一种新型贝壳鱼礁的大型底栖藻类附着效果初探
2019-05-30姜大为郝振林王路平李多慧张瑜洋
赵 静,姜大为,郝振林,郭 栋,王路平,李多慧,张瑜洋
(1.大连市水产研究所,辽宁 大连 116087; 2.大连海洋大学,农业部海洋水产增养殖学重点开放实验室,辽宁 大连 116023; 3.辽宁省海洋水产科学研究院,辽宁省海洋生物资源与生态重点实验室,辽宁 大连 116023 )
近年来,由于近海渔业生产捕捞强度过大和沿海经济发展带来的环境污染,导致近岸环境恶化,海洋生物的产卵场和索饵场消失,渔业资源严重衰退。为了有效改善近岸海洋生态环境,遏制渔业资源衰退趋势,我国自20世纪70年代末开始人工鱼礁建设,经历了近半个世纪的探索和实践,已取得显著成效[1]。人工鱼礁是人为设置在海里的构造物,用于改善海域生态环境,为海洋生物提供索饵、繁殖、生长、发育等场所,从而达到保护和增殖海洋生物资源的目的[2-3]。随着科学技术的发展,人工鱼礁的材质也日益向着生态化、节能化、绿色化方向发展,主要包括天然材料、废弃材料、建筑材料、综合材料等。资料显示,目前用于构建人工鱼礁的材料已超过249种[4]。研究表明,藻类附着效果与鱼礁的材料和形状密切相关,为利于海藻孢子的附着和生长,藻礁的设计多为表面凹凸,表面和内部多孔结构,礁体内部添加肥料和藻类生长所需的营养物质等[5-7]。
我国人工鱼礁建设处于探索阶段,用于鱼礁建设的材料多为混凝土构件和石块,对于其他材料的研究缺乏相关数据积累。本研究选择一种新型贝壳鱼礁(简称贝壳礁)投放在自然海域,在相同海域设置混凝土构件礁区和空白对照区,于投礁后1个月、半年和1年开展主要环境因子监测和大型底栖藻类的调查。根据调查数据,分析3个区域藻类群落演替规律及其与环境因子的关系,评价贝壳礁上藻类的附着效果,旨在为人工鱼礁选材及相关海域海洋牧场建设提供科学依据。
1 材料与方法
1.1 礁体材料
新型贝壳鱼礁是由日本Doken株式会社研发,在日本经过15年的使用和推广已取得突出效果,对海参、海胆等海珍品有较好的诱集和增殖效果,是目前处于世界领先水平的专利产品。2012年由大连源正海物实业有限公司引进,在大连市长海县大长山岛生产。主要成分由虾夷扇贝(Patinopectenyessoensis)壳及生物固化剂组成,具有很高的强度和很好的透水性。单体质量约为2.5 t,渔礁的外径长2.4 m,高0.75 m。鱼礁外轮为十二边形,内轮为正六边形,外轮和内轮每隔一个侧面留一个用于海水流动的开口。同时内轮比外轮高出15 cm,鱼礁中心及内外轮交界处留有泥沙沉淀口,可以及时将泥沙排除(图1)。
混凝土构件礁采用凹槽形构件礁,凹槽形构件礁参数为1.2 m×1.2 m×1.0 m,中心圆内径长0.35 m,礁体表面黏附小砂砾和石块,以增加表面粗糙度(图2)。
1.2 鱼礁建设海区
人工鱼礁区项目建设位于大连市长海县大长山岛近海,N 39°16′20″~39°19′12″,E 122°40′48″~122°45′00″,水深5~20 m,该海域受北黄海环流影响,海水平均流速约0.1 m/s,海域内无大的入海河流和排污口,海底多为砾石和砾砂带[8]。试验在目标海域选取A、B、C 3个区域,本底调查结果显示,3个区域底质和水质理化因子情况基本相同。2013年10—11月向A区投放混凝土构件礁3000件,礁体间距为礁体尺寸的1.0~1.5倍,礁区面积达10 000 m3·空;同一时间向C区投放贝壳礁1700件,礁体间距为礁体尺寸的1.0~1.5倍,礁区面积达10 000 m3·空;B区,不投放人工鱼礁,以自然海底礁石作为对照。3个区域均设6个调查站位,共计18个(图3),分别于投礁1个月(2013年12月)、半年(2014年6月)和1年(2014年12月)后,对3个区域开展海洋环境和大型底栖藻类资源状况调查。
1.3 样品采集和处理
利用多参数水质仪(YSI556MPS)测定礁区海域溶解氧、pH、盐度和水温。依据《海洋监测规范》(GB 17378—2007)[9]和《海洋调查规范》(GB/T 12763—2007)[10],在预设的站位用采水器取样,采集样品装瓶标记,带回实验室分析。测定的水质指标包括:氨氮、硝酸盐、亚硝酸盐、磷酸盐和叶绿素a。
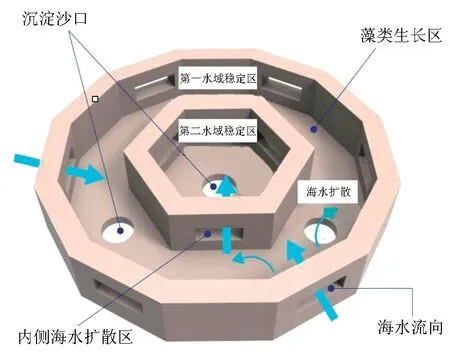
图1 新型贝壳人工鱼礁结构

图3 调查站位
大型底栖藻类调查采用规格为50 cm×60 cm的PVC管制样方框,由潜水员水下作业完成采样,分别于投礁后1个月、半年和1年,对3个区域取样,每个区域平行取3个样方,以确保样品的广泛性和试验的准确性。样品采集后转入自封袋内密封,内置标签,当日带回实验室,用海水冲掉沙砾,挑去杂质,对海藻进行分类,记录海藻的数量。用吸水纸吸去藻体表面海水,称量湿质量,并计算出生物量(g/m2)。
用PRIMER 5.0软件包中的Margelf丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数,进行不同礁区大型底栖藻类生物多样性研究[11-12]。利用Canoco 4.5软件对藻类群落和环境因子进行去趋势对应分析、主成分分析和冗余分析,以研究藻类群落结构与环境因子的相关关系[13]。
2 结果与分析
2.1 3个区域附着藻类的种类组成、生物量及季节变化
经过3次潜水采样调查,共发现3门12种大型底栖海藻。其中绿藻门3种,分别为孔石莼(Ulvapertusapertusa)、羽藻(Bryopsisplumosa)、刺松藻(Codiumfragile);红藻门5种,分别为石花菜(Gelidiumamansii)、鸡毛菜(Pterocladiacapillacea)、中间石枝藻(Lithothamnionintermedium)、蜈蚣藻(Grateloupiaasiatuca)、角叉菜(Chondrusocellatus);褐藻门4种,分别为海黍子(Sargassummuticum)、鼠尾藻(Sargassumthunbergii)、海带(Saccharinajaponica)、裙带菜(Undariapinnatifida)(表1)。褐藻门藻类生物量最大,全年平均值为199.78 g/m2。历次取样中,2013年12月空白对照区藻类生物量最小,为33.83 g/m2、藻类种类数为7种;而2014年6月贝壳礁区藻类生物量最大,为276.83 g/m2,藻类种类为10种。藻类总生物量投礁前后依次为2013年12月(122.83 g/m2)<2014年12月(333.17 g/m2)<2014年6月(481.40 g/m2)。其中,红藻门藻类和褐藻门藻类与藻类群落总生物量的变化趋势一致;而绿藻门藻类生物量投礁前后变化趋势略有不同,依次为2013年12月(22.59 g/m2)<2014年6月(50.50 g/m2)<2014年12月(98.86 g/m2)(图4)。3个区域附着藻类生物量的变化趋势大致相同,夏季(6月)高于冬季(12月),不同区域间藻类年平均生物量依次为贝壳礁区(152.19 g/m2)>混凝土构件礁区(90.83 g/m2)>空白对照区(46.79 g/m2)(图5)。
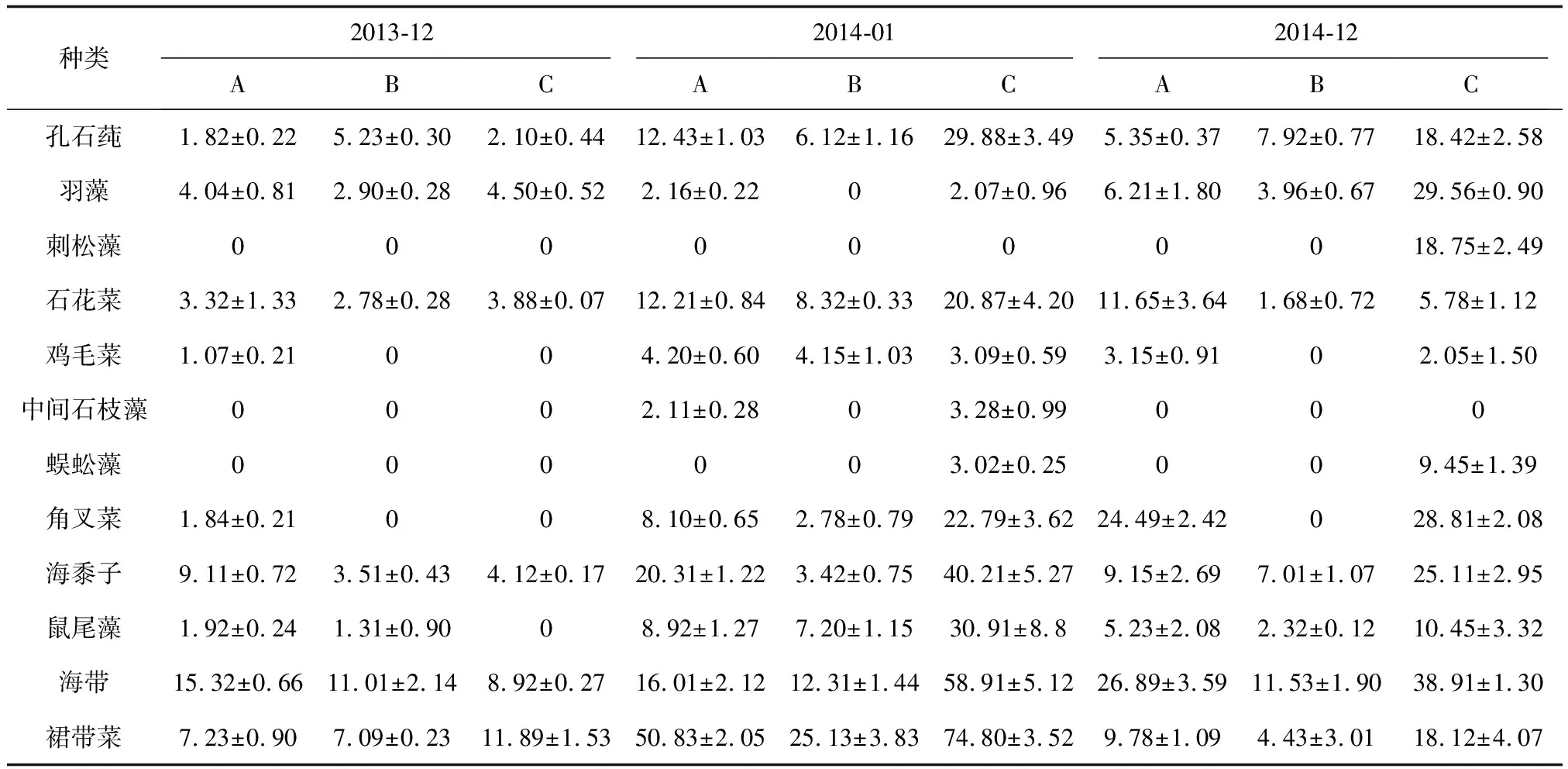
表1 3个区域投礁前后附着藻类的生物量分布 g/m2
注:A.混凝土构件礁区; B.空白对照区; C.贝壳礁区.下同.
2.2 3个区域附着藻类的群落结构特征
根据藻类群落参数(表2)可知,投礁初期,贝壳礁区藻类群落的物种丰富度指数和多样性指数低于另外2个区域;随着投礁时间延长,贝壳礁上附着藻类群落的物种丰富度指数和多样性指数均有所增长,投礁半年后,贝壳礁区的物种丰富度高于空白对照区,但低于混凝土构件礁区,多样性指数均高于另外2个区;投礁1年后,贝壳礁区附着藻类群落的物种丰富度指数和多样性指数均高于另外2个区。冬季各区域的均匀度高于夏季,相同季节投礁后各区域的均匀度高于投礁前,但差异不明显,贝壳礁区夏季均匀度最低。
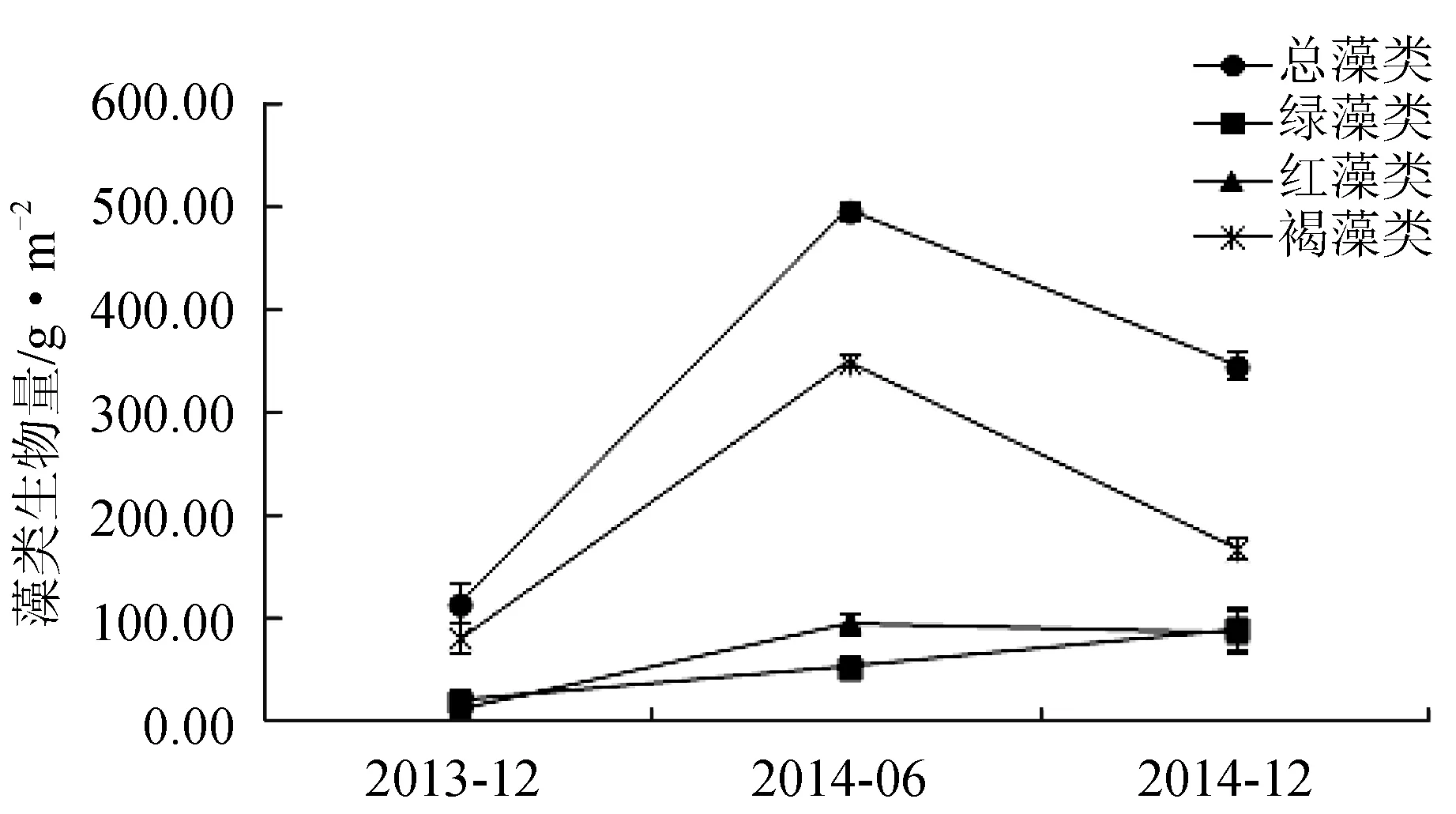
图4 不同采样时间不同附着藻类的生物量变化
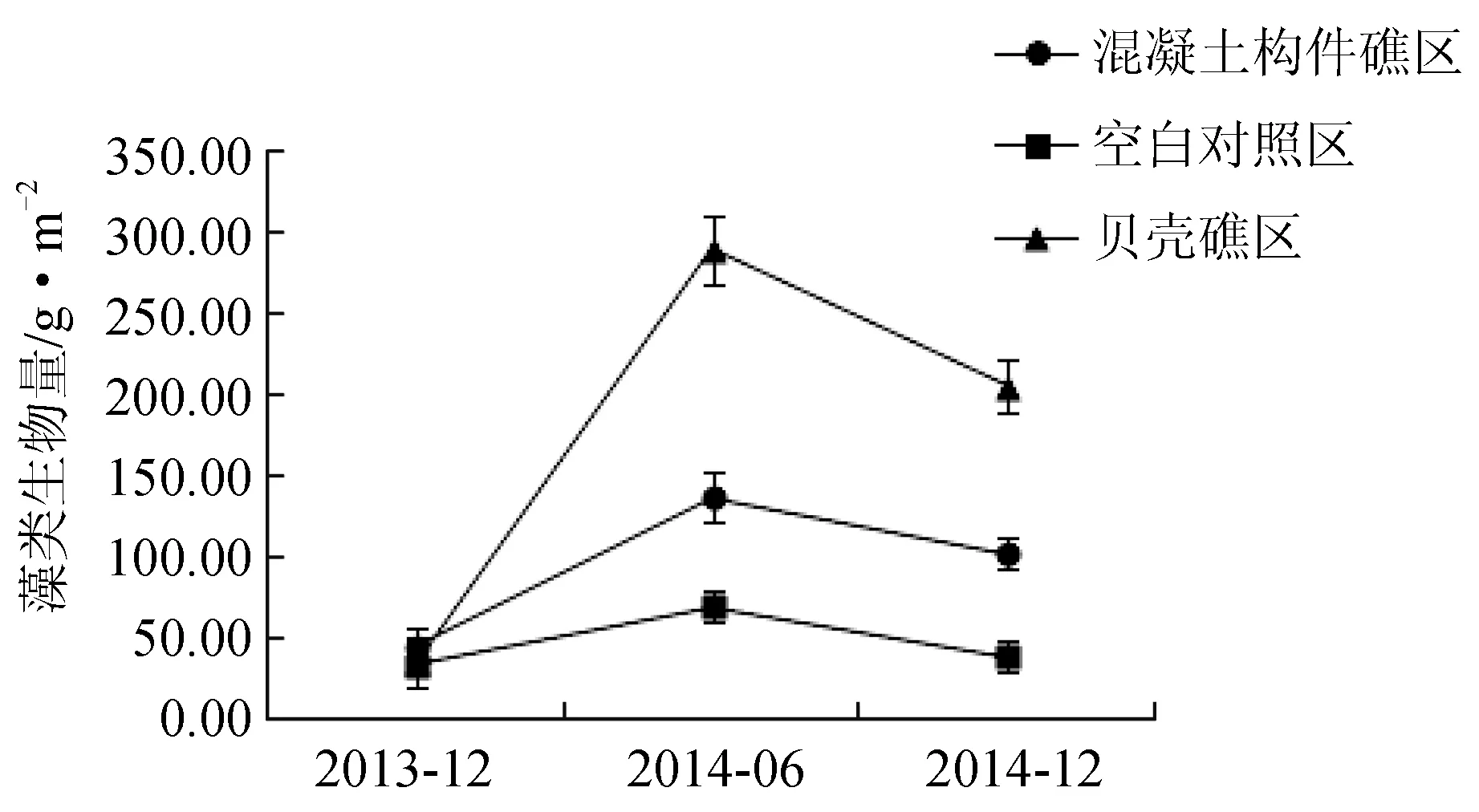
图5 不同采样时间3个调查区域附着藻类的生物量变化

采样时间调查区域样方编号种类数生物量/g·m-2种类丰富度多样性指数均匀度2013-12A1945.672.091.860.852013-12B2733.831.701.760.902013-12C3635.411.401.640.922014-06A410137.281.831.910.832014-06B5869.451.651.820.882014-06C611289.831.761.990.832014-12A79101.901.731.970.902014-12B8738.851.641.800.912014-12C911205.411.882.220.93
2.3 3个区域附着藻类的群落动态与环境因子的关系
采用去趋势对应分析对3个区域3次采集的9个样方进行分析,结果表明,4个排序轴的特征值分别为0.1933、0.0246、0.0015、0.0008,第1轴特征值最大,体现了最多的生态信息,第2轴次之。根据样方在前2轴的得分做出二维排序图(图6),物种的分布表现为某些物种之间关系非常密切(图中两物种之间的距离表示两个物种间的亲疏关系,距离越近越亲密,反正差异越大[14])。
去趋势对应分析排序的结果显示,4个轴的长度均小于3,数据类型符合线性模型。综合9个环境因子组成9×9矩阵以及藻类群落生物量矩阵进行主成分分析。9个环境因子对藻类群落生物量的总解释量为96.6%。环境因子前2个轴对环境的解释量分别为65.8%和21.7%,累积解释量为87.6%,说明前2个轴很大程度可以反映环境数据的变化,其中与第1轴相关性较高的环境因子为磷酸盐、叶绿素a和氨氮;与第2轴相关性高的为氨氮、温度、亚硝酸盐和硝酸盐等。褐藻门的鼠尾藻、海带、海黍子和红藻门的角叉菜受第1轴影响较大,即与磷酸盐、叶绿素a和氨氮正向相关。绿藻门的羽藻、刺松藻和红藻门的蜈蚣藻受第2轴影响较大,即与氨氮、温度、亚硝酸盐和硝酸盐负相关(图7)。

图6 基于去趋势对应分析的藻类群落样方及物种排序Ulv,孔石莼;Bry,羽藻;Cod,刺松藻;Gel,石花菜;Pte,鸡毛菜;Lit,中间石枝藻;Gra,蜈蚣藻;Cho,角叉菜;Sar,海黍子;Thu,鼠尾藻;Sac,海带;Und,裙带菜.下同.
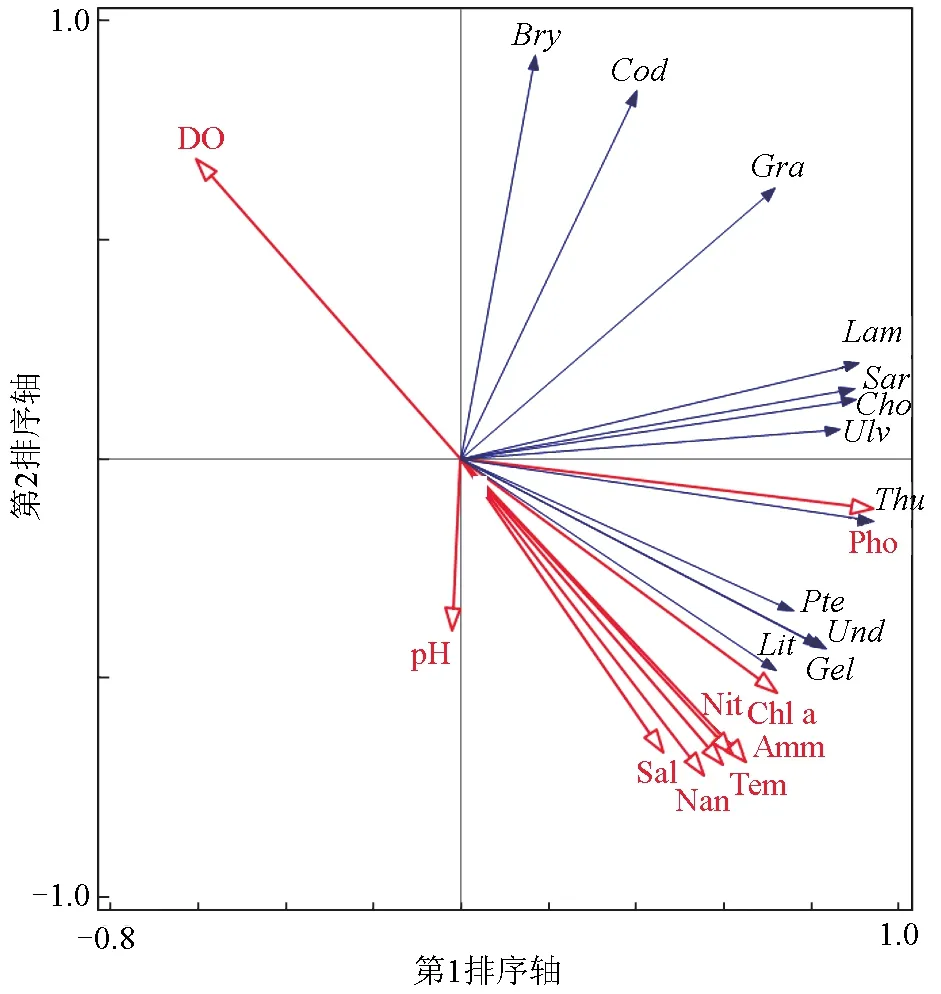
图7 基于主成分分析的藻类生物量—环境排序Tem,温度;Sal,盐度;DO,溶解氧;Amm,氨氮;Nit,亚硝酸盐;Nan,硝酸盐;Pho,磷酸盐;Chl a,叶绿素a.
分别对3个区域的藻类群落和主要环境因子指标进行冗余分析。结果显示,影响3个区域藻类群落变化的第1环境因子均为水温,影响混凝土构件礁上附着藻类群落变化的第2、3影响因子为氨氮和亚硝酸盐,影响自然礁石(空白对照区)附着藻类群落变化的第2、3影响因子为磷酸盐和氨氮,影响贝壳礁上附着藻类群落变化的第2、3环境因子为溶解氧和叶绿素a(表3)。
3 讨 论
大型底栖海藻主要由生活在岩礁或砾石上的红藻、褐藻、绿藻和少量蓝藻等组成,是海洋生态系统的重要组成部分,在海洋能量流动、物质循环和信息传递中发挥着极其重要的作用[15-16]。根据《中国黄渤海海藻》[17]记载,辽宁海区有大型海藻228种。本次在3个试验海区共采集到12种大型底栖海藻,其中红藻门种类最多,褐藻门生物量最高。从藻类生物量的增长趋势来看,红藻门、褐藻门和总藻类的增长趋势相同,呈现夏季高于秋季、投礁后高于投礁前的趋势;绿藻门略有不同,呈现投礁后持续增长的趋势。红藻门的石花菜和角叉菜,褐藻门的鼠尾藻、海带和裙带菜生长盛期均在春、夏季[18],多为大型藻类,在生物量上占优势,对藻类总生物量影响较大。绿藻门的羽藻和刺松藻生长盛期在秋季[18],在投礁当年的秋季才显示出优势,但因其体型较小,对藻类总生物量的贡献小。
本试验设置的3个调查区均位于同一海区,投礁前本底调查结果显示,3个区域底质和水质理化因子情况基本相同,投礁后1个月调查发现,3个区域藻类种类和数量差异不大,随着投礁时间延长,2个投礁区(混凝土构件礁和贝壳礁)大型底栖藻类的生物量和种类均大于空白对照区,其中投放贝壳礁的区域藻类生物量和种类增加得更为显著。表面凹凸和内部多孔结构的鱼礁更有利于海藻孢子的附着和生长[19-20],分析认为贝壳礁结构上增加了鱼礁的表面积,材料为粉碎贝壳和固化剂,表面凹凸不平且透水性好,这些设计为藻类生长提供了良好的条件。从这种鱼礁在日本多年的使用效果来看,其恢复渔业资源和水域生态环境的效果良好,更有利于藻类的附着和生长。本试验结果也表明,投放贝壳礁的区域藻类数量和生物量增加都更为显著。
生物多样性和群落结构的变化反映生态系统的波动。生物多样性在生态系统变化过程中的作用已引起全球关注[21]。分析3个调查海区藻类群落结构和生物多样性,横向比较来看,投礁前贝壳礁区的藻类群落的物种丰富度指数和多样性均低于其他2个海区,投礁后贝壳礁区藻类群落的物种丰富度指数和多样性逐渐超过空白对照区和混凝土构件礁区。纵向比较来看,贝壳礁区藻类群落的物种丰富度指数和多样性指都高于投礁前,且呈现持续增长的趋势,均匀度投礁后夏季最低,可能是由于夏季海黍子、海带和裙带菜等生物量占绝对优势,致使群落的均匀度较低。由此可知,贝壳礁区藻类群落结构有更早变稳定的趋势。

表3 影响3个区域藻类群落变化的重要环境排序
基于去趋势对应分析的样方排序图可以反映样方的相似关系,桂东伟等[22]在研究昆仑山北坡策勒河流域生态因素对植物群落的影响时认为,去趋势对应分析可以有效反映植物样方的相似关系。去趋势对应分析结果表明,褐藻门的鼠尾藻、海带和海黍子关系较近,绿藻门刺松藻和红藻门的蜈蚣藻关系较近。生物群落变化与环境因子的梯度响应模型主要有线性模型和单峰模型两种,由于去趋势对应分析4个排序轴长度均小于3,因此适用于线性模型。在线性模型中,主成分分析旨在利用降维的思想,把多指标转化为少数几个综合指标(即主成分),其中每个主成分均能反映原始变量的大部分信息,且所含信息互不重复。9个环境因子对藻类群落生物量的总解释量为96.6%,说明选取的环境因子合理。结合主成分分析结果得出,藻类样方点在排序轴上的分布沿第1、2轴均有明显的梯度变化。生长盛期在夏季的鼠尾藻和海带与磷酸盐、叶绿素a和氨氮正相关,生长盛期在秋季的松藻和绿藻与氨氮、温度、亚硝酸盐和硝酸盐负相关。分别对3个区域的藻类群落和主要环境因子指标进行冗余分析,结果显示,影响3种礁体上藻类群落变化的第1环境因子均为水温。温度是植物生长最重要的影响因素之一,任何植物的生长、发育都需要一定的热量和温度范围,超过或者低于适宜温度时,植物生长受抑制。除了水温外,氮、磷营养盐仍然是制约自然礁石区和混凝土构件礁区上藻类群落的第2、3环境因子;而在贝壳礁区,氮、磷营养盐的影响不再显著,叶绿素a和溶解氧成为贝壳礁区限制藻类群落的第2、3环境因子。分析认为,由贝壳礁特殊的材料和结构而营造的附近水域微环境中营养盐丰富,使得氮、磷营养盐不再成为限制藻类群落的主要因子。一方面贝壳礁能有效增加礁体的生物附着量,改善海区生态系统,增加水域生产力(叶绿素a)[23-24];另一方面随着礁区生态系统日益复杂,溶解氧含量变化较大,对藻类群落结构会产生较大影响。此外,3个区域环境因子除了与季节、礁体形状和材料相关外,还可能受到周边海区水产养殖和陆源输入等其他因素的影响,笔者仅收集了9个环境因子,可能存在一些不足,贝壳礁与大型底栖藻类的关系有待进一步的研究。
本研究表明,投放贝壳礁的海区大型藻类数量和种类增长的幅度更为明显,且藻类群落结构更趋于稳定,贝壳礁的藻类附着效果较好;贝壳礁特殊的材料和结构更能有效改良海区生态环境,使氮、磷营养盐不再成为限制藻类群落的主要因子。本试验采用的贝壳礁已在日本海域使用15年以上,其在形状设计、材料组成和生态修复效果上具有以下优点:(1)表面粗糙,能为附着生物提高良好附着基[25];(2)来源广泛,材料绿色,少污染;(3)相比混凝土礁具有良好的生物亲和性,不会释放碱性物质[26];(4)贝壳礁区生态更快达到稳定阶段,有利于发挥人工鱼礁生态效益,修复海洋环境。因此,利用废弃贝壳建设人工鱼礁,是使用生物材料进行海洋生态修复的有效方法,值得大力推广和发展。同时,关于贝壳礁的设置布局和相关海洋环境调控功能还有待进一步研究。
