接种到上述各试验组中培养,培养液最终体积为200 mL,起始藻种密度为1.10×104个/mL,培养条件同1.1,每日定时摇瓶3次。在指数生长期末期,测定各试验组细胞密度、相对生长率和叶绿素含量。
1.2.2 重金属离子对抑食金球藻生长的综合效应试验
通过1.2.1试验筛选出对抑食金球藻生长影响较大的3种重金属离子,选取每种重金属离子3个合适的质量浓度,设计正交试验,除以上3种重金属离子质量浓度外,其他营养盐均采用L1培养基[10],培养条件同1.1。试验组培养液最终体积为200 mL,起始藻种密度为1.80×104个/mL,每日定时摇瓶3次。正交试验进行7 d,试验结束后测定各试验组细胞密度、相对生长率、叶绿素含量和3种金属离子质量浓度。
1.3 参数测定
叶绿素含量测定的方法为过滤10 mL藻液后放干燥冷藏箱保存,采用分光光度计法进行分析,即以丙酮溶液提取藻细胞色素,依次在664、647、630 nm波长下测定吸光值,按Jeffrey-Humphrey的方程式计算叶绿素a的质量浓度[11],以μg/L表示。
藻细胞密度计数采用分光光度法测定。将培养好的藻按比例稀释成一定密度组,每组用计数框测定细胞密度3次,取平均值。另将上述相应的密度组用分光光度计(UV Power)测定其吸光度。将吸光度和细胞密度进行拟合,求得回归方程y=0.000738x+0.0177(r2=0.9987)[y为吸光度,x为藻细胞密度(104个/mL)]。测定每个试验组藻细胞吸光度,代入方程可得藻细胞密度。
藻液中的锰、铜、锌和铁4种重金属离子按《海洋行业规范》(HY/T 147.1—2013)[12]中的分析方法进行。藻细胞吸收重金属量按下式计算:
重金属吸收量/μg=V×(Ca-Cb)
式中,V为培养液的体积(L),Ca为初始加入重金属的质量浓度(μg/L),Cb为试验结束后测得重金属的质量浓度(μg/L)。
每日定时取样并测定藻细胞密度,按下式计算相对生长率:
相对生长率=(lnNt-lnN0)/t
式中,N0为初始藻细胞密度(个/mL),Nt为t日后藻细胞密度(个/mL),t为培养时间(d)。
1.4 统计分析
采用SPSS 11.5软件对试验数据进行单因子方差分析、多重比较(P<0.05表示显著),并对抑食金球藻细胞密度、相对生长率和叶绿素含量与重金属离子吸收量之间进行相关性分析。用Origin Pro 8.0软件作图。
2 结果与分析
2.1 4种重金属离子对抑食金球藻的生长影响
不同锰离子质量浓度对抑食金球藻细胞密度、相对生长率和叶绿素含量的影响见图1。单因子方差分析结果表明,锰离子对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度有显著影响(P<0.05)。当锰离子质量浓度为0~25 μg/L时,上述参数随质量浓度的升高呈现上升的趋势,当锰离子质量浓度大于25 μg/L时,抑食金球藻细胞密度、相对生长率和叶绿素质量浓度均呈现下降趋势。多重比较结果显示,25 μg/L组细胞密度、相对生长率和叶绿素质量浓度显著高于其他组,分别为2.52×106个/mL、0.776和10.6 μg/L。0 μg/L试验组上述参数显著低于其他试验组,分别为1.58×106个/mL、0.710和5.58 μg/L。因此,抑食金球藻生长最适宜的锰离子质量浓度为25 μg/L。
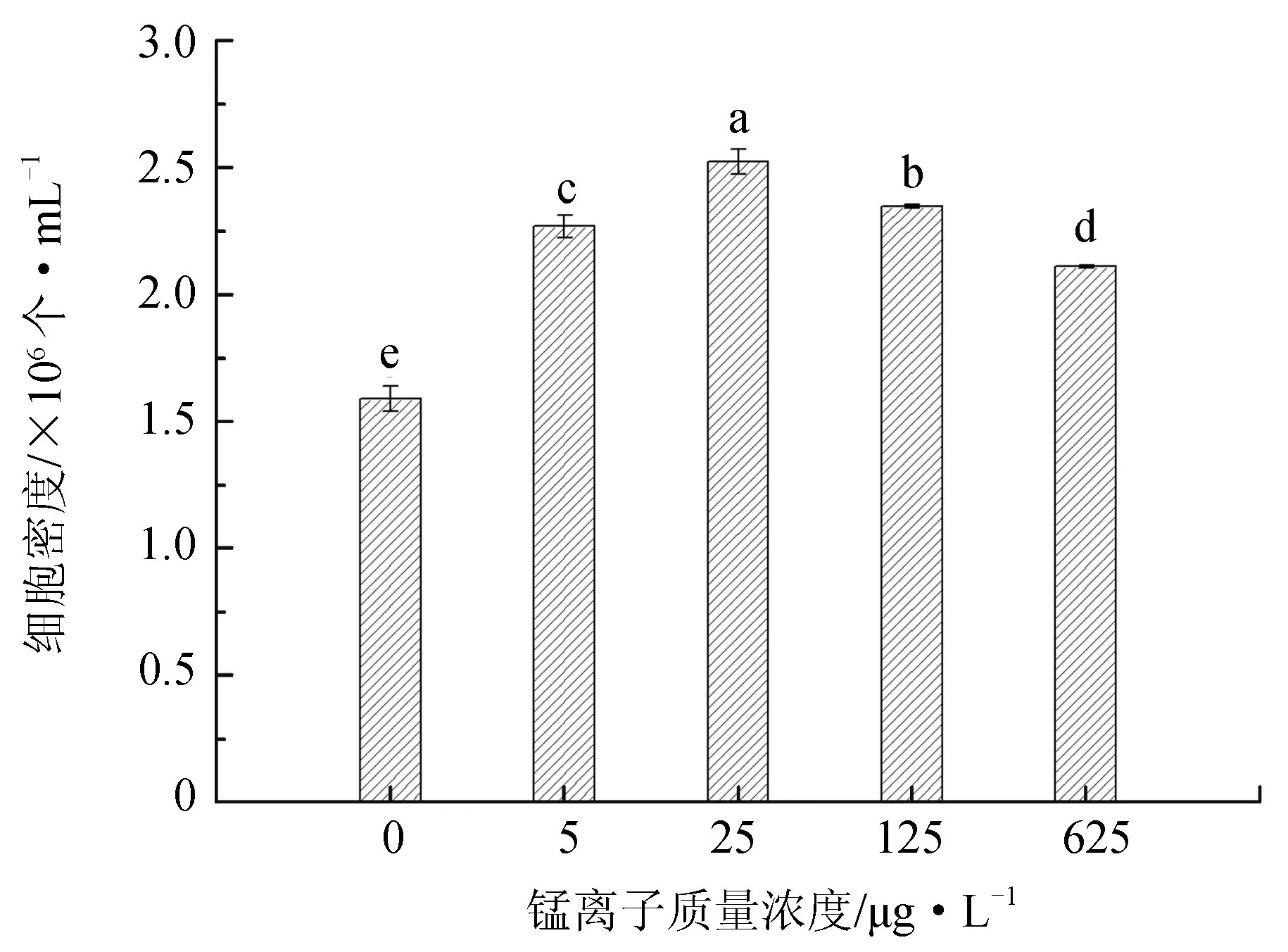
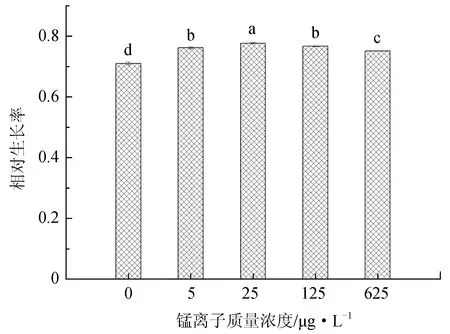
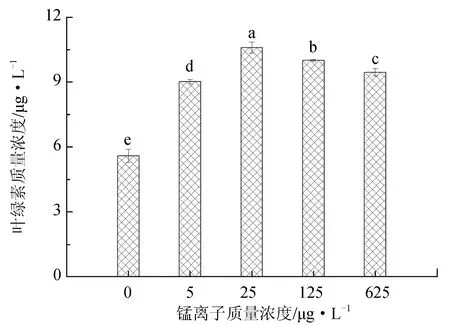
图1 不同锰离子质量浓度对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度的影响
不同铜离子质量浓度对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度的影响见图2。单因子方差分析结果表明,铜离子对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度有显著影响(P<0.05)。多重比较结果显示,5 μg/L试验组细胞密度、相对生长率和叶绿素质量浓度显著高于其他试验组,分别为2.41×106个/mL、0.770和12.5 μg/L。125 μg/L和625 μg/L试验组细胞密度和相对生长率显著低于其他组,两组之间差异不显著(P>0.05)。625 μg/L试验组叶绿素质量浓度最低,仅为7.01 μg/L。因此,抑食金球藻生长最适宜的铜离子质量浓度为5 μg/L。
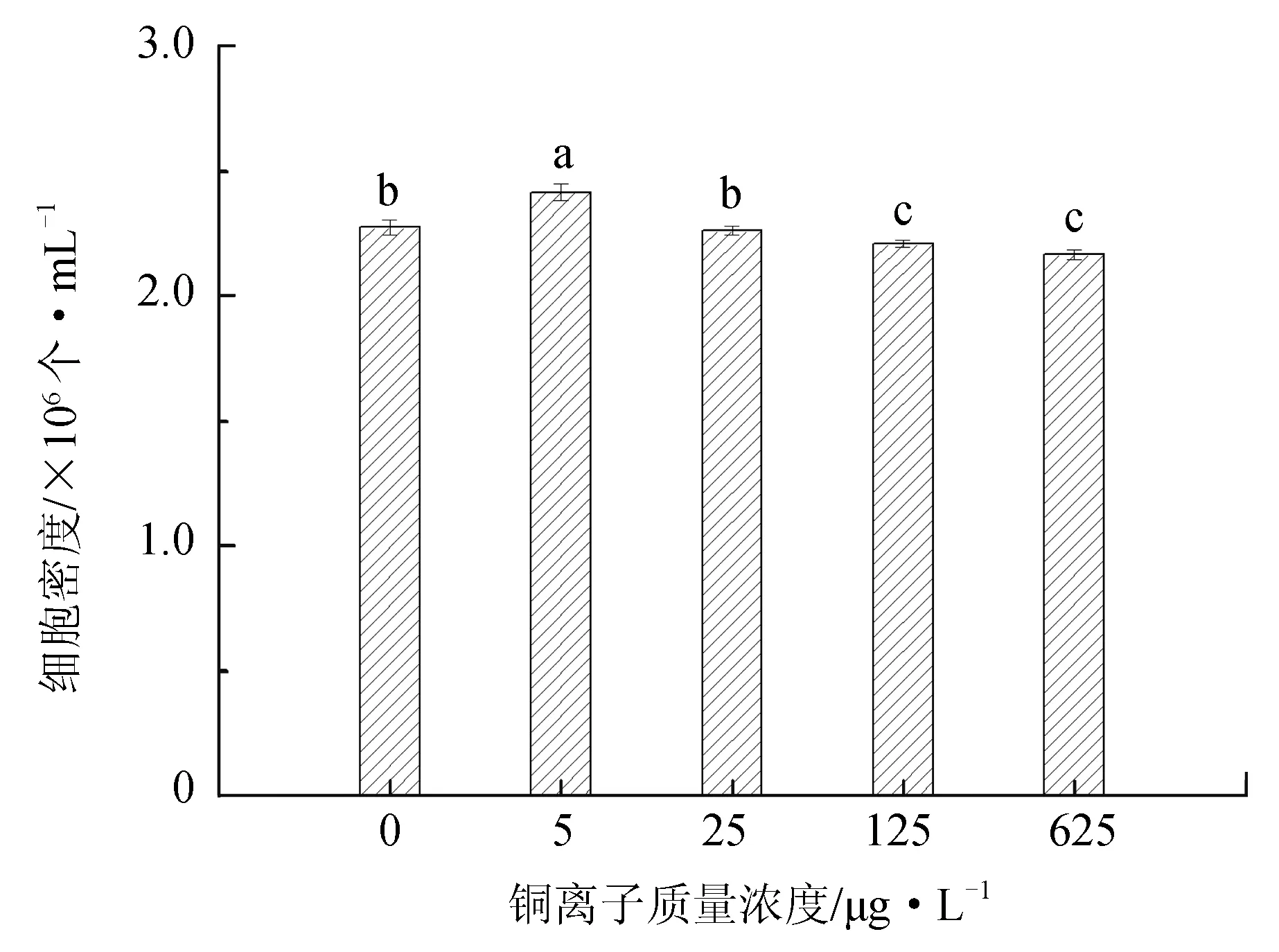
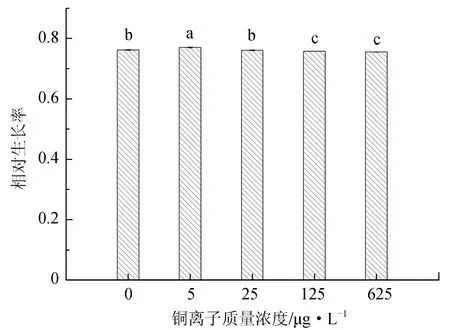
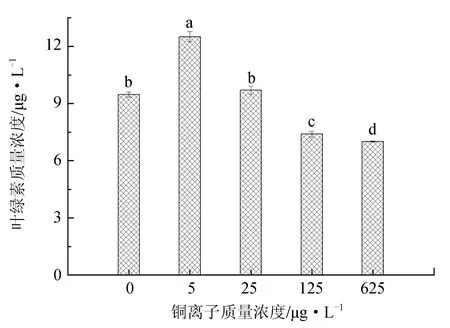
图2 不同铜离子质量浓度对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度的影响
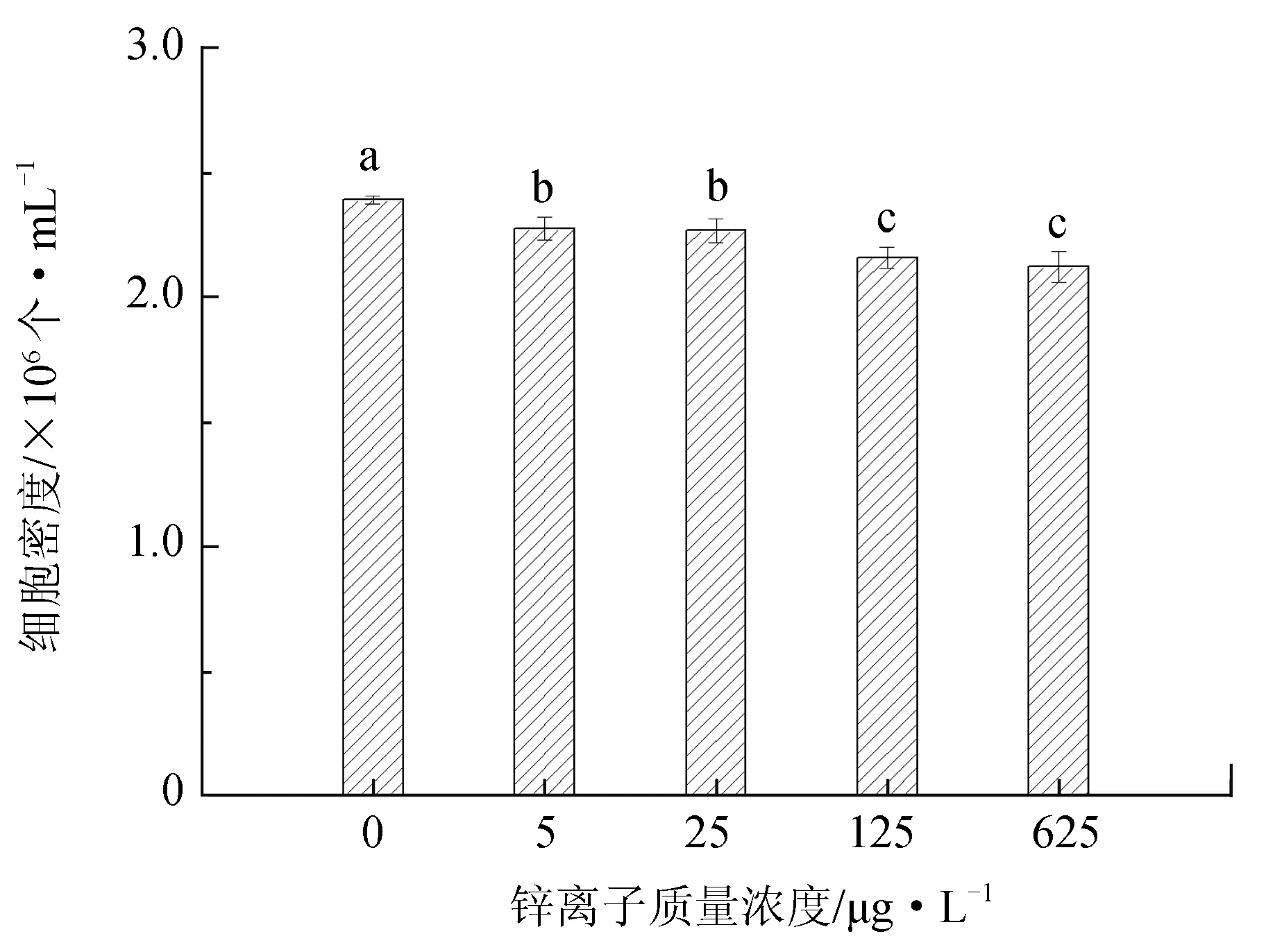
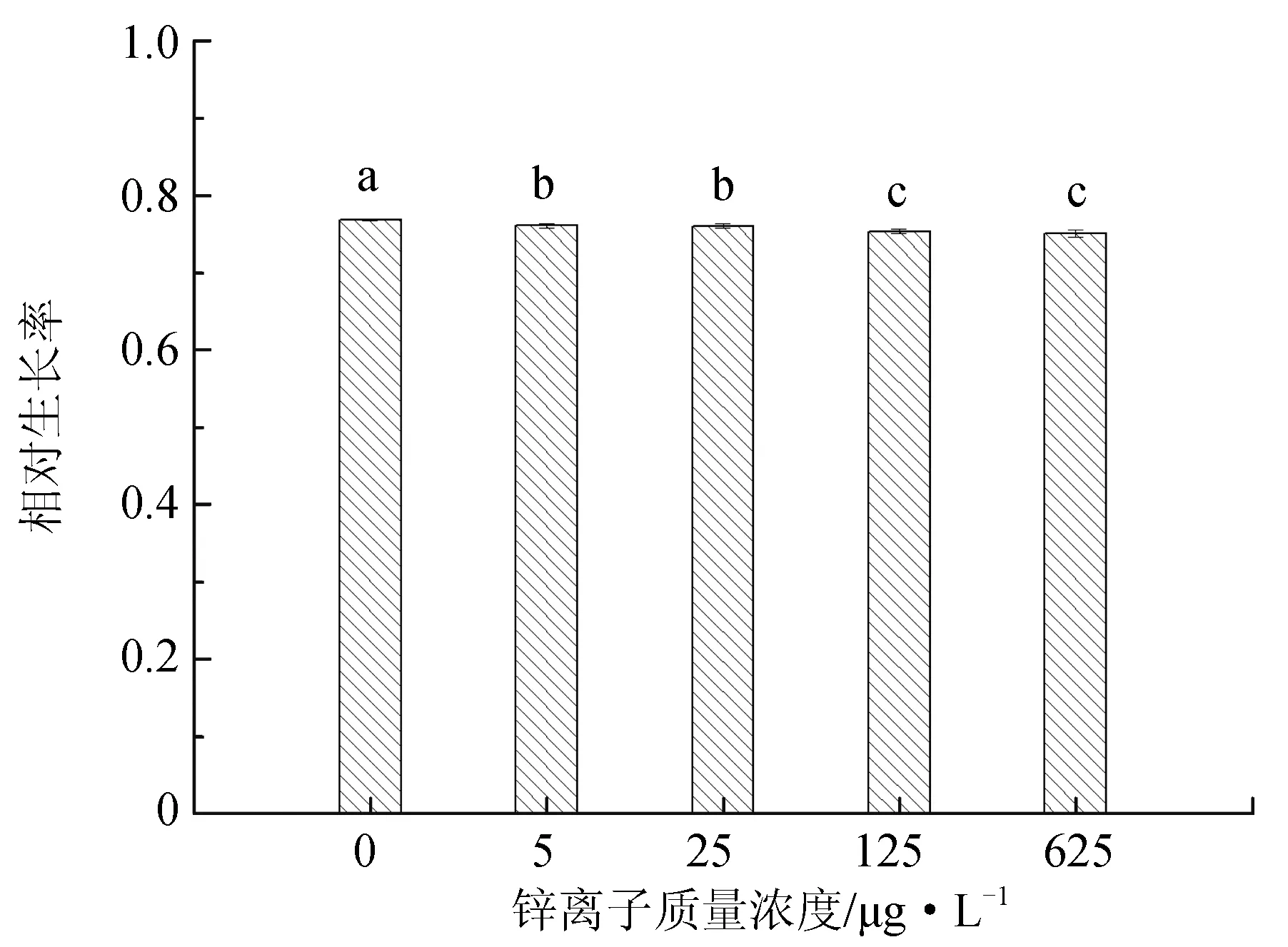
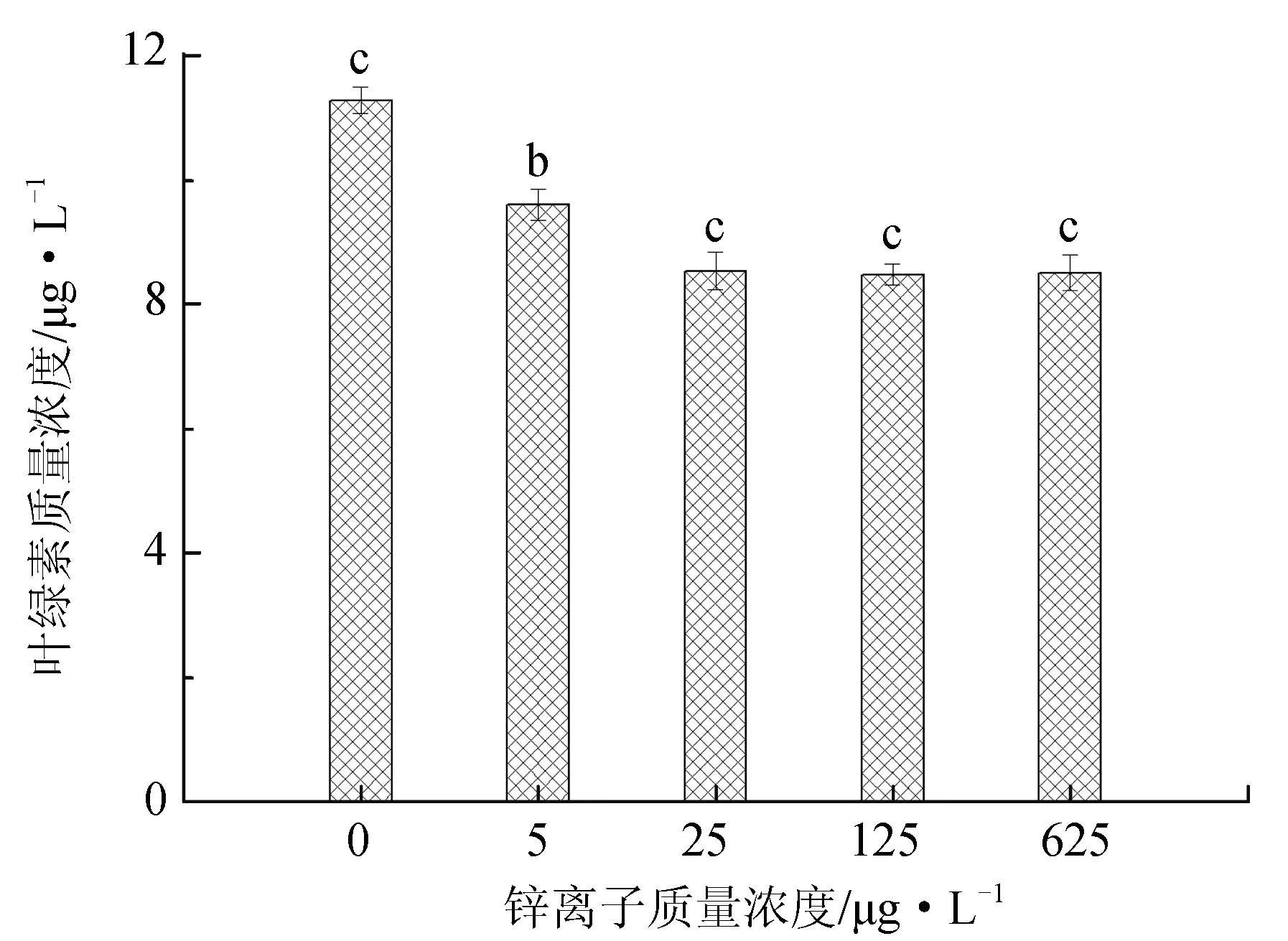
图3 不同锌离子质量浓度对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度的影响
不同锌离子质量浓度对抑食金球藻细胞密度、相对生长率和叶绿素含量的影响见图3。单因子方差分析结果表明,锌离子对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度有显著影响(P<0.05)。多重比较结果显示,0 μg/L对照组细胞密度、相对生长率和叶绿素质量浓度显著高于其他试验组,分别为2.38×106个/mL、0.768和11.3 μg/L。125 μg/L 和625 μg/L试验组细胞密度和相对生长率显著低于其他组,两组之间差异不显著(P>0.05)。25、125 μg/L 和625 μg/L试验组叶绿素质量浓度显著低于其他两组,分别为8.54、8.48 μg/L 和8.51 μg/L,3组之间差异不显著(P>0.05)。除对照组外,锌离子其他试验组对抑食金球藻生长产生抑制作用,因此不将锌离子作为后续综合效应试验的因子。
不同铁离子质量浓度对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度的影响见图4。单因子方差分析结果表明,铁离子对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度有显著影响(P<0.05)。多重比较结果显示,12 500 μg/L和62 500 μg/L试验组细胞密度、相对生长率和叶绿素质量浓度显著高于其他组,但两者之间上述参数差异均不显著(P>0.05)。对照组的细胞密度、相对生长率和叶绿素质量浓度最低,分别为7.21×104个/mL、0.268和0.179 μg/L。考虑到62 500 μg/L质量浓度较高,因此,对抑食金球藻生长最适宜的铁离子质量浓度为12 500 μg/L。
2.2 重金属离子对抑食金球藻生长综合效应
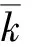
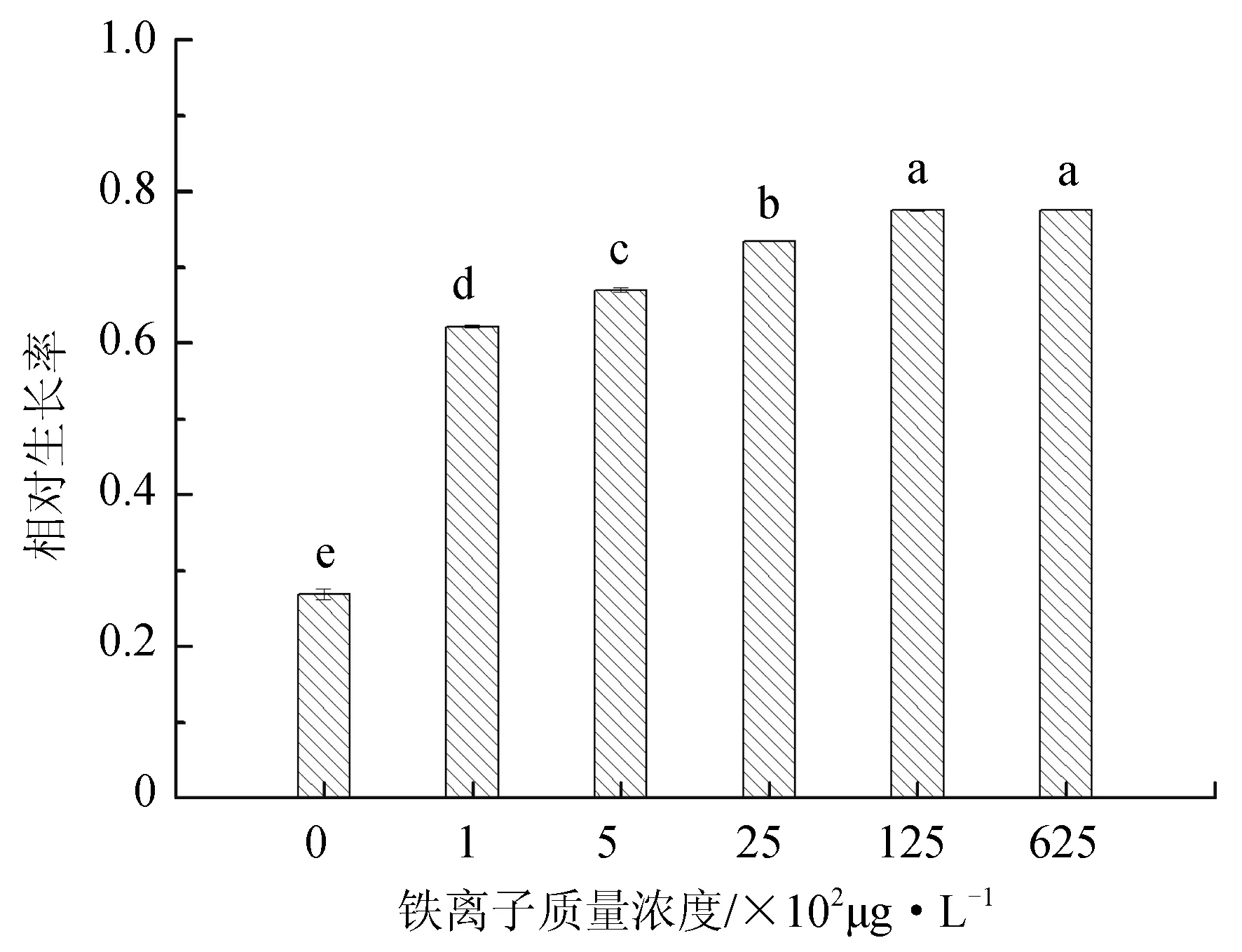

图4 不同铁离子质量浓度对抑食金球藻细胞密度、相对生长率和叶绿素质量浓度的影响

表1 正交试验重金属离子质量浓度设置 μg/L
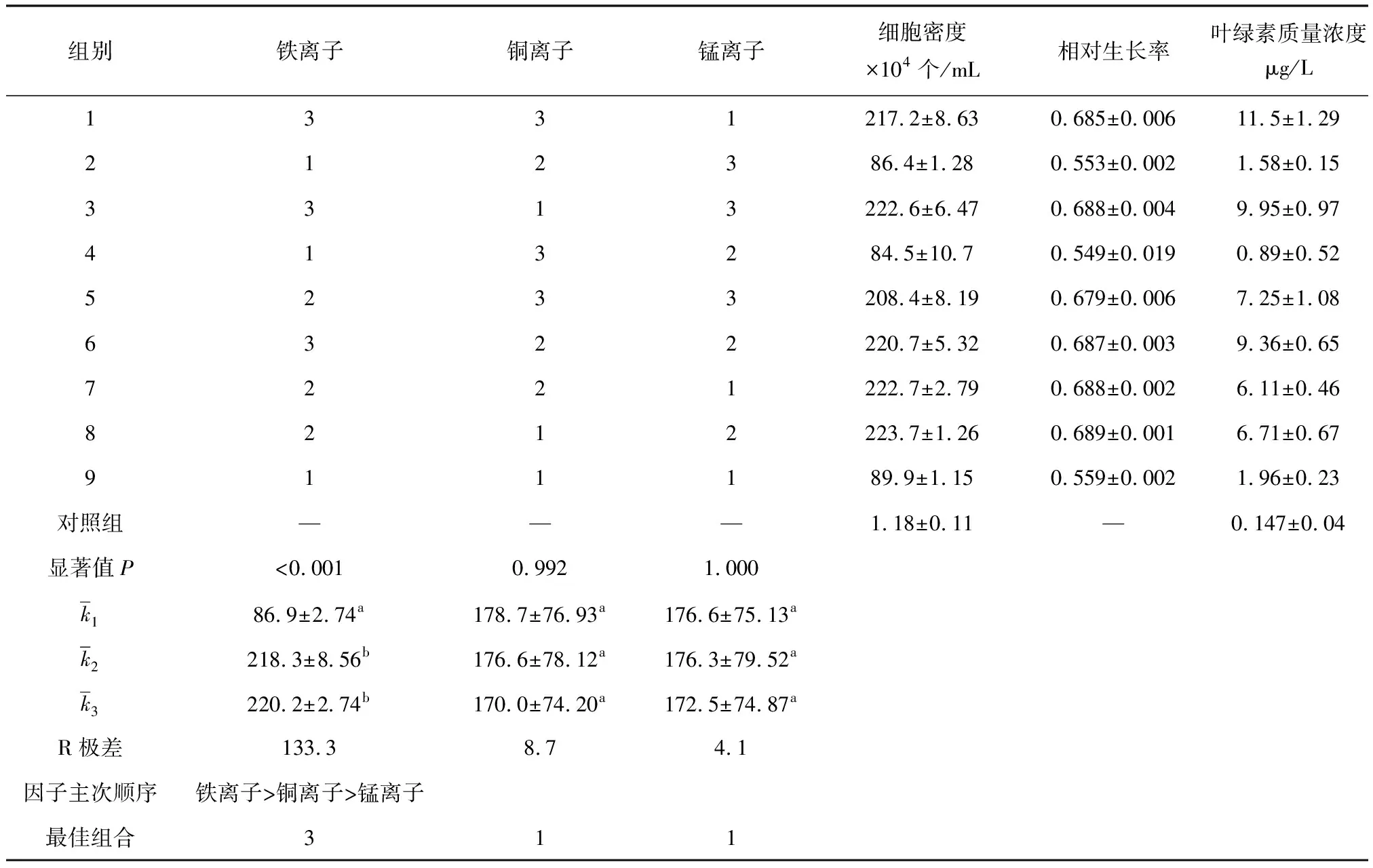
表2 正交试验表及响应值
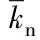
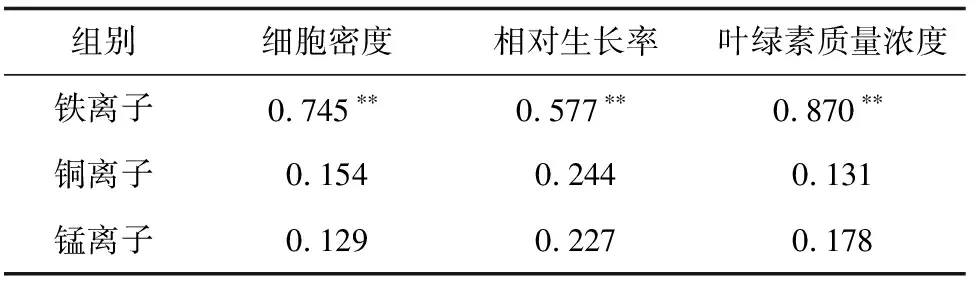
表3 3种重金属离子吸收量与细胞密度、相对生长率和叶绿素含量相关性分析统计表
注:“**”表示相关性极显著(P<0.01),其余为相关性不显著(P>0.05).
3 讨 论
抑食金球藻褐潮形成的元素不仅包含营养盐、温度、盐度和光照等[2,13],重金属对褐潮的形成也有一定的作用。重金属离子是藻类生长发育过程中必不可少的元素,在藻类光合作用、呼吸作用和蛋白质与核酸合成等代谢过程中发挥着重要的作用[14]。低质量浓度的金属离子促进藻细胞生长,而当金属离子超过一定质量浓度时,对藻细胞生长表现为抑制特性。
3.1 锰离子对抑食金球藻生长的影响
锰是藻类叶绿素合成的辅助因子,也是维护叶绿素体膜结构的必需元素,并在光合电子传递过程中参与水的光解作用[15]。同时锰离子也是硝酸还原酶的活化剂,在藻类对硝酸盐的吸收代谢中起着关键的作用[2,6]。缺锰时可能引起藻类叶绿素合成受阻,叶绿素体膜结构被破坏,使藻类光合活性受到抑制[16-17],藻类对硝酸盐的转化过程受到影响,藻细胞生长受到限制。在试验中,锰离子质量浓度为0~25 μg/L时,藻细胞密度随锰离子质量浓度的升高而增加,说明此时锰离子为细胞生长的限制因子。随着锰离子质量浓度的增加,细胞内叶绿素合成逐渐恢复,光合作用和呼吸作用均能正常进行,细胞生长较快。锰离子质量浓度大于25 μg/L时,细胞生长受到抑制。郭金耀等[17]研究了锰离子对盐藻(Dunaliellasalina)的生长、蛋白质和β-胡萝卜素合成的影响,结果表明,当锰离子质量浓度过高或过低时,藻细胞密度较低,同时蛋白质和β-胡萝卜素含量也较低,盐藻生长受阻。曹春晖等[16]研究结果显示,当锰离子浓度为10-12~10-8mol/L时,米氏凯伦藻(Kareniamikimotoi)细胞密度和叶绿素相对含量随锰离子浓度的升高而增加;当锰离子浓度为10-8~10-4mol/L时,细胞密度随锰离子浓度的升高而降低。上述研究结果与本试验结果基本相符。
3.2 铜离子对抑食金球藻生长的影响
铜离子通过与藻细胞中的藻胆体结合,参与光合电子传递过程,促进光合作用的二氧化碳固定, 有利于藻细胞的生长发育。但高质量浓度的铜离子,能使藻细胞积累活性氧,导致脂质和蛋白质的过氧化作用,从而使丙二醛含量升高,超氧化物歧化酶活性降低,阻碍其生长发育[18]。也有学者认为,低质量浓度的铜离子能促进藻细胞生长;但高质量浓度铜离子与细胞表面的官能团结合,使其活性丧失,从而影响细胞的光合作用和呼吸作用,细胞生长受阻[19]。试验中,当铜离子质量浓度为0~5 μg/L时,对细胞生长表现为促进作用;当铜离子质量浓度为25~625 μg/L时,细胞密度随质量浓度的升高而降低。晋利等[20]研究了不同铜离子浓度及不同时间对铜绿微囊藻(Microcystisaeruginosa)生长及叶绿素荧光铜锈的影响,结果表明,96 h内铜离子浓度为1 μmol/L时,其细胞密度、PSⅡ最大光合效率、PSⅡ实际光合效率和光能利用效率与对照组差异不显著(P>0.05),随着铜离子浓度的增加,上述参数均显著降低。张树林等[21]研究了不同质量浓度铜离子(0、0.3、0.6、0.9、1.2、1.5 mg/L)对铜绿微囊藻生长和毒性影响,结果表明,96 h时不同质量浓度铜离子对铜绿微囊藻的生长均产生胁迫作用,细胞密度均比对照组低。其铜离子质量浓度设置较高,最小质量浓度为0.3 mg/L,而本试验中铜离子质量浓度为0~625 μg/L,质量浓度设置较低,当质量浓度大于5 μg/L时,抑食金球藻生长也受到抑制。
3.3 锌离子对抑食金球藻生长的影响
锌是藻类光合作用和生长过程中许多酶的重要组成成分和激活剂,其中碳酸酐酶是藻类吸收利用无机氮的关键酶[22]。高含量锌能导致藻细胞类囊体大多溶解消失,叶绿体遭到破坏,叶绿素a含量下降,光合作用受到影响,并抑制叶绿体总还原型辅酶Ⅱ的形成,从而使细胞生长受阻[23]。高含量锌也能抑制苹果酸脱氢酶的合成,导致细胞糖代谢受到影响。杨洪等[24]试验结果表明,不同浓度(0~100 μmol/L)的锌离子均能对小球藻(Chlorellavulgaris)的生长产生抑制作用。胡晗华等[25]认为,锌含量过高或过低均能抑制藻细胞生长。锌对藻细胞的促进作用与藻种有关,不同的藻类生长所需的锌含量不同。张铁明[22]研究结果表明,当锌离子质量浓度为0.02~1 μg/L时,铜绿微囊藻生长较好,细胞密度显著增加;当锌离子质量浓度大于100 μg/L时,其生长受到影响。锌离子对斜生栅藻(Scenedesmusobiquus)生长的影响也呈现相似的规律,当锌离子质量浓度为0.1~100 μg/L,其细胞密度显著高于对照组;锌离子质量浓度大于1000 μg/L时,藻细胞生长受到影响,细胞密度低于对照组。本试验中,锌离子质量浓度为5~625 μg/L,抑食金球藻生长均受到抑制,这与杨洪等[24]试验结果相符,与张铁明[22]的铜绿微囊藻试验结果的情况也一致,但与斜生栅藻生长所需锌离子质量浓度差异较大。
3.4 铁离子对抑食金球藻生长的影响
铁是藻细胞生长代谢过程中多种酶系的辅助因子,是细胞色素、铁巯基蛋白和铁氧化还原蛋白的重要组成元素[22],能促进藻类的生长,同时也是藻类生长的限制因子之一[22,26]。铁含量较低将导致藻类叶绿素合成受阻[27],光合作用能力下降,藻细胞生长受到影响。在本试验中,当铁离子质量浓度为0~12 500 μg/L时,细胞密度随铁离子质量浓度的升高而增加,叶绿素含量和相对生长率也呈现相同的规律。当铁离子质量浓度较低时,铁离子为藻细胞生长的限制因子,藻细胞随铁离子质量浓度的增加而迅速生长,铁离子表现为促生长作用。当铁离子质量浓度为62 500 μg/L时,铁离子不再是限制因子,此时并未表现出促生长作用。左冬梅等[26]研究表明,在10-9~10-6mol/L时,随着铁浓度的增加,藻类光合放氧及叶绿素合成均受到促进,当铁浓度较低(10-9~10-8mol/L)时,尖刺拟菱形藻(Pseudonitzschiapungens)生长受到抑制。曹春晖等[28]研究结果显示,铁对旋链角毛藻(Chaetoceroscurvisetus)的细胞密度、世代时间和生长率均有显著影响。张铁明[22]试验结果表明,低质量浓度铁离子限制了铜绿微囊藻、斜生栅藻和脆杆藻(Fragilariasp.)的生长,当铁离子质量浓度升高时藻细胞密度和叶绿素含量也升高,表现为促进作用。以上研究结论与本试验结果相符。
3.5 重金属离子对抑食金球藻生长的综合效应
在海域环境中存在有很多藻类生长所需微量元素,这些元素对藻类生长不是单一效应,而是相互间表现为协同或拮抗作用等综合效应[28-29]。曹春晖等[28]研究表明,铁离子对旋链角毛藻生长的影响大于锰离子,其中锰离子对细胞密度没有影响。本试验研究结果显示,铁离子对抑食金球藻生长的影响显著大于铜离子和锰离子。与曹春晖等[28]研究结果相一致。而秦晓明等[29]试验结果显示,影响锥状斯氏藻(Scrippsiellatrochoidea)生长的主次因素为Mn>Fe-EDTA>NO3-N>PO4-P。李灏等[30]通过研究N、P、Fe和Mn对亚历山大藻(Alexandriumsp. LC3)生长的影响得出,4种因素对其影响次序为P>Mn>N>Fe。以上研究得出,铁对藻类生长的影响小于锰,这主要可能由于是藻种的类群不同,不同藻种类群生长发育时对金属离子的代谢途径也不同,本试验研究的抑食金球藻属于棕鞭藻门,而锥状斯氏藻和亚历山大藻均属于甲藻门。金属吸收速率和藻细胞的生长速率有一定相关性[31],本试验通过对3种离子吸收量与抑食金球藻生长的相关性分析得出,铁离子与细胞生长显著相关,而其他2个金属离子与细胞生长相关性均不显著。因此,本试验结果表明,抑食金球藻受到铁离子质量浓度的影响最大。
有研究表明[32-33],近海海域中大量营养盐的输入影响浮游植物对重金属离子的吸收。本研究仅从重金属离子质量浓度方面讨论其对抑食金球藻生长的影响,对于营养盐与重金属联合作用对褐潮形成的影响,重金属离子影响褐潮形成的机制,以及离子质量浓度阈值等方面的认知还需要更进一步的深入研究。
