海滨雀稗自交结实突变体及野生型幼穗组织的转录组分析
2019-05-23钱晨刘智微钟小仙吴娟子张建丽潘玉梅
钱晨,刘智微,钟小仙*,吴娟子,张建丽,潘玉梅
(1.江苏省农业科学院畜牧研究所, 江苏 南京 210014;2.农业部种养结合重点实验室, 江苏 南京 210014)
禾本科植物的自交不亲和属于配子体自交不亲和,受到2个不连锁的复等位基因座S和Z控制, 比单位点控制的自交不亲和分子机制更为复杂[1]。至今为止, 这2个基因仍然没有被成功克隆[2]。近年来禾本科自交不亲和相关机制研究主要围绕在转录水平开展并取得了明显进展。天蓝虉草(Phalarisarundinacea)的研究中发现了新的自交不亲和控制位点T[3]并利用差异显示的方法中克隆到一个与自交不亲和相关的关键基因:硫氧还蛋白(thioredoxin)基因[4];多年生黑麦草(Loliumperenne)的研究中则通过cDNA-AFLP扩增获得的一些与自交不亲和现象密切相关的蛋白酶类转录衍生片段基因, 如蛋白激酶、肌动蛋白、GTP结合蛋白或泛素相关的蛋白, 其中包括钙依赖受体激酶(calcium-dependent protein kinase, CDPK)以及一些包含有钙离子结合结构域的多肽[5-6]。
随着现代分子生物学研究技术的快速发展, 利用芯片杂交或基于第二代高通量测序技术等可以在全基因组转录水平分析基因表达特征, 对于没有参考基因组的物种, 可以进行denovo转录组测序研究, 这为研究植物自交不亲和反应提供了更有利的条件。近年来这项技术已被应用于植物自交不亲和反应的分子机理研究[7], Zhang等[8]通过比较分析短莛飞蓬(Erigeronbreviscapus)开花组织自花授粉和异花授粉的表达谱, 初步阐述了菊科植物自交不亲和反应的分子机理,同时筛选了240个相关差异表达基因并克隆了7个自交不亲和反应的关键基因。
海滨雀稗(Paspalumvaginatum)作为禾本科多年生草本植物, 具有耐涝、耐旱、耐践踏以及耐盐等特性[9-14], 是优良的草坪草种和牧草资源[15-16], 但由于其自交不亲和特性限制了它的推广应用。获得自交结实的突变体是研究植物自交不亲和反应分子机制的关键环节。由于缺乏自交结实突变体, 海滨雀稗自交不亲和的研究进展缓慢。随着科学技术的进步人们逐渐掌握了创造突变体的各种手段, 利用化学药剂诱发植物产生遗传变异获得自然界没有的或一般常规方法难以获得的新表现型、新突变、新基因, 丰富了遗传变异范畴[17]。钟小仙等[18]以二倍体海滨雀稗品种Adalayd (2n=20, DD)为材料, 通过化学诱变获得了自交结实的体细胞突变体, 细胞学检测发现突变体的染色体数目较野生型没有发生变化(2n=20), 二倍体自交结实型海滨雀稗特异种质的获得, 为研究海滨雀稗自交结实分子机制提供了可能。但由于海滨雀稗基因数据库资源十分匮乏, 限制了相关的研究[19], 本研究以自交结实突变体和野生型为材料, 结合转录组测序方法筛选自交不亲和相关的差异表达基因, 为解析二倍体海滨雀稗自交不亲和分子机制的研究提供了新的思路。
1 材料与方法
1.1 试验材料
试验材料为二倍体自交结实海滨雀稗突变体“SP2008-3”和海滨雀稗品种“Adalayd”, 自交结实海滨雀稗突变体“SP2008-3”由江苏省农业科学院畜牧研究所化学诱变获得。突变体“SP2008-3”自交结实率可达80%以上, 海滨雀稗品种“Adalayd”自交不结实。试验于2013 年9 月在江苏省农业科学院畜牧研究所大田中进行, 选取长势良好、健康、生长期一致植株的5个幼穗组织, 幼穗组织中包含了柱头、子房、花药等植物生殖器官组织。剪取幼穗组织后迅速将其放入纸袋内, 立即用液氮速冻后保存于实验室-80 ℃超低温冰箱中备用。
1.2 RNA提取
利用TRhoL法提取试验材料海滨雀稗幼穗组织的总RNA。提取的RNA样品加入75%乙醇于-80 ℃超低温冰箱中保存备用。
1.3 转录组测序及数据组装
提取样品总RNA, 建立测序文库后用Illumina HiSeq 2000 进行测序。采用Illumina两个末端(PE)测序法, 原始的测序结果去除制备文库时产生的接头序列、两端低质量序列和低度复杂序列, 利用SOAPdenovo软件进行序列拼接, 之后通过连接两末端和填补空位, 将拼接成的重叠群(contig), 进一步组装成Unigene。转录组测序产生的原始序列信息已提交到NCBI的SRA数据库(SRA Bioproject: SRR3747528)。
1.4 功能注释、分类及代谢途径分析
将样本Unigene与公共数据gene进行比较, 通过gene的相似性进行功能注释。基因相似性比对主要基于BLAST算法。使用BLAST程序将拼接得到的 Unigene与核酸、蛋白质数据库进行比对(E值≤1×10-10), 选取最佳的功能注释。核酸数据库为NCBI的非冗余核酸序列数据库(non-redundant nucleotide database, Nt), 蛋白质数据库包NCBI的非冗余核酸数据库(non-redundant protein database, Nr)和蛋白质序列数据库(swissprot protein sequence database,SwissProt)。根据NCBI数据库的功能注释信息, 使用Blast2 GO软件得到Unigene的GO条目, 然后用WEGO软件对所有的Unigene进行GO功能分类统计。然后对Unigene分别进行蛋白质直系同源数据库(cluster of orthologous groups, COG)功能分类和京都基因与基因组百科全书(kyoto encyclopedia of genes and genomes, KEGG)代谢途径分析。
1.5 差异表达富集分析
二倍体自交结实海滨雀稗突变体SP2008-3和野生型两个表达谱之间的差异表达分析, 旨在找出不同样本间存在差异表达的Unigene, 再利用RNA-seq数据比较分析两个样品中同一个Unigene是否存在差异表达。利用错误发现率(false discovery rate,FDR)值的计算方法先要对每个Unigene进行P-value的计算, 再用FDR错误控制法对P-value作多重假设检验校正, fold change是实验组与对照组的差异表达倍数。默认筛选差异的条件为P≤0.05, 本实验设定的fold change是表达相差2倍以上为有意义差异表达基因, 最终筛选出两个表达谱之间的差异表达基因。
2 结果与分析
2.1 海滨雀稗幼穗组织转录组测序及Unigene的功能注释
本研究采用了Illumina Hiseq 2000高通量测序技术对海滨雀稗幼穗组织进行转录组测序, 共得到了68175654个Raw reads片段。denovo拼接共获得了≥200 bp的contig序列208884个, 其中≥500 bp的contig序列有122390个, 而≥1000 bp的contig序列有76934个。在contig数据的基础上, 进一步对序列进行组装, 共获得117619个Unigene序列, 平均长度为717.88 bp, N50为1259 bp;其中≥500 bp的Unigene有47168个, 而≥1000 bp的Unigene序列有26553个(表1)。
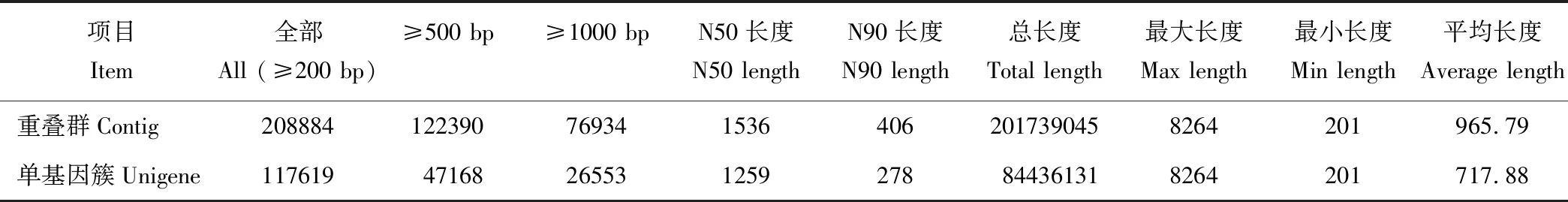
表1 海滨雀稗转录contig和Unigene数据组装统计Table 1 Data assembly for contig and Unigene in the transcriptome of P. vaginatum (bp)
将样本Unigene与公共数据gene进行比较, 通过Unigene的相似性进行功能注释。将Unigene序列, 分别与蛋白质序列数据库SwissProt、翻译核苷酸序列蛋白数据库TrEMBL(translated EMBL nucleotide sequence data library)、保守结构域数据库CDD(conserved domain database)、蛋白家族数据库PFAM(protein families database)、非冗余核酸数据库Nr和真核同源群数据库KOG(eukaryotic orthologous groups)进行比对, 取相似度>30%, 且e<1e-5的进行注释, 合并得到的所有注释详细信息, 注释结果表明有58810个Unigene可在Nr数据库中注释上信息, 占总Unigene数的50%;相似序列匹配的近缘物种中, 高粱(Sorghumbicolor, 53.72%)占的比例最高,之后依次为玉米(Zeamays, 20.64%)、水稻(Oryzasativacv.Japonica, 8.25%)、短柄草(Brachypodiumdistachyon, 3.50%)、另外一个水稻品种(Oryzasativacv.indica, 3.38%)、大麦(Hordeumvulgare1.51%)(图1)。在与蛋白质序列数据库SwissProt、翻译核苷酸序列蛋白数据库TrEMBL、保守结构域数据库CDD、蛋白家族数据库PFAM 4个数据比对后, 有28663、59709、26042、50942个Unigene分别被注释上信息(表2),有近50%的Unigene在数据库中无法被注释上信息。
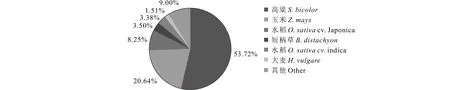
图1 海滨雀稗Unigene的序列Nr数据库相似性分析Fig.1 Characteristics of blast homology search of all assembled Unigenes against the Nr
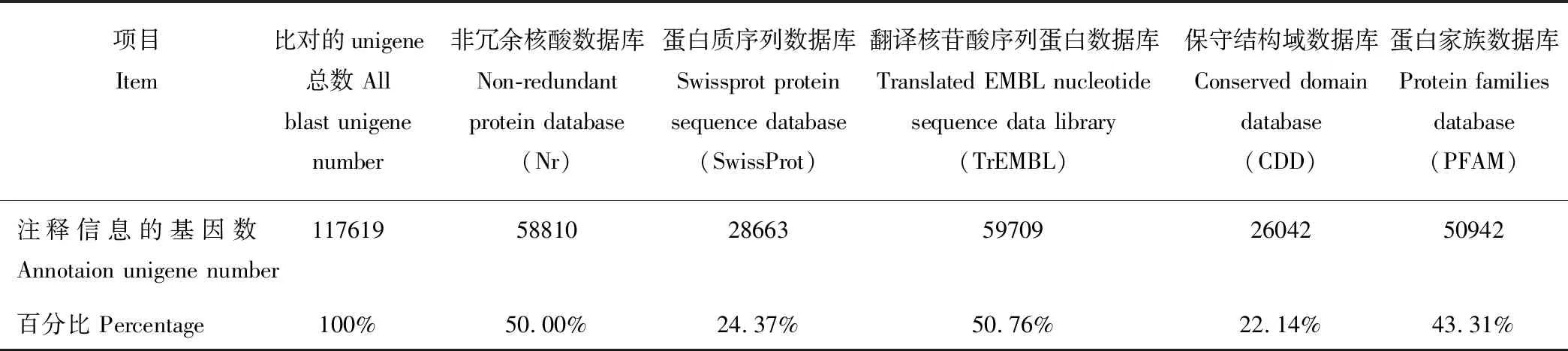
项目 Item比对的unigene总数 All blast unigene number非冗余核酸数据库Non-redundant protein database (Nr)蛋白质序列数据库Swissprot protein sequence database (SwissProt)翻译核苷酸序列蛋白数据库Translated EMBL nucleotide sequence data library (TrEMBL)保守结构域数据库Conserved domain database (CDD)蛋白家族数据库Protein families database (PFAM)注释信息的基因数 Annotaion unigene number1176195881028663597092604250942百分比 Percentage100%50.00%24.37%50.76%22.14%43.31%
2.2 Unigene的分类和代谢途径分析
GO分析表明, 可将海滨雀稗Unigene划分为62个功能组, 并对每个功能组涉及的Unigene进行统计分析(图2)。分析发现19028个Unigene归属细胞组分, 36159个Unigene归属分子功能, 32782个Unigene归属生物学过程。Unigene的功能在生物学过程(biological process)中主要聚集于细胞进程(cellular process)和代谢进程(metabolic process);在细胞组分(cellular component)中主要聚集于细胞成分(cell part)和细胞(cell);在分子功能(molecular function)分类中主要聚集于结合活性(binding activity)和催化活性(catalytic activity)。
对海滨雀稗的Unigene进行COG功能分类, 预测发现共有15835个Unigene被注释上25种COG分类(图3)。Unigene涉及的COG功能类别比较全面, 涉及了大多数的生命活动。其中信号传导机制(signal transduction mechanisms)类的基因最多(3559个)、其次是翻译后修饰, 蛋白质折叠和分子伴侣(posttranslational modification, protein turnover, chaperones)(2966个)、一般功能预测(general function prediction only)(2418个)、翻译, 核糖体结构和生物发生(translation, ribosomal structure and biogenesis)类基因(1615个)、细胞内运转, 分泌和小泡运输(intracellular trafficking, secretion, and vesicular transport)(1533个);细胞运动(cell motility)类基因则是最少的(9个);其他类别的基因所占数量各不相同。
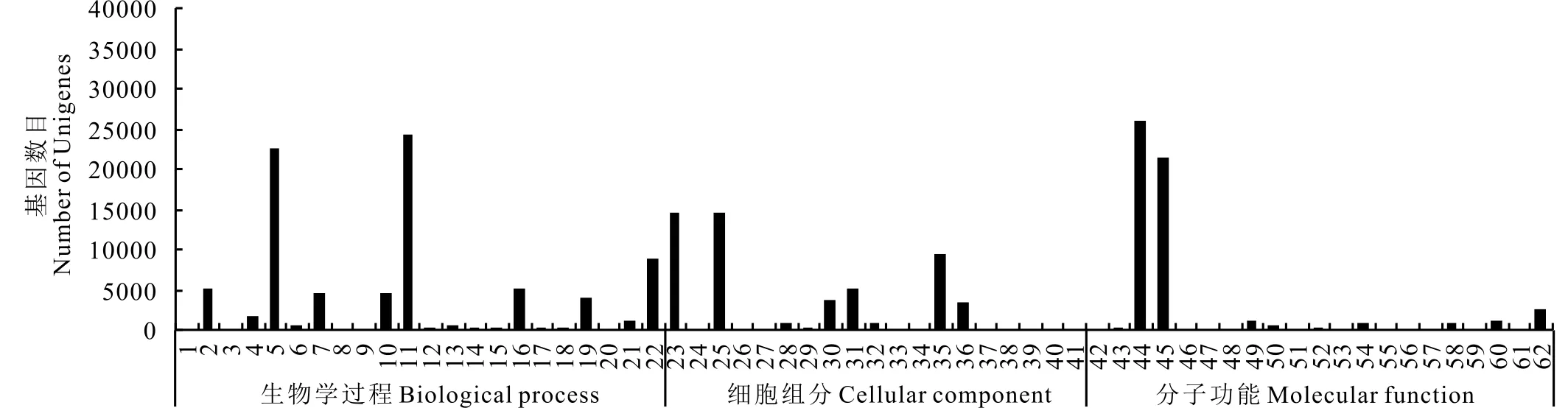
图2 海滨雀稗Unigene的GO分类Fig.2 Gene ontology (GO) classifications of assembled Unigenes 1:生物黏附Biological adhesion;2:生物调节Biological regulation;3:细胞杀伤Cell killing;4:细胞成分组织的生物发生Cellular component orgarization of biogenesis;5:细胞进程Cellular process;6:发育进程Developmental process;7:定位活性Establishment of localization;8:生长Growth;9:免疫系统进程Immune system process;10:定位Localization;11:代谢进程Metabolic process;12:有机体进程Multi-organism process;13:有机体多细胞进程Multicellular organismal process;14:生物进程负调控Negative regulation of biological process;15:生物进程正调控Positive regulation of biological process;16:生物进程的调控Regulation of biological process;17:繁殖Reproduction;18:繁殖进程Reproductive process;19:应激反应Response to stimulus;20:节律进程Rhythmic process;21:信号传导Signaling;22:单组织进程Single-organism process;23:细胞Cell;24:细胞连接Cell junction;25:细胞成分Cell part;26:胞外基质Extracellular matrix;27:胞外基质成分Extracellular matrix part;28:胞外区域Extracellular region;29:胞外区域成分Extracellular region part;30:复杂大分子Macromolecular complex;31:膜成分Membrane part;32:膜关闭内腔Membrane enclosed lumen;33:内质网相关的复杂黏附物Mitochondrion-associated adherens complex;34:类核Nucleoid;35:细胞器Organelle;36:细胞器成分Organelle part;37:共质体Symplast;38:突触Synapse;39:突触成分Synapse;40:病毒Virion;41:病毒成分Virion part;42:D-丙酰胺载体活性D-alanyl carrier activity;43:抗氧化活性Antioxidant activity;44:结合活性Binding activity;45:催化活性Catalytic activity;46:通道调控活性Channel regulation activity;47:细胞趋化活性Chemoattractant activity;48:化学抗性活性Chemorepellent activity;49:电子传递体活性Electron carrier activity;50:酶调节活性Enzyme regulator activity;51:金属伴侣活性Metallochaperone activity;52:分子转导活性Molecular transducer activity;53:形态发生素活性Morphogen activity;54:结合核酸转录活性Nucleic acid binding transcription factor activity;55:营养库活性Nutrient reservoir activity;56:结合蛋白转录活性Protein binding transcription factor activity;57:蛋白标签Protein tag;58:受体活性Receptor activity;59:受体调控活性Receptor regulator activity;60:结构分子活性Structural molecule activity;61:翻译调节活性Translation regulator activity;62:转运活性Transporter activity.
2.3 自交不亲和相关的差异表达基因的筛选及分析
2.3.1差异表达基因的筛选 对海滨雀稗测序获得的Unigene进行功能注释后, 筛选自交结实海滨雀稗突变体SP2008-3与对照野生型Adalayd之间的显著性差异表达基因。本研究中, 差异表达基因条件是差异表达的倍数≥2(即 |log2Ratio|≥1)且 FDR≤0.001 的基因,比较两个表达谱发现SP2008-3较Adalayd共有差异表达基因1303个,其中上调表达基因373个,下调表达基因930个(图4)。
2.3.2差异表达基因的GO分析 通过比较SP2008-3和Adalayd两个表达谱获得1303个差异表达的Unigene之后, 对差异表达Unigene进行GO分析, 对其功能进行描述。 统计每个GO条目中所包括的差异Unigene个数, 并用超几何分布检验方法计算每个GO条目中差异表达的Unigene富集的显著性(图5)。可以根据GO分析的结果结合生物学意义挑选用于后续研究的Unigene。Unigene的功能在生物学过程(biological process)中主要聚集于应激反应(response to stress)和光合作用(photosynthesis);在细胞组分(cellular component)中主要聚集于细胞壁(cell wall)和叶绿体(chloroplast);在分子功能(molecular function)分类中主要聚集于果胶质酶活性(pectinesterase activity)和酶抑制剂活性(enzyme inhibitor activity)。
2.3.3差异表达基因的KEGG分析 将1303个差异表达基因的注释信息与KEGG中酶注释的关系文件进行分析从而映射到不同的通路中, 分析结果总共获得了147张通路图, 如图6列出了富集, 其中戊糖和葡萄糖醛酸盐相互转化(pentose and glucuronate interconv-ersions)和光合天线蛋白(photosynthesis-antenna proteins)富集的差异表达基因最多。分别将上调基因和下调基因进行KEGG归类, 分析发现930个下调基因归类出106个通路, 其中归为淀粉和蔗糖代谢途径的差异表达基因最多, 共有31个;其次是葡萄糖醛酸转换途径, 共有27个差异表达基因归入其中;再次是植物和病原菌互作途径, 共有20个基因归入其中。而将373个上调基因进行KEGG功能分析发现, 共归纳出86张通路图, 其中归为内质网蛋白加工途径的差异表达基因最多, 共有23个;其次是光和生物固碳, 共有20个差异表达基因归于其中;再次是谷胱甘肽代谢途径, 共有12个差异表达基因归入其中。
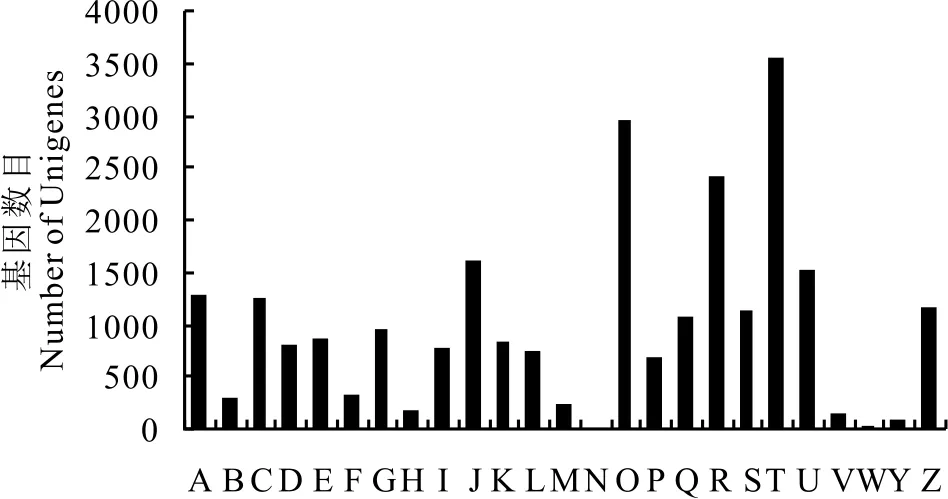
图3 海滨雀稗Unigene的COG功能分类Fig.3 Histogram of clusters of orthologous groups (COG) classification A:RNA加工和修饰 RNA processing and modification;B:染色质结构和活力 Chromatin structure and dynamics;C:能量生成和转换 Energy production and conversion;D:细胞周期控制, 细胞分裂, 染色体分区 Cell cycle control, cell division, chromosome partitioning;E:氨基酸运输和代谢 Amino acid transport and metabolism;F:核苷酸运输和代谢 Nucleotide transport and metabolism;G:碳水化合物运输和代谢 Carbohydrate transport and metabolism;H:辅酶运输和代谢 Coenzyme transport and metabolism;I:脂质运输和代谢 Lipid transport and metabolism;J:翻译, 核糖体结构和生物发生 Translation, ribosomal structure and biogenesis;K:转录 Transcription;L:复制, 重组和修复 Replication, recombination and repair;M:细胞壁/薄膜/膜的生物发生 Cell wall/membrane/envelope biogenesis;N:细胞运动 Cell motility;O:翻译后修饰, 蛋白质折叠和分子伴侣 Posttranslational modification, protein turnover, chaperones;P:矿脂运输和代谢 Inorganic ion transport and metabolism;Q:次生代谢物合成, 运输和代谢 Secondary metabolites biosynthesis, transport and catabolism;R:一般功能预测 General function prediction only;S:功能未知 Function unknown;T:信号传导机制 Signal transduction mechanisms;U:细胞内转运, 分泌及小泡运输 Intracellular trafficking, secretion, and vesicular transport;V:防卫机制 Defense mechanisms;W:胞外结构 Extracellular structures;Y:核结构 Nuclear structure;Z:细胞骨架 Cytoskeleton.
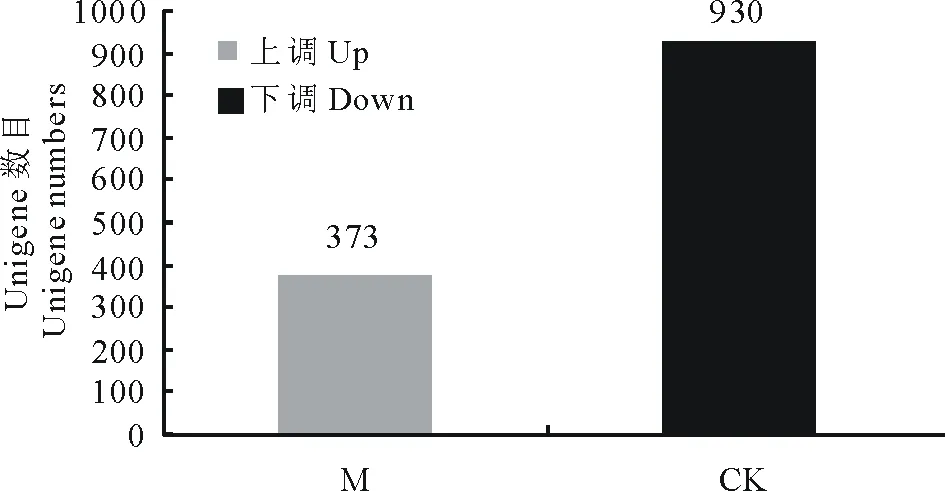
图4 海滨雀稗SP2008-3突变体与Adalayd的差异表达基因Fig.4 Differentially expressed genes in self-compatibility mutant SP2008-3 and Adalayd M: SP2008-3突变体Self-compatibility mutant SP2008-3; CK: Adalayd.
2.4 二倍体海滨雀稗自交结实相关的分子事件
通过比较分析测序基因在自交结实突变和自交不亲和Adalayd对照中的表达量差异, 结合前期研究中发现的与植物自交不亲和相关的通路或基因, 对1303个差异表达基因进行筛选。本研究获得的数据中筛选出了3类与二倍体海滨雀稗自交不亲和相关的通路或基因, 包括14个钙离子通路基因, 3个F-box基因和5个硫氧还蛋白基因(表3)。
2.4.1钙信号通路差异表达基因 本研究通过比较突变体和Adalayd的表达谱发现, 其中有一个钙调素基因(comp36284) 在Adalayd中发生下调表达;其中有8个钙依赖蛋白激酶(comp53237, comp37823,comp24441, comp61028,comp61403, comp58623,comp10880, comp9941)在Adalayd中发生下调表达;5个类钙调素互作蛋白激酶基因(comp63471, comp10442, comp8817, comp8819, comp49346), 其中有3个类钙调素互作蛋白激酶基因(CBL-interacting protein kinase, CIPK)基因(comp8817, comp8819, comp49346)在Adalayd中发生下调表达, 2个CIPK基因(comp63471, comp10442) 则在突变体中发生上调表达。进而对14个钙离子通路基因的KEGG归类发现其中钙调素基因(comp36284)归为植物和病原菌互作途径, 8个钙依赖蛋白激酶则共同归入了toxoplasmosis途径中, 而5个类钙调素互作蛋白激酶基因并未在KEGG数据库中找到相应的功能途径。对14个钙离子通路基因的GO分析发现8个钙依赖蛋白激酶基因中有7个基因(comp53237, comp24441, comp61028, com61-403, comp58623, comp10880, comp9941)在生物学过程(biological process)中归为蛋白质磷酸化, 一个钙依赖蛋白激酶基因(comp37823)和一个钙调素基因(comp36284) 仅知其为钙离子途径基因, 参与的生物学过程尚且未知;5个类钙调素互作蛋白激酶基因中3个类钙调素互作蛋白激酶基因(comp8817, comp8819, comp49346)在生物学过程(biological process)中归为蛋白质磷酸化和信号传导, 另2个类钙调素互作蛋白激酶基因(comp63471, comp10442)的生物学过程未知。由此可初步判断钙信号途径中多数基因是通过蛋白磷酸化的方式参与到了二倍体海滨雀稗自交不亲和途径中, 其中涉及了钙信号的传导, 传导的过程主要是蛋白激酶参与其中。
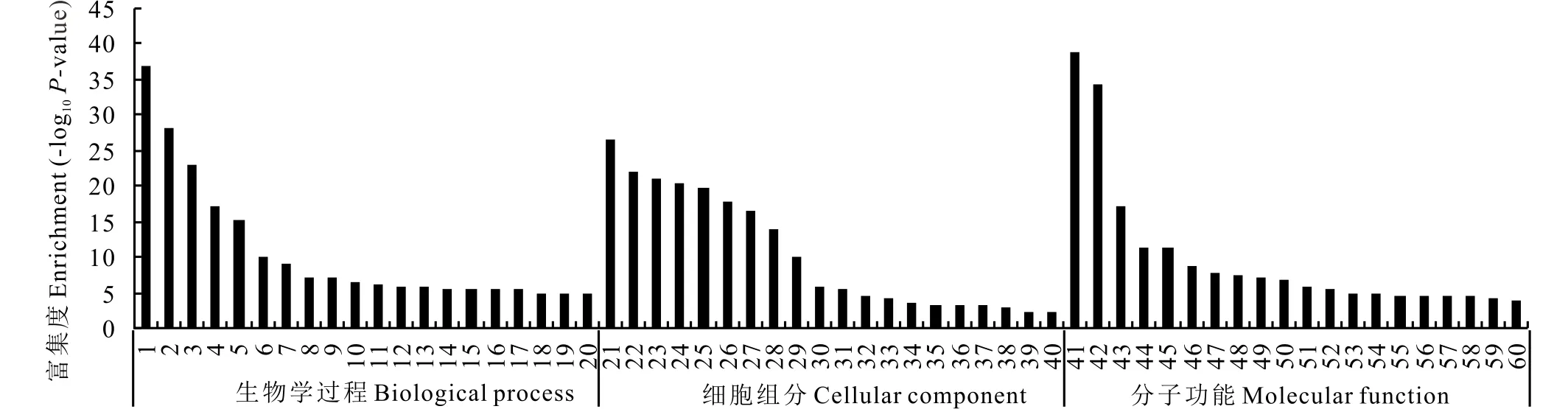
图5 按照参与的生物学过程、分子功能、细胞组分标准差异表达基因的GO分类结果Fig.5 Classfication of differentially expressed genes according to biological process, molecular function, cellular component 1: 应激反应Response to stress;2: 光合作用Photosynthesis;3: 催化活性负调控Negative regulation of catalytic activity;4: 细胞壁修饰Cell wall modification;5: 光合作用的光能捕获Photosynthesis light harvesting;6: 五碳糖磷酸还原循环Reductive pentose-phosphate cycle;7: 植物形态的细胞壁组织Plant-type cell wall organization;8: 染色体蛋白的连接Protein-chromophore linkage;9: 脂运输Lipid transport;10: 糖运输Carbohydrate transport;11: 硫同化Sulfate assimilation;12: 果实催熟Fruit ripening;13: 过氧化氢分解代谢进程Hydrogen peroxide catabolic process;14: 固氮作用Nitrogen fixation;15: 水反应Response to water;16: 有性生殖Sexual reproduction;17: 噬菌作用Phagocytosis;18: 谷氨酰胺生物合成过程Glutamine biosynthetic process;19: 多糖分解代谢进程Polysaccharide catabolic process;20: 乙烯生物合成过程Ethylene biosynthetic process;21: 细胞壁Cell wall;22: 叶绿体Chloroplast;23: 光系统 Ⅰ Photosystem Ⅰ;24: 胞外区Extracellular region;25: 质体Plastid;26: 叶绿体类囊体膜Chloroplast thylakoid membrane;27: 类囊体Thylakoid;28: 光系统 Ⅱ Photosystem Ⅱ;29: 光系统Ⅰ反应中心Photosystem Ⅰreaction center;30: 叶绿体膜Chloroplast membrane;31: 叶绿体基质Chloroplast stroma;32: 液泡Vacuole;33: 细胞外间隙Extracellular space;34: 膜外Extrinsic to membrane;35: 放氧符合体Oxygen evolving complex;36: 叶绿体类囊体腔Chloroplast thylakoid lumen;37: 类囊体膜Thylakoid membrane;38: 膜Membrane;39: 细胞质Cytoplasm;40: 液泡膜Vacuolar membrane;41: 果胶质酶活性Pectinesterase activity;42: 酶抑制剂活性Enzyme inhibitor activity;43: 天冬氨酰酯酶活性Aspartyl esterase activity;44: 果胶裂解酶活性Pectate lyase activity;45: 叶绿素结合Chlorophyll binding;46: 脂结合Lipid binding;47: L-抗坏血酸过氧化物酶活性L-ascorbate peroxidase activity;48: 裂解酶活性Lyase activity;49: 钙调蛋白结合Calmodulin binding;50: 谷胱甘肽转移酶活性Glutathione transferase activity;51: 外切多聚半乳糖醒酸酶活性Galacturan 1,4-alpha-galac-turonidase activity;52: 1-氨基环丙烷-1-羧酸氧化酶活性1-aminocyclopropane-1-carboxylate oxidase activity;53: 谷氨酸氨连接酶活性Glutamate-ammonia ligase activity;54: 过氧化物酶活性Peroxidase activity;55: 磷酸甘油酸激酶活性Phosphoglycerate kinase activity;56: L-抗坏血酸结合L-ascorbic acid binding;57: β-淀粉酶活性Beta-amylase activity;58: 果糖二磷酸醛缩酶活性Fructose-bisphosphate aldolase activity;59: 甘油醛-3-磷酸脱氢酶Cytosolic glyceraldehyde-3-phosphate dehydrogenase;60: 碳酸盐脱水酶活性Carbonate dehydratase activity.
2.4.2F-box基因 F-box蛋白基因被证实在S-RNase调控的植物自交不亲和反应中发挥了关键的作用, 在本研究中发现有3个F-box基因在结实突变体和自交不亲和Adalayd的表达谱中存在差异表达, comp37969, comp38061 和comp45767这3个基因在突变体中较Adalayd发生了显著的下调表达, 其中comp37969 和comp38061较comp45767发生了更为显著的下调表达, 由此判断F-box基因可能参与了二倍体海滨雀稗的自交不亲和反应。
2.4.3硫氧还蛋白 (thioredoxin)基因 本研究发现有5个硫氧还蛋白基因在结实突变体和自交不亲和Adalayd的表达谱中存在差异表达, 其中2个硫氧还蛋白基因(comp62783, comp62777)在突变体中较Adalayd发生了上调表达, 而有3个硫氧还蛋白基因(comp34593, comp20916, comp63644)在突变体中较Adalayd发生了下调表达, 对5个硫氧还蛋白基因的GO分析发现4个硫氧还蛋白基因(comp62783,comp62777,comp34593, comp63644)在细胞组分中归为细胞质, 生物学过程(biologica process)中归为甘油醚代谢过程及运输, 1个硫氧还蛋白基因(comp20916)则在细胞组分中归为细胞质, 在生物学过程(biological process)中归为色氨酸的代谢过程及运输, 基因的差异表达表明硫氧还蛋白基因可能也参与了二倍体海滨雀稗的自交不亲和反应, GO分析获悉5个硫氧还蛋白基因作用于细胞的细胞质中, 而不同表达量以及不同的生物学过程则需进一步研究。
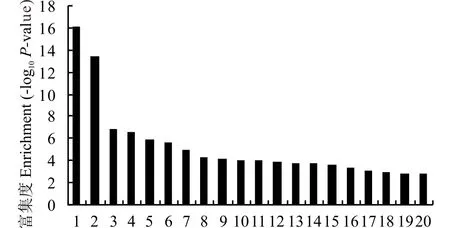
图6 1303个差异表达基因的KEGG分类结果Fig.6 Classfication of differentially expressed genes according to KEGG database 1:戊糖和葡萄糖醛酸盐相互转化Pentose and glucuronate interconversions;2:光合天线蛋白Photosynthesis-antenna proteins;3:光合生物中的碳固定Carbon fixation in photosynthetic organisms;4:光合作用Photosynthesis;5:淀粉和蔗糖的代谢Starch and sucrose metabolism;6:弓形虫病Toxoplasmosis;7:抗原加工和提呈Antigen processing and presentation;8:植物和病原菌互作Plant-pathogen interaction;9:谷胱甘肽代谢Glutathione metabolism;10:化学致癌Chemical carcinogenesis;11:抗坏血酸与醛酸代谢Ascorbate and aldarate metabolism;12:细胞色素p450对外源化合物的代谢Metabolism of xenobiotics by cytochrome p450;13:内质网蛋白加工Protein processing in endoplasmic reticulum;14:硫代葡萄糖苷的生物合成Glucosinolate biosynthesis;15:细胞色素p450的药物代谢Drug metabolism-cytochrome p450;16:类NOD受体信号通路NOD-like receptor signaling pathway;17:半胱氨酸和蛋氨酸代谢Cysteine and methionine metabolism;18:硫代谢Sulfur metabolism;19:苯丙氨酸代谢Phenylalanine metabolism;20:甲烷代谢Methane metabolism.
3 讨论
3.1 海滨雀稗穗部的转录组测序为自交不亲和反应的研究提供了信息基础
海滨雀稗相关的分子生物学研究基础相对薄弱, 基因组测序尚未完成, 因此通过经典遗传学手段开展海滨雀稗自交不亲和的分子机制研究十分困难, 随着高通量测序技术的出现, 此技术广泛地应用于很多无参考基因组物种的研究, 为研究海滨雀稗自交不亲和反应的分子机制, 挖掘关键基因提供了新的思路[20]。本研究应用 Illumina 高通量测序技术对海滨雀稗穗部开花组织的转录组进行测序, 目的是为了比较自交结实突变体SP2008-3和自交不亲和Adalayd两个材料间表达谱的差异, 筛选差异表达基因, 同时也为海滨雀稗的分子生物学研究提供穗部的基因组数据信息。本研究的测序结果获得了117619个单基因簇(Unigene),平均长度为717.88, N50为1259 bp, N50的数值越大, 则表现为组装得到的长片断越多, 组装质量就越高。本研究结果表明, 此次序列组装的长度可以满足转录组分析的基本要求。但在本研究中, 由于海滨雀稗尚未完成基因组测序, 因此只有58810 个Unigene(总Unigene的50%)被注释上信息, 这说明有另外50%的Unigene无法预测其功能, 而这些未知功能的基因可能在海滨雀稗自交不亲和反应中发挥作用。但在现有研究基础下, 无法进行筛选, 有待于将来海滨雀稗全基因组测序完成进行深入研究和挖掘。
本研究结合生物信息学理论对海滨雀稗测序结果进行了细致深入的分析, GO、COG以及KEGG分析可以初步将测序获得的Unigene进行注释和归类。贾新平等[21]首次对海滨雀稗的叶部完成了转录组测序分析, 对46169个Unigene进行了GO、COG和KEGG分析, 将本研究结果与其进行初步的比较发现, 在GO分析上结果大致相似;在COG分析结果上存在差异, 在叶部转录组测序的结果中归类于“一般功能预测类基因”的Unigene最多, 而本研究中更多的Unigene归类于“信号传导机制”, 这些差异可能和不同的测序组织部位相关, 本研究的测序分析结果进一步丰富了海滨雀稗不同组织部位的基因组数据, 同时还对自交结实突变体与野生型对照之间的差异表达基因进行了GO、COG和KEGG分析, 为进一步挖掘海滨雀稗穗部自交不亲和的重要表达基因, 开展相关重要基因的克隆及功能验证等研究奠定了数据及理论基础。
3.2 海滨雀稗自交不亲和反应相关的信号通路以及关键基因
比较自交结实突变体和对照之间表达谱的差异, 结合相关的注释信息从1303个差异表达的Unigene中发掘了一批可能与海滨雀稗自交不亲和相关的基因, 其中包括了钙离子通路基因、硫氧还蛋白基因以及F-box基因。
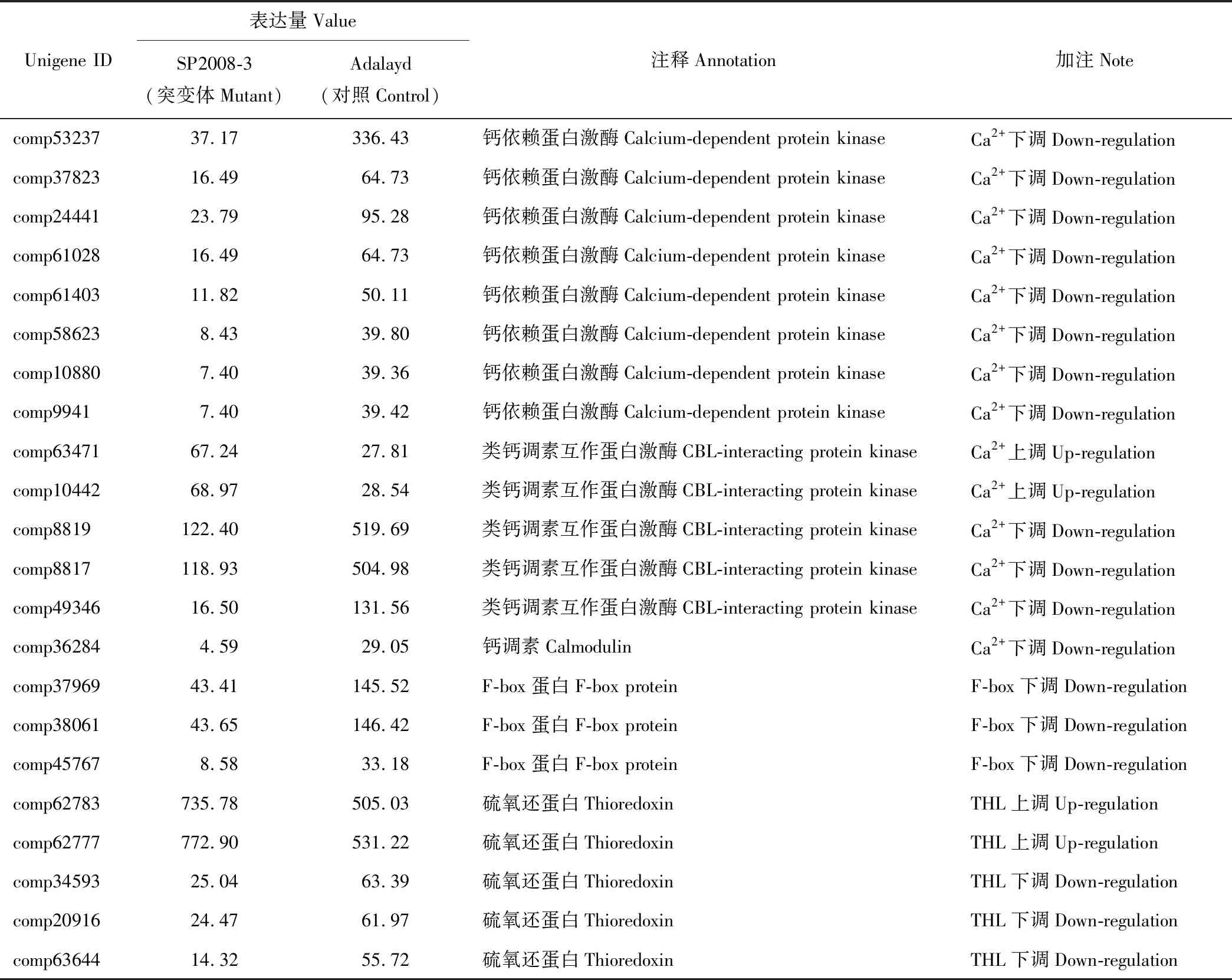
表3 各个通路上的差异表达基因分类及表达特征Table 3 Classification of gene expression pattern in different pathways
3.2.1钙离子信号通路 Ca2+信号通路作为植物细胞的第二信使广泛的参与到了植物的生长发育, 抗病抗逆等生理活动[22], 前期研究发现Ca2+介导了罂粟科、禾本科等一些物种的自交不亲和反应[23-25], Ca2+主要作用体现在Ca2+浓度的升高能够抑制植物花粉管的伸长[26], 在罂粟科虞美人(Papaverrhoeas)自交不亲和的研究中发现Ca2+通过传递信号介导柱头的细胞程序性死亡从而影响花粉管生长造成罂粟科虞美人发生自交不亲和反应[27-28], 而在禾本科裸麦(Secalecereale)的研究中发现蛋白激酶阻遏物和Ca2+通道阻滞剂能够抑制裸麦自交不亲和反应的发生[29], 由此可见Ca2+参与自交不亲和与蛋白激酶相关, 而Klaas等[6]对黑麦草(Loliumperenne)自交不亲和的研究证实了这一点, 研究以自交结实突变体为基础通过对黑麦草不成熟柱头的cDNA-AFLP扩增筛选获得了一个与自交不亲和相关钙依赖受体激酶(CDPK), 以及一些包含有钙离子结合结构域的多肽, 这些研究结果都表明Ca2+作为重要的传导信号参与了植物自交不亲和反应, 并且在参与过程中钙离子信号相关的受体激酶在其中扮演着非常重要的作用, 在本研究中筛选到了1个钙调素基因、8个钙依赖蛋白激酶基因以及5个类钙调素互作蛋白激酶基因在体细胞突变体SP2008-3和野生型Adalayd之间存在差异表达, 结合它们的表达量差异结果, 这些钙离子通路基因可能参与了海滨雀稗自交不亲和反应, 具体的功能正在进一步研究。
3.2.2硫氧还蛋白和F-box蛋白 硫氧还蛋白在禾本科以及十字花科的自交不亲和反应中发挥着重要的作用, 其中禾本科天蓝虉草的自交不亲和研究中发现的关键基因bm2是类硫氧还蛋白, 功能验证发现bm2在天蓝虉草自交不亲和中发挥着非常重要的作用[4], 但bm2在其他禾本科植物中的同源基因并没有发挥类似的作用, 因此类硫氧还蛋白参与禾本科自交不亲和反应存在着物种特异性, 本研究发现在自交结实突变体和对照之间有5个硫氧还蛋白基因存在差异表达, 这5个硫氧还蛋白是否参与了海滨雀稗自交不亲和反应以及具体发挥的功能则需要将来的研究进行验证;F-box蛋白基因是在S-RNase体系中发现在植物自交不亲和反应中发挥作用[30], 尚未有参与禾本科自交不亲和反应的报道, 在体细胞突变体SP2008-3和野生型Adalayd的表达谱比较分析后, 也发现了3个F-box基因存在差异表达, 而这些基因是否参与到了海滨雀稗自交不亲和反应以及其功能则需要将来进一步地验证。
本研究首次将转录组测序技术应用于海滨雀稗自交不亲和的研究中, 建立了海滨雀稗幼穗组织的转录组数据库, 丰富了海滨雀稗的转录本信息, 在此基础上挖掘了一批可能参与海滨雀稗自交不亲和反应的候选基因, 有助于进一步研究海滨雀稗自交不亲和反应, 也为揭示禾本科植物自交不亲和反应的分子机制奠定了重要的基础。
