松江鲈肾组织细胞系生物学特性研究
2019-05-30张晓雨秦艳杰单莉娟周诗嘉王茂林王缙云
张晓雨,李 霞,2,秦艳杰,单莉娟,周诗嘉,王茂林,梁 艳,王缙云
(1.大连海洋大学,辽宁省海洋生物资源恢复与生境修复重点实验室,辽宁 大连116023; 2.大连海洋大学,农业农村部北方海水增养殖重点实验室,辽宁 大连 116023 )

松江鲈(Trachidermusfasciatus)又名四鳃鲈,属鲉形目、杜父鱼科、松江鲈属,为降海洄游性鱼类,是我国渤海、黄海、东海沿岸及相邻淡水水域的习见鱼类。松江鲈肉质鲜美细嫩且体态多变,被誉为“中国四大名鱼”之一[12]。同时,松江鲈由于近年来数量的减少,已被列为国家二级重点保护野生动物。关于松江鲈的生物学研究较少,邵炳绪[13]研究了松江鲈的生长习性;韦正道等[14]报道了影响松江鲈生长的环境因子。近几年关于松江鲈的人工育苗和养殖研究的报道较多[15-18]。在细胞方面,目前已报道的相关研究有松江鲈脾[19]和鳍[20]细胞系的短期体外培养以及重金属对松江鲈肾细胞系的毒性研究[10]等。笔者对可连续传代的松江鲈肾组织细胞系进行生长状况描述和细胞遗传学分析,并测定其对病毒、重金属镉的敏感性,旨在探明松江鲈肾细胞系的特性,为该细胞系在松江鲈病毒性疾病的研究及其作为重金属镉污染的生物学标志物研究提供基础资料。
1 材料与方法
1.1 材料
试验用松江鲈肾组织细胞系由大连海洋大学细胞工程实验室提供。鲤春病毒血症病毒(Springviraemiaofcarpvirus)和鱼类诺达病毒(Piscinenodavirus)由大连海洋大学病害实验室提供。
1.2 方法
1.2.1 细胞培养
细胞培养于25 ℃、5% CO2培养箱,所用的培养基为20% FBS-DMEM/F12,采用文献[18]中胰蛋白消化法传代。
1.2.2 细胞冻存与复苏
参照文献[21]稍作改动,按照1∶1∶3比例,将二甲基亚砜(DMSO)、FBS和DMEM/F12配制成冻存液,并于4 ℃冰箱中保存备用。选取75代生长状态良好,并处于对数生长期的松江鲈肾细胞,用0.25%的胰酶消化后,加入培养基重悬细胞并收集在无菌离心管中。将细胞悬液以转速1500 r/min离心5 min。弃上清液,向离心管中加入1 mL冻存液,重悬细胞,调整细胞密度至1.0×106个/mL后,转移到无菌冻存管中。将装有细胞的冻存管按照顺序进行梯度降温,操作步骤为:4 ℃冰箱中放置30 min,-20 ℃冰箱中4 h,-80 ℃冰箱中24 h,之后转移到液氮(-196 ℃)中,长期保存。
从液氮中取出已冻存30 d的细胞,迅速放在40 ℃的温水中水浴解冻,待冻存液90%融化后,转移至25 ℃的温水中水浴1 min。将细胞悬液转移至无菌离心管中,向离心管中加入1 mL 20% FBS-DMEM/F12的培养基,1500 r/min,离心5 min,弃上清液。在离心管中加入2 mL 20% FBS-DMEM/F12的培养基重悬细胞。充分混匀后,吸取100 μL细胞悬液到青霉素小瓶中,向其中加入100 μL的台盼蓝,在光学显微镜下,用血球计数板进行细胞计数,计算冻存细胞存活率。
细胞存活率/%=(n2/n1)×100%
式中,n1为细胞总数(个),n2为活细胞数(个)。
1.2.3 生长曲线的测定
取生长状态良好的75代松江鲈肾细胞,用0.25%的胰酶消化,加入2 mL 20% FBS-DMEM/F12培养基吹打细胞,制成细胞悬液,调整细胞悬液密度至5×104~8×104个/mL。将细胞悬液接种至24孔板中,每孔接种500 μL,共接种21孔。将24孔板放于25 ℃、5% CO2恒温培养箱中培养,每24 h取3孔细胞进行细胞计数,并计算其密度,连续计数7 d。以培养时间(d)为横坐标,以细胞密度(个/mL)为纵坐标,绘制细胞生长曲线。根据公式计算细胞的群体倍增时间:
T=t×lg2/lg(nt/n0)
式中,T为群体倍增时间(d),nt为时间t(d)后的细胞数(个),n0接种细胞数(个)。
1.2.4 细胞染色体分析
染色体制备方法参照文献[22]并稍做改动。取35代生长状态良好并处于对数生长期的松江鲈肾细胞,向培养基中加入秋水仙素,调整秋水仙素质量浓度为1 μg/mL,放回25 ℃、5% CO2的恒温培养箱中继续培养4 h后,用0.25%的胰酶消化重悬细胞。将细胞悬液转移至无菌离心管中,1000 r/min,离心10 min,弃上清液。向离心管中加入低渗液,在25 ℃下低渗30 min,低渗液为0.075 mol/mL KCl溶液。30 min后1000 r/min离心5 min。用Carnoy固定液固定15 min,连续固定3次,之后放入4 ℃冰箱中过夜。染色体制片方法采用冷滴片法,并在室温下自然干燥。用Giemsa染液染色30 min,用流水轻轻洗去浮色后室温下自然干燥。在显微镜下找到完整的染色体分裂相,统计数目并拍照。
1.2.5 两种病毒对松江鲈肾细胞的感染试验
选取生长状态良好的50代松江鲈肾细胞,吸去旧培养基,用无菌PBS轻轻冲洗细胞2次。将细胞分为3组,分别为2个试验组和1个对照组,2个试验组分别加入0.1 mL鲤春病毒血症病毒液和0.1 mL鱼类诺达病毒液,随后各加入0.1 mL 20% FBS-DMEM/F12培养基,每组设置3个平行。将3组细胞放入25 ℃、5% CO2恒温培养箱中继续培养2 h后,分别吸去病毒液和培养基,加入5 mL 20% FBS-DMEM/F12培养基,再放回25 ℃、5% CO2培养箱中继续培养。每日观察细胞形态并拍照记录细胞病变效应,无论接毒细胞是否出现细胞病变效应,都继续进行盲传接毒,每隔5 d盲传一次。
1.2.6 鲤春病毒血症病毒在松江鲈肾细胞系上的滴度测定
选取生长状态良好的50代松江鲈肾细胞,用0.25%胰酶消化重悬细胞,将细胞悬液密度调整至1.0×105个/mL。将细胞悬液接种至96孔板中,每孔接种200 μL,放回25 ℃、5% CO2恒温培养箱中继续培养48 h后,吸除旧培养基,更换为相同体积的病毒悬液,病毒悬液按照10倍系列稀释,每稀释度8孔。之后放回25 ℃、5% CO2恒温培养箱中继续培养,每日观察细胞形态并拍照记录细胞病变效应出现情况。根据Reed-Muench公式计算病毒滴度值。
1.2.7 松江鲈肾细胞系对镉离子敏感性研究
取对数生长期生长状态良好的60~70代松江鲈肾细胞,用0.25%胰酶消化重悬细胞,调整至细胞密度为1.0×105个/mL,接种在96孔细胞培养板中,每孔接种200 μL悬液。将其放在25 ℃、5% CO2培养箱中培养24 h。弃培养基,加入等体积的含有氯化镉的培养基,镉离子浓度分别为0、20、40、60、80、100、120、140、160、180、200 μmol/L,每个浓度设置3个重复。放回25 ℃、5% CO2培养箱中培养24 h后,向每孔中加入40 μL MTT,放入CO2培养箱中连续培养4 h。弃培养基,用PBS快速冲洗2次。在每个孔内加入150 μL二甲基亚砜(DMSO),室温振荡15 min。用酶标仪测其在570 nm波长下的吸光度值(A)。将氯化镉浓度为0的一组设为阴性对照组,没有细胞只加入相同体积新鲜培养液的一组设为阳性对照组。
细胞存活率/%=(A试验组-A阳性对照组)/(A阴性对照组-A阳性对照组)×100%
2 结 果
2.1 细胞冻存与复苏
松江鲈肾细胞冻存30 d后复苏,细胞贴壁情况良好,36~48 h后,可以铺满瓶底。经计算,存活率为(90.8±1.37)%。
2.2 生长曲线测定
由第75代松江鲈肾细胞生长曲线可见,在25 ℃、5% CO2的恒温培养箱中,于20% FBS-DMEM/F12培养基培养的松江鲈肾组织细胞在0~2 d处于潜伏期,2~4 d进入对数生长期,4~6 d进入稳定期(图1),经计算,松江鲈肾组织细胞的群体倍增时间为55.2 h。
2.3 染色体分析
通过对35代松江鲈肾组织细胞中100个图像清晰、中期分裂相分散较好的松江鲈肾细胞的染色体进行统计计数,发现染色体数目分布为37~42,其中染色体众数为40的细胞占全部计数细胞的70%。通过核型分析可知,核型公式为K(2n)=40=16m+10sm+14t,NF=76,即松江鲈肾细胞有8对中部着丝点染色体(m),5对亚中部着丝点染色体(sm),7对端部着丝点染色体(t)(图2、图3)。
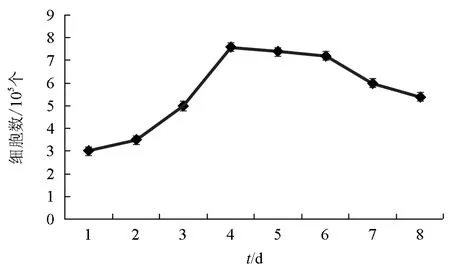
图1 第75代松江鲈肾细胞的生长曲线
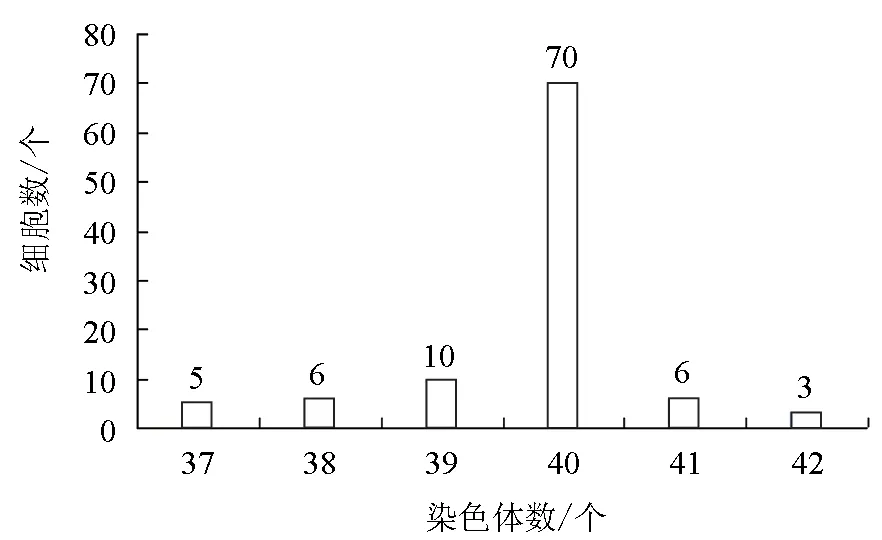
图2 松江鲈肾细胞的染色体数目分布

图3 松江鲈肾细胞的中期分裂相
2.4 两种病毒对松江鲈肾细胞的感染试验
正常松江鲈肾细胞为长纤维型(图4a),被鲤春病毒血症病毒感染3 d后的松江鲈肾细胞收缩变圆、折光度增加,聚集成团或解体死亡。感染4 d后的松江鲈肾细胞碎片增多,细胞脱落,产生典型细胞病变效应(图4b),经测定病毒滴度为106.53/mL。在盲传2代后发现松江鲈肾细胞对鱼类诺达病毒不敏感,细胞未出现病变。
2.5 镉离子对松江鲈肾细胞成活率的影响
用不同浓度的氯化镉处理松江鲈肾细胞后,随着镉离子浓度的增加松江鲈肾细胞存活率逐渐降低,细胞存活率与镉离子浓度有明显的剂量依赖关系(图5)。根据存活率与浓度的自然对数剂量—效应方程y=-0.00448x+1.1025(r2=0.95832),可得半致死浓度为(130±9.1) μmol/L。当镉离子浓度达到200 μmol/L时,细胞难以存活。

图4 正常松江鲈肾细胞(a)和接种病毒后的典型细胞病变效应(b)
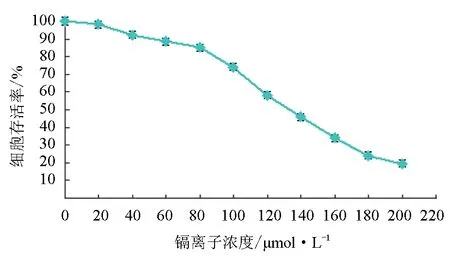
图5 镉离子对松江鲈肾细胞成活率的影响
3 讨 论
3.1 松江鲈肾细胞生长特性分析
曾有研究提出松江鲈肾细胞系的染色体数目与本试验结果相同,均为2n=40,但是其染色体臂数为NF=64和NF=60[26],与本试验中的核型公式K(2n)=40=16m+10sm+14t中NF=76不同。分析可能为松江鲈所处的生活环境和取样时间不同所致。
李树深[27]指出,染色体端部着丝粒较多的种类为原始种类,中部和亚中部着丝粒较多的是特化种类,染色体臂数较少的类群同染色体臂数较多的类群相比,染色体臂数较多的类群特化。绒杜父鱼(Hemitripterusvillosus)同松江鲈同属于杜父鱼科,其核型为2n=46=20m+16sm+10t,NF=82[28]。绒杜父鱼的染色体臂数多于松江鲈,说明绒杜父鱼相比松江鲈,为特化类群,松江鲈端部着丝粒多于绒杜父鱼,中部和亚中部着丝粒少于绒杜父鱼,由此可以说明,松江鲈相较绒杜父鱼,为原始种类。
3.2 松江鲈肾细胞的病毒敏感性
一般情况下,动物病毒对物种及其细胞系的敏感性均有特异性。但是有研究表明,一种鱼类细胞系对几种病毒均具有敏感性,例如斜带石斑鱼(Epinepheluscoioides)脑组织细胞系(GB11)对神经坏死病毒(Neuronecrosisvirus)和石斑鱼虹彩病毒(Grouperiridovirus)敏感[29]。同时也有研究发现,一种病毒能感染几种细胞系,例如中华鳖病毒(Trionyxsinesisvirus)能使鲤鱼上皮乳状瘤细胞、鳗鲡性腺细胞和草鱼卵巢细胞产生病变[30];神经坏死病毒感染斜带石斑鱼脑细胞[29]和花鲈(Lateolabraxmaculatus)脑细胞[31]后,均能使细胞产生病变。
鲤春病毒血症病毒,又名鲤弹状病毒,属弹状病毒科,水泡病毒属(Vesiculorirus)。鲤春病毒血症是由鲤春病毒血症病毒引起鲤鱼科的一种急性、出血性、传染性疾病,死亡率极高,多发于鲤科鱼类,是全球性鱼类疾病[32]。研究表明,本试验建立的松江鲈肾组织细胞系对鲤春病毒血症病毒敏感,可用来分离和扩增该病毒。目前在松江鲈的人工养殖中尚未发现大规模病毒病的暴发,但肾组织细胞系的建立可为将来病毒病的研究打下基础。
3.3 镉离子对松江鲈肾细胞成活率的影响
镉是我国水生生态系统中常见的重金属污染物,具有高致毒性,较低剂量的镉就能使水生动物中毒,甚至死亡[33]。本试验用不同浓度的镉离子处理松江鲈肾细胞24 h后,结果显示,随着镉离子浓度的升高细胞存活率降低,半致死浓度为(130±9.1) μmol/L。近年来,关于镉离子对鱼类的毒性研究较多,镉离子对大弹涂鱼(Boleophthalmuspectinirostris)的半致死浓度为433.2 μmol/L[34],对鲫鱼和泥鳅(Misgurnusanguillicaudatus)的半致死浓度分别为344 μmol/L和351 μmol/L[35],说明相对于成体鱼,细胞系对镉离子更敏感,这可能与鱼体本身具有自我调节能力有关,但也说明细胞系更适宜用来作为水质监测的指标和机理研究。
