高山草原鼠兔(Ochotona curzoniae)洞口密度与植被物种和土壤性质的关系:以申扎县为例
2019-05-28詹天宇
侯 阁,詹天宇,刘 苗
(1.中国科学院地理科学与资源研究所 生态系统研究网络综合研究中心,北京 100101;2.成都理工大学旅游与城乡规划学院,四川 成都 610059)
高原鼠兔(Ochotona curzoniae)是青藏高原一种特有的小型穴居类啮齿动物,其栖息地主要分布在植被低小、视野开阔的草地生境,主要位于我国的青海、甘肃南部、四川西北部和西藏等青藏高原区域的高山草甸和高山草原[1-3]。高原鼠兔喜食双子叶植物的叶片,其挖掘土壤以及产生排泄物等活动对高原草地产生一定的影响,是高原草地生态系统的关键种[2-5]。
关于高原鼠兔的研究主要集中在高原鼠兔栖息地的生境状况以及对草地生态系统的影响作用。栖息地的生境状况对高原鼠兔分布数量具有重要的影响[3, 6]。首先,高寒草原植物的高度和盖度是高原鼠兔数量的主要制约因素,同时植物的种类以及群落组成也很大程度上影响着高原鼠兔的分布,比如高寒草甸禾本科和棘豆科植物对鼠兔的分布数量具有一定的促进作用[6-8];其次,栖息地土壤物理环境也影响着高原鼠兔分布数量,研究发现,高寒草甸土壤含水量极高或极低的草地都不利于鼠兔的生存[9-10]。此外有研究表明,高寒草甸土壤紧实度越高,鼠兔数量越多;高原鼠兔栖息地的选择过程中,不同草地生境特征对高原鼠兔的分布数量具有一定的制约作用[11]。与此同时,不同数量鼠兔分布活动对草地生境同样也产生不同程度的反馈作用。
目前,高原鼠兔对高寒草原生态系统的影响作用还存在着很大的争议[8, 12]。有研究表明,高原鼠兔在高寒草原生态系统中发挥消极作用[13],主要有以下几方面原因:首先,鼠兔在土壤挖掘活动中,将下层土壤暴露于地面,致使土壤营养和水分流失,从而导致土壤肥力下降[14-15];其次,鼠兔对植物的种子、茎叶以及根茎的啃食,使植物遭到破坏甚至死亡,从而导致植物生产力以及群落多样性降低[7-8];最后,鼠兔洞穴的分布,使草地大面积裸露,有助于杂草生长,导致草地退化甚至沙化[13, 16]。持相反观点的学者则认为鼠兔活动可以改变高寒草甸土壤结构,增加土壤有机质含量、水分含量以及营养元素含量,从而促进植物的生长[2, 10]。高原鼠兔作为高寒草原生态系统的关键种,鼠兔的摄食活动可促进植物种子的传播,从而提高生态系统动植物的多样性,提高生态系统的稳定性[17-19]。刘菊梅和司万童[20]研究表明,鼠兔不会直接导致草地沙化,且适当的鼠兔活动对植物种群结构具有一定的促进作用。
但是,现有关于高原鼠兔的研究大多集中在青藏高原高寒草甸,而关于青藏高原高山草原鼠兔的研究却甚少。申扎县在青藏高原中部地区,属于高山草原类型,也是高原鼠兔的主要分布区域[21]。高山草原鼠兔栖息地生境如何影响鼠兔分布数量,高山草原生态系统对不同数量高原鼠兔的干扰具有怎样的响应,成为了研究亟待解决的科学问题。因此,本研究通过对申扎地区不同洞口密度高原鼠兔分布栖息地,土壤理化性质,植被高度、盖度以及群落组成进行调查分析,探究高山草原草地生境、高原鼠兔洞口密度以及草地生态系统之间的关系,以期对高山草原生态环境保护提供一定的理论依据。
1 研究区概况与方法
1.1 研究区概况
申 扎 县 (87°40'-89°48' E, 30°10'-32°10' N)位于青藏高原西南区域,平均海拔在4 750 m左右(图1);属于高原亚寒带季风气候,气温偏低,年均气温为0.4 ℃左右,气候干燥,年均降水量仅为 298.6 mm,且年均蒸发量高达 2 181.1 mm,年均日照时数长达2 919.5 h[22-24]。该地区可利用草地面积160万hm2,约占申扎总面积的40%,其主要发育高山草原植被,以紫花针茅(Stipa purpurea)为主,伴生种有羊茅(Festuca ovina)、毛瓣棘豆(Oxytropis sericopetala)、苔草 (Carexspp.)、火绒草 (Leontopodium nanum)等[22-23];土壤类型主要以高山草原土、高寒草甸土、高山荒漠土为主[24]。
图1 研究区区位图Figure 1 Location of the study area
1.2 试验设计与野外调查
2017年8月在青藏高原那曲地区申扎县草原监测站附近,选取了 5 个 25 m × 25 m(625 m2)的样地,通过观察鼠洞洞口土壤的新旧程度和洞口状况,排除废弃洞口,计算统计有效鼠洞数量,将5 个样地划分为 T1(176 个·hm-2)、T2(496 个·hm-2)、T3(752 个·hm-2)、T4(992 个·hm-2)和 T5(1 296 个·hm-2)5个鼠洞密度梯度。每个梯度样地内,随机选取3个0.5 m × 0.5 m 的样方,样方间距离不小于 5 m,进行样方调查统计。样方调查统计内容包括植物个体的名称、株数、高度、盖度以及土壤温度、紧实度等。植物盖度的测定,利用目测法对样方内每种植物垂直投影下的面积进行统计,即为该物种的盖度(%),所有植物盖度之和,则为总盖度(%)。土壤温湿度的测定,主要利用土壤温湿度计,对0-10、10-20以及20-30 cm深度土壤温度进行3次重复测定,最终求得3层土壤的温度和湿度的均值;土壤容重的测定,利用环刀对0-10、10-20和20-30 cm深度土壤3次重复取样,在105 ℃的温度下烘干至恒重称重,最终求得3层土壤的容重均值。调查完植被特征和土壤性质后采集土壤样品,按照0-10、10-20以及20-30 cm分层收集土样,装入密封袋带回实验室分析。利用收割法对地上生物量进行测定,将样方内植物地上部分刈割,分植物物种装入纸袋,通过 105 ℃ 杀青 2 h ,80 ℃ 恒温下烘干至恒重,称其质量为植物群落地上生物量。
1.3 土壤数据测定
土壤全碳 (soil total carbon,STC)、全氮 (soil total nitrogen,STN)采用燃烧法测定,土壤全磷(soil total phosphorus,STP)采用钼锑抗比色法测定,土壤有机碳 (soil organic carbon,SOC)采用重铬酸钾外加热法测定,土壤速效氮 (soil available nitrogen,SAN)采用碱解扩散法测定,土壤速效磷(soil available phosphorus,SAP)采用NaHCO3浸提钼锑抗比色法测定,测定方法均参考《土壤农化分析》[25]。
1.4 数据处理与分析
采用Margalef指数和Simpson指数分别评测样地植物物种丰富度指数和多样性指数[26],具体计算公式如下:

式中:S为物种数,i为第i种,Pi为第i种的多度比例;Pi=Ni/N,Ni为第i种的个体数,N为所有种类总个体数。
本研究利用Excel软件对数据进行基础的处理,利用 SPSS 16.0 (SPSS,Chicago, IL,USA)对数据进行方差差异性分析,利用 Sigmaplot 10.0 (Systat Software,Inc.,Chicago,IL,USA)软件采用相关性分析方法,分析不同高原鼠兔鼠洞密度下植物及土壤性质的变化规律;采用主成分分析法(Principal Component Analysis,PCA)以及热度图 (Heat map)方法对高原鼠兔鼠洞密度、植物以及土壤特征数据进行分析并利用 R (R Foundation for Statistical Computing,Vienna,Austria)软件作图[27-29]。此外,利用 ArcGIS 10.2(ESRI,Inc.,Redlands,CA,USA)描述研究区基本概况并出图 (图 1)。
2 结果与分析
2.1 不同鼠洞密度土壤环境的差异性
不同高原鼠兔鼠洞密度下,草地生境土壤物理性质存在差异(图2)。随着鼠兔密度的增加,土壤温度、土壤含水量以及土壤容重总体上都呈下降趋势,但是土壤含水量的变化趋势未通过显著性水平检验 (P> 0.05)(图 2B);此外,T1梯度的土壤温度和土壤容重与 T4、T5梯度差异显著 (P< 0.05);T1梯度土壤含水量与其他梯度差异显著(P< 0.05)。
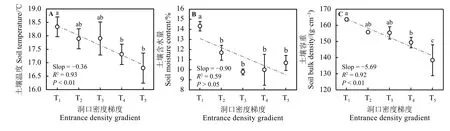
图2 不同高原鼠兔密度梯度下土壤环境的差异性Figure 2 Differences of soil environment under different Ochotona curzoniae density gradients
2.2 不同鼠洞密度植被群落组成的差异性
随着高原鼠兔鼠洞密度的增大,植物群落物种种类减少,植物总数量增加。主要表现为高山豆(Tibetia himalaica)数量明显增多,冰川棘豆(Oxytropis glacialis)数量相对稳定,其他植物数量减少,在T5梯度仅存高山豆和冰川棘豆两种物种(表1)。
2.3 不同鼠洞密度下土壤营养元素和植物的变化特征
在不同数量鼠兔的干扰下,高寒草原土壤全碳(R2= 0.60)、土壤全氮 (R2= 0.56)、土壤全磷 (R2= 0.73)、土壤有机碳 (R2= 0.63)、土壤速效氮 (R2= 0.66)以及土壤速效磷 (R2= 0.36)均呈现出增加的趋势 (slope >0),但均未达到显著性水平 (P> 0.05)(图 3)。此外,方差分析(ANOVA)结果显示,土壤速效性养分在不同数量鼠兔梯度下差异显著(P< 0.05)(图3D、E、F),且速效氮在T4和T5梯度离散程度大(图3E);T3、T4和T5梯度下速效磷同样表现出较大的离散性 (图 3F)。
随着鼠兔数量的增加,草地物种多样性降低,地上生物量增多;植物物种多样性Simpson指数与Margalef指数均呈现下降的趋势,即高寒草原鼠兔数量过多导致物种多样性和丰富度降低;Margalef指数变化显著 (P< 0.05)(图 4B),而 Simpson 指数变化未达到显著性水平(P> 0.05)(图4A);此外,随着鼠洞密度的增加,地上生物量却呈现显著增加的趋势 (P< 0.05)(图 4C)。
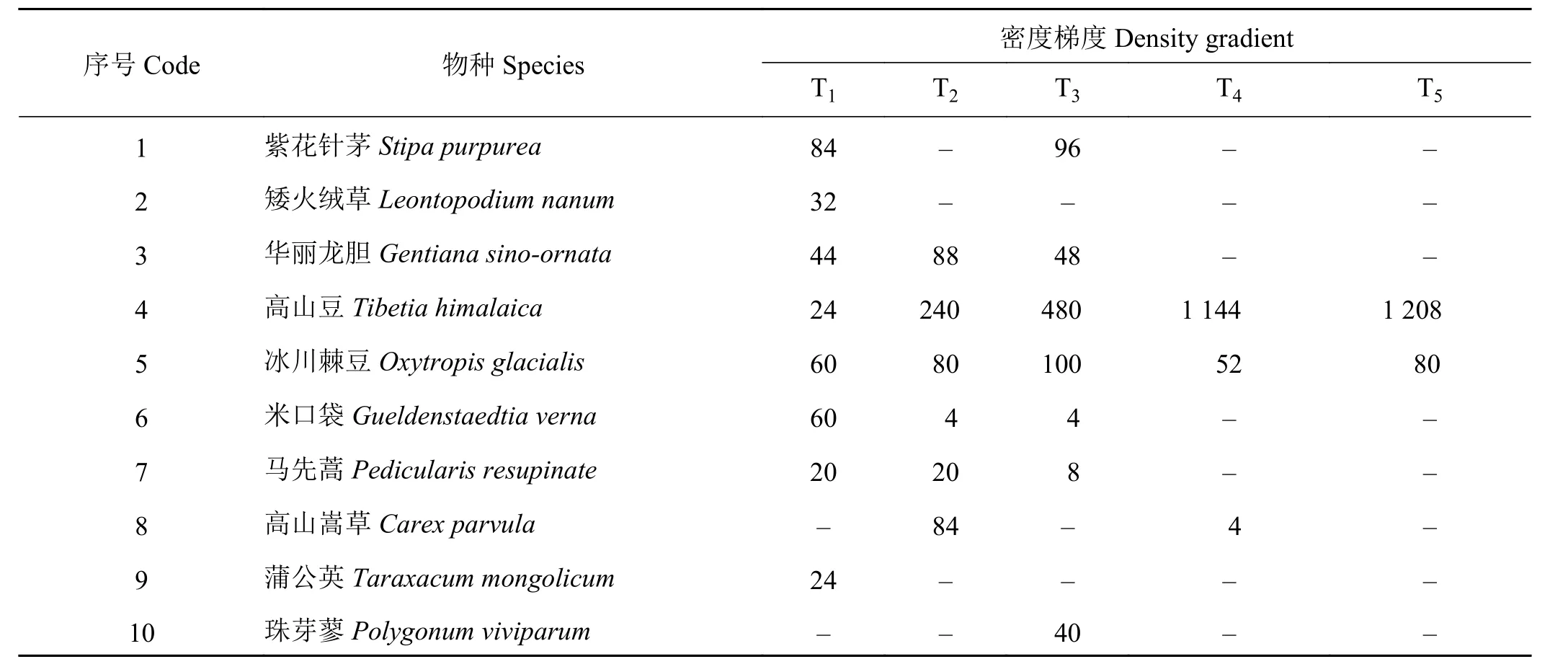
表1 不同鼠兔密度梯度草地植被群落物种数量Table 1 Plant species along an Ochotona curzoniae density gradient plant·m-2
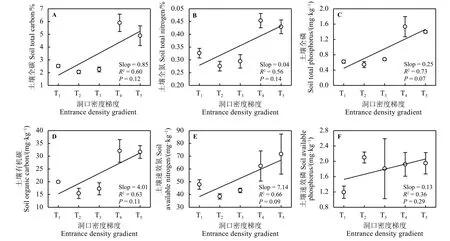
图3 不同高原鼠兔密度干扰对土壤养分的影响Figure 3 Effects of different Ochotona curzoniae densities on concentrations of soil nutrients

图4 不同高原鼠兔密度干扰对植物多样性以及生物量的影响Figure 4 Effects of different Ochotona curzoniae densities on plant diversity (Simpson and Margalef indices)and aboveground biomass
2.4 不同鼠洞密度下植物生物量变化的影响因素
对不同高原鼠兔鼠洞密度草地土壤养分变量和地上生物量进行主成分分析以及热度相关性分析。主成分分析结果发现,具有89.9%的土壤养分驱动变量影响地上生物量的变化(图5A),不同数量鼠兔干扰下土壤驱动因子对地上生物量的变化趋势,其中第一主轴解释了72%,其中土壤速效磷(SAP)驱动变量的第一主成分系数小于0.02,而其他土壤养分驱动变量的第一主成分系数却均高于0.75;而第二主轴解释了17.9%,其中土壤速效磷(SAP)驱动变量的第一主成分系数高于0.97,而其他土壤养分驱动变量的第一主成分系数却均小于0.20(图5A)。鼠兔数量(PIKA)的增加对土壤全碳(STC)、土壤全氮(STN)、土壤全磷(STP)、土壤有机质(SOC)以及土壤速效氮 (SAN)具有显著促进作用 (P< 0.05)(图 5B),不同数量鼠害梯度的土壤全碳(STC)、土壤全氮(STN)、土壤全磷(STP)、土壤有机质(SOC)以及土壤速效磷(SAP)对其地上生物量(AGB)具有显著的促进作用 (P< 0.05)(图 5B)。
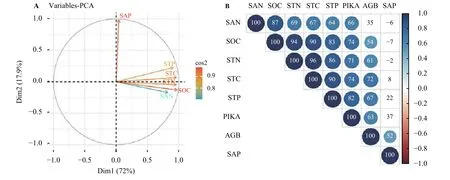
图5 不同高原鼠兔鼠洞密度下植物地上生物量变化驱动力Figure 5 Driving forces of aboveground biomass (AGB) of plants under different Ochotona curzoniae density gradients
3 讨论与结论
3.1 鼠兔宜居地的植物及土壤的特征分析
高原鼠兔是青藏高原一种小型洞穴型哺乳动物,对栖息地的选择更倾向于喜食植物较多,植被高度盖度较低,地势高而平坦的草地[30]。本研究针对土壤的物理环境以及植物群落组成,分析影响高原鼠兔对草地生境的倾向性选择的环境因素。结果发现,在一定的范围内,高原鼠兔更倾向于生活在土壤温度较低、含水量较低、土壤空隙率较高的草地(图2)。土壤环境特性是决定高原鼠兔栖息环境优劣的重要因素。首先,土壤温度直接关系到鼠兔的穴居生活热量传递方式,保暖是高原鼠兔选择土壤洞穴的基础条件之一[31-32],使之能够躲避青藏高原冷酷的气候环境。但是本研究发现,随着鼠兔数量多,土壤温度有所降低,但是幅度不大,最大温差在2 ℃范围内(图2)。洞穴土壤的保暖性对鼠兔活动具有重要的决定性作用,适宜的土壤温度更有利于鼠兔的生活。本研究结果可能由于鼠兔的挖掘活动,致使土壤通透性增加,从而使土壤温度有所降低,对鼠兔的正常生活没有太大影响。其次,高原鼠兔是哺乳性动物,土壤含水量低、土壤干燥有利于鼠兔的正常生活,同时也有利于食物的储存,满足其地势高且干燥的选择依据[30-31]。此外,维护生理舒适度也是高原鼠兔选择干燥洞穴的重要原因[31]。土壤容重一方面反映了土壤的松紧程度,另一方面则反映了土壤的空隙率,土壤容重越低,土壤松紧程度越小,越有利于高原鼠兔挖掘复杂的洞穴,越有利于其食物的储存和生存[33-34]。除了土壤条件外,食物也是高原鼠兔选择栖息地必要考虑因素。
高原鼠兔的食物主要来源于双子叶植物的种子、花、叶、茎、根,由于其对植物鲜嫩多汁的器官喜食程度较高,因此,高原鼠兔主要对植物的叶与须根进行采食[31, 33]。本研究发现,高原鼠兔主要偏好于禾本科以及豆科棘豆属植物,其中随着高山豆数量的增多,冰川棘豆数量基本稳定,高原鼠兔数量也随之增多(表1)。分析原因可能是高山豆和冰川棘豆植被高度低,紧贴地面有利于高原鼠兔的活动、行走以及天敌的躲避[35];其次,其他植物的减少,有利于高山豆等肥大根系植物的入侵与生长[36],同时高原鼠兔的食物组成以双子叶植物为主,高山豆(Tibetia yadongensis)和冰川棘豆(Oxytropis glacialis)为双子叶植物,多汁鲜嫩是高原鼠兔最具喜好食物[31, 37]。此外由于高山嵩草(Kobresia pygmaea)、紫花针茅以及华丽龙胆(Gentiana sinoornata)含水量低、纤维含量较高,高原鼠兔对此类植物的喜食程度较低[31, 38]。本研究结果与卫万荣[31]的研究结果基本一致。因此,高寒草原鼠兔的分布与其喜好植物的分布具有一定的关联度。
3.2 高山草地土壤性质以及地上植物生物量对鼠兔洞口数量变化的响应
土壤是植物生长和繁殖的基床,而土壤养分含量决定了植物的生长发育,维持群落生产力以及保障群落稳定性[12]。鼠兔等生物活动的干扰很容易改变土壤养分含量,本研究发现,在一定高原鼠兔数量范围内,鼠兔数量的增加一定程度上会促进土壤养分(碳、氮、磷)的增加(图3)。高原鼠兔对植物根系的啃食,致使植物根部的死亡,植物死亡根系在微生物的分解下,促进土壤碳素含量增加;此外鼠兔的掘洞活动,促使土壤结构疏松,土壤空隙率增加[39-40],有利于微生物的分解活动,促进有机质分解产生有机碳[12, 39]。高寒草原土壤氮素主要来源于动植物尸体以及动物粪便的分解。高原鼠兔摄食活动,致使植物根系残枝数量增多,同时鼠兔粪便活动,使土壤中氮素的来源增多[41],同时其挖洞活动使土壤的通气性提高,一方面加速了微生物对动植物尸体及鼠兔粪便的分解活动,另一方面促进土壤中硝化细菌数量和活性的提高,加速土壤铵态氮向硝态氮的转化[42-43]。因此,随着高原鼠兔数量的增加,土壤中全氮以及速效氮含量增加,本研究结果与于成等[41]的研究结果基本一致。土壤全磷以及速效磷含量随着鼠兔数量的增加,具有一定的增加趋势。土壤中的磷元素来源于成土母质,以及土壤中有机质的分解[44-45]。本研究中土壤磷素含量的增加,主要因为土壤中动植物尸体,以及鼠兔粪便的增多,从而增加了土壤磷素的来源[44, 46]。此外,在鼠兔的干扰作用下,植物的群落组成以及生物量增加,因此土壤中磷素含量在不同数量的鼠兔梯度下具有一定的差异性[44]。本研究结果与王莹等[46]研究结果基本一致。鼠兔活动对土壤养分产生改变,同时对植物的生长也具有一定的影响。
植被的变化是草地生态系统变化的最直观表现,随着高原鼠兔数量的增多,植物物种多样性降低,地上植物生物量增多(图4)。究其原因,高原鼠兔对禾本科以及棘豆植物的偏好性,致使鼠兔偏食性植物数量减少,杂草类植物增多,从而使植被物种多样性降低[12, 47]。本研究结果与Wei等[47]关于高原鼠兔的研究结果基本一致。另外,分析草地地上生物量的增多的原因,主要有以下几方面:首先,鼠兔的摄食活动、挖掘活动以及粪便活动在增加土壤养分含量的同时,也促进植物对光的可利用性[48]。鼠兔对植物的摄食,致使地上植物稀疏,一方面减缓植物在资源利用上的竞争,另一方面,增加植物对光的利用效率[19, 49]。其次,高原鼠兔的挖掘活动,增加了高寒草原草地土壤异质性,改变了原有植物物种之间的竞争关系,抑制了优势种的生长,促进更多植物的生长[19, 49]。最后,鼠兔摄食和粪便活动致使土壤养分增多,而鼠兔主要对植物的须根进行采食,植物可以通过未损坏的根系,吸收土壤中的养分从而促进其生长[50]。此外本研究发现,速效磷含量的增多,对植物地上生物量变化影响最大(图5)。该结果与Zhang等[48]研究结果相似,表明鼠兔活动对鼠洞周边草地地上生物量具有一定的促进作用。因此在青藏高寒草原,适度的高原鼠兔活动有助于地上生物量增加,但是植物物种多样性会降低。
3.3 结论
本研究通过分析高原鼠兔的草地生境、土壤养分以及植物特征之间的内在联系,结果表明:
1) 高寒鼠害分布主要取决于土壤的水热条件以及植被状况,高原鼠兔更倾向于土壤保暖性好、含水量较低、土壤空隙率较高的草地;植物偏好于棘豆类植物,本研究中主要是高山豆。
2) 随着高原鼠兔干扰强度加大,土壤养分(碳、氮、磷)呈现出一定增加趋势,此外地上生物量也随之增加,但植物群落多样性降低。
综上,在一定鼠兔数量范围内,高原鼠兔活动增加,可以改善高寒草原的土壤营养环境,促进地上生物量的增长。
