基于SRK基因序列分析的甘蓝自交不亲和系单倍型鉴定及验证
2018-03-08朱陈曾刘梦慈徐冰冰马存发李勤菲任雪松宋洪元
郑 敏 朱陈曾 刘梦慈 徐冰冰 马存发 李勤菲 任雪松 司 军 宋洪元
(西南大学园艺园林学院,南方山地园艺学教育部重点实验室,重庆市蔬菜学重点实验室,重庆 400715)
甘蓝(Brassica oleraceaL. var.capitataL.)为十字花科芸薹属甘蓝种的一个变种,由不结球的野生甘蓝经长期栽培演化而来,是世界各地广泛栽培的蔬菜种类之一。甘蓝作为典型的异花授粉作物,具有花期自交不亲和的特性,属于孢子体自交不亲和性植物(sporophytic SI,SSI)(Tantikanjana et al.,2010)。自交不亲和系在遗传上由多个复等位基因高度多态性的S位点控制(Takasaki et al.,2000)。S位点基因通常以一个单位遗传给后代,几乎不会发生重组现象,因此S等位基因又称作S单倍型。目前,已经分离到3类与该位点连锁的基因:SRK受体激酶基因(S-locus receptor kinase)(Nasrallah et al.,1985)、SLG糖蛋白基因(S-locus glycoprotein)(Stein et al.,1991)、SCR(S 富 含 半胱氨酸蛋白基因,S-locus cysteine rich)/SP11(S蛋白 11基因,S-locus protein11)(Schopfer et al.,1999)。SLG基因和SRK基因在雌蕊柱头中表达,SCR/SP11基因在雄蕊花药中表达,这3个基因紧密连锁,在花粉、柱头识别过程中起重要作用(Kusaba et al.,2000;Takasaki et al.,2000; 卢军 等,2007),其中SRK基因与SLG基因均呈高度多态性,同一S等位基因系中的SLG基因与SRK基因在序列上协同进化,因此将这两个基因及其连锁序列合称为一种S单倍型(S-haplotype)(Boyes & Nasrallah,1993;Okamoto et al.,2004)。迄今,芸薹属中已有逾100个S单倍型被确定,在甘蓝中已经找到了逾50个S单倍型(Ockendon,2000)。不同S单倍型自交不亲和系之间花期授粉能正常结籽,相同S单倍型之间花期授粉不能结籽或结籽率极低。因此,利用不同单倍型的自交不亲和系进行杂种一代配制是目前甘蓝杂种优势利用的重要途径之一。
目前,甘蓝育种家通过连续自交、小孢子培养等途径育成大量的自交不亲和系,这些在球形、熟性、抗病性等农艺性状表型不同的自交不亲和系存在相同的S单倍型情况。在育种实践中,不同自交不亲和系之间杂交组合配制时除了进行蕾期彻底去雄外,还需同时进行花期杂交,确认参与配组的两个自交不亲和系是否属于同一个单倍型。该方式在一定程度上降低了杂交组合设计的针对性,且田间工作量大。因此,在甘蓝杂交育种工作中进行不同自交不亲和系的单倍型鉴定和分析是非常重要的。早期的自交不亲和系单倍型鉴定和分析方法,如花期亲和指数法(方智远 等,1983)、荧光显微法(张恩慧,1989;陈世儒 等,1990)、免疫测定 法(Kandasamy et al.,1989;Chen & Nasrallah,1990)、等电聚焦电泳法(Nishio & Hinata,1978)等方法存在易受环境条件影响、操作过程繁琐复杂、应用难度较大等问题(高启国 等,2011);基于DNA分子的PCR-RFLP(restriction fragment length polymorphism-polymerase chain reaction,限制性片段长度多态性-聚合酶链式反应)鉴定法受酶切片段多态性的限制难以准确进行多个单倍型的区分(Nasrallah & Nasrallah,1989;朱利泉和王小佳,2000;张爱芬 等,2008);根据SRK激酶区序列设计ClassⅠ型以及ClassⅡ型引物仅能将不同自交不亲和系分为ClassⅠ型和ClassⅡ型,无法确定相应的S单倍型(田磊 等,2011)。随着芸薹属植物自交不亲和性研究的深入,包括甘蓝在内的大量S单倍型所含的SLG、SRK和SCR基因序列的确定以及DNA测序技术的飞速发展,结合生物信息学分析,使得在基因序列水平上快速鉴定芸薹属自交不亲和S单倍型成为可能(李霞 等,2015)。根据SRK基因序列比对,甘蓝自交系所属S单倍型(高启国 等,2011;田磊 等,2011;Tian et al.,2013)以及一代杂种所属的S单倍型可被确定(苗雯雯 等,2013)。然而,基于SRK基因序列分析的单倍型鉴定法是否与传统的花粉管萌发荧光鉴定法、花期亲和指数鉴定法具有一致性仍缺少相应的报道。
本试验针对自交不亲和雌蕊决定因子SRK基因设计简并引物,对23个稳定的甘蓝自交不亲和系SRK基因进行PCR扩增、克隆测序,获得的SRK基因序列进行NCBI数据库的比对分析以及相互之间的序列比较分析,确定相应自交不亲和系的单倍型;并在此基础上进行相同单倍型之间、不同单倍型之间花期杂交的荧光显微法鉴定以及花期亲和指数鉴定,以期验证基于SRK基因序列分析可否快速确定相应甘蓝自交不亲和系所属单倍型,从而实现有计划地设计甘蓝杂交组合,提高育种效率。
1 材料与方法
1.1 试验材料
供试材料为23份稳定的甘蓝自交不亲和系(表1),均由西南大学十字花科蔬菜研究所保存提供。2016年7月育苗,8月定植于西南大学十字花科蔬菜研究所歇马基地。

表1 参试甘蓝自交不亲和系来源及特征特性
1.2 甘蓝基因组DNA的提取
分别取甘蓝自交不亲和系植株幼嫩叶片,利用改良的CTAB法(王关林 等,2014)提取基因组DNA,用1%琼脂糖凝胶电泳检测DNA浓度和纯度,电泳条带单一且分光光度计检测D260/D280比值在1.8~2.0范围内的DNA样品为合格样品,置于-20℃冰箱备用。
1.3 SRK基因引物设计及PCR扩增
根据甘蓝SRK基因序列保守区域设计简并引物:SRK-F:5′-TCAAVCAAYAGAACACTT-3′和SRK-R:5′-CTTYATCCTKGTAAAACCAT-3′(引物中V=A/C/G、Y=C/T、K=G/T),由北京六合华大基因科技有限公司合成。利用该简并引物,以23份甘蓝自交不亲和系DNA为模板,扩增SRK基因片段。PCR扩增体系(25 μL):10×Buffer(含Mg2+)2.5 μL,dNTPs(10 mmol·L-1)0.5 μL,上下游引物(50 μmol·L-1)各 0.1 μL,TaqDNA polymerase(5 U·μL-1)0.2 μL, 模 板 DNA 1.0 μL,ddH2O补齐25 μL。PCR反应程序:95 ℃5 min;95 ℃ 30 s,45 ℃ 30 s,72 ℃ 1 min,30 个循环;72 ℃7 min。获得预期大小的PCR产物,用1.2%琼脂糖凝胶电泳检测,紫外凝胶成像仪系统进行拍照。
1.4 目的片段的回收、克隆及测序分析
利用OMEGA琼脂糖凝胶回收试剂盒,回收目的片段,连接到TRKARA公司的pMD™19-T载体上,转化大肠杆菌感受态细胞DH5α,将获得的克隆进行PCR鉴定。鉴定后的阳性克隆送北京六合华大基因科技有限公司测序。每份自交不亲和系的SRK基因片段测定2~3个克隆。
1.5 甘蓝自交不亲和系S单倍型的确定
将获得的各自交不亲和系的SRK基因序列在NCBI数据库进行Blast比对,确定相应的单倍型。在此基础上利用在线软件Multialin进行相同单倍型以及不同单倍型之间的SRK序列比较分析。根据Tian等(2013)的报道,设计PK1/PK4和KD4/KD7两对引物,进一步对23份自交不亲和系的单倍型(ClassⅠ型、ClassⅡ型)进行鉴定。
1.6 单倍型的验证分析
2017年4月,在西南大学园艺园林学院甘蓝育种基地,根据不同甘蓝自交不亲和系SRK基因序列比较确定的S单倍型,设计20个杂交组合,含相同单倍型、不同单倍型甘蓝自交不亲和系之间的花期、蕾期杂交授粉,并以各自交不亲和系花期和蕾期自交授粉为对照,3次重复,每重复至少50朵花/蕾。
1.6.1 花粉原位萌发的荧光显微镜观察 不同甘蓝自交不亲和系之间花期和蕾期授粉3~5 h后,取其柱头于固定液(冰乙酸∶无水乙醇=9V∶1V的混合液)中固定,24 h后转移至70%的乙醇中保存。固定后的柱头在2 mol·L-1NaOH溶液中55 ℃软化至透明(约2 h),再用50 mmol·L-1偏磷酸钾溶液清洗3~5次,置于0.1%苯胺蓝染液中染色2 h,取出柱头并用50%甘油压片于Leica DM6000B荧光显微镜380 nm激发光下进行花粉管萌发情况观察。每个组合调查至少3个柱头。
1.6.2 亲和指数的测定 完成全部授粉后统计授粉的花朵数,待种荚成熟后逐一考种,计算花期和蕾期杂交亲和指数。3次生物学重复,采用SPSS 19.0软件进行统计分析。
亲和指数=种子数/授粉的花朵数
2 结果与分析
2.1 SRK基因的PCR扩增
利用SRK基因简并引物SRK-F和SRK-R,分别从23份甘蓝自交不亲和系中扩增出预期约
1.0 kb大小的目的片段(图1)。目的片段进行回收,克隆于pMD™19-T载体上,转化大肠杆菌DH5α,对PCR鉴定呈阳性的克隆进行测序分析。
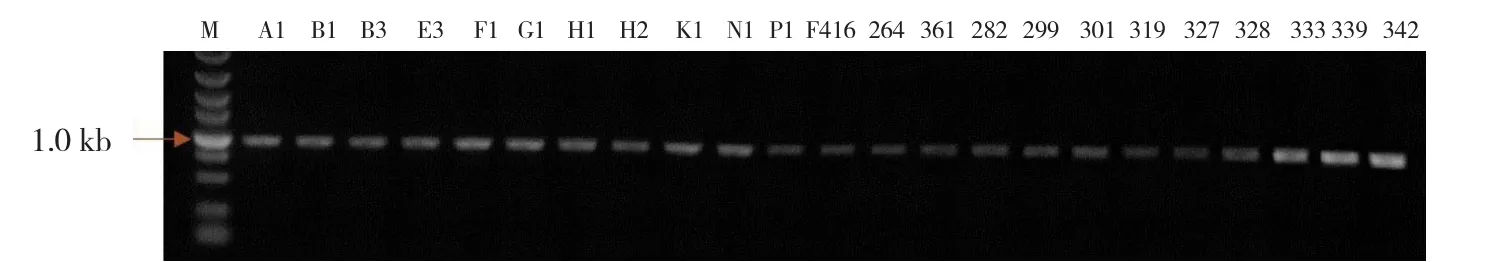
图1 23份甘蓝自交不亲和系SRK基因片段的PCR扩增产物
2.2 甘蓝自交不亲和系单倍型的确定
从表2可以看出,23份甘蓝自交不亲和系的SRK序列均在NCBI库中找到对应的单倍型,其序列一致性在95%~99%之间。如自交不亲和系E3的SRK基因对应NCBI中已知的SRK36单倍型,其序列覆盖率达到99%,序列相似度达到98%,因此将甘蓝自交不亲和系E3确定为SRK36单倍型。其他的自交不亲和系单倍型的确定按类似的方法进行,23份甘蓝自交不亲和系分属10个不同的单倍型,其中属于SRK36单倍型的有5份,占参试甘蓝自交不亲和系的22%;SRK7、SRK16和SRK583种单倍型均只在1份材料中发现。
将属于相同S单倍型的自交不亲和系SRK基因序列以及分属于不同S单倍型的自交不亲和系SRK基因序列进行比较。结果表明:属于同一S单倍型(SRK36)的5个自交不亲和系(282、K1、G1、P1、E3)SRK基因序列同源性极高,仅个别碱基存在差异或个别自交不亲和系发生片段缺失现象,如G1材料的SRK36缺失45 b p序列(图2-a);而分属不同S单倍型自交不亲和系F416(SRK3)、B1(SRK28)、P1(SRK36)、H2(SRK45) 的SRK基因序列差异较大(图2-b)。
进一步针对23份甘蓝自交不亲和系所属的ClassⅠ型以及ClassⅡ型自交不亲性进行鉴定,其中PK1/PK4引物在23份甘蓝自交不亲和系中均扩增获得大小为950 bp的条带,KD4/KD7引物在7 份材料(B3、G1、H2、K1、282、301、339)中扩增获得大小为1 100 bp的条带(图3)。说明除B3、G1、H2、K1、282、301、339外的其他材料属于ClassⅠ型自交不亲和,而7份含1 100 bp条带的材料属于ClassⅠ型和ClassⅡ型杂合材料。
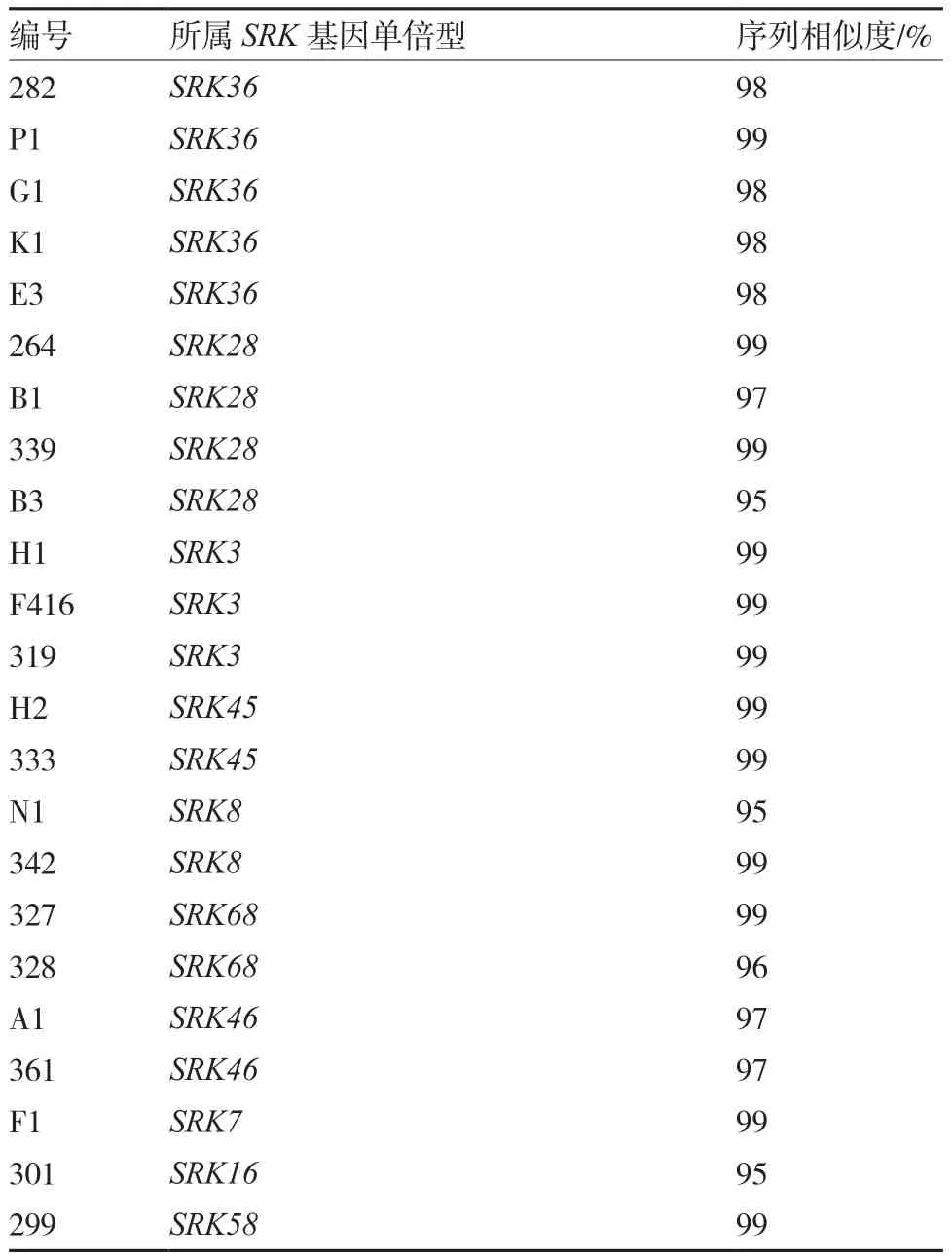
表2 不同甘蓝自交不亲和系所属单倍型

图2 相同单倍型SRK基因序列以及不同单倍型SRK基因序列的多重 比较
2.3 单倍型的验证分析
2.3.1 花粉原位萌发的荧光显微观察 芸薹属作物自交不亲和性表现为开花后自花花粉或相同S单倍型花粉在其柱头上萌发受阻或虽然萌发但花粉管不能正常伸长穿过柱头完成受精。如甘蓝自交不亲和系F416(SRK3单倍型)花期自交授粉后可明显看到花粉粒被吸附在柱头表面但是未形成花粉管束穿过柱头乳突细胞(图4-a),但其蕾期自交授粉后可观察到大量花粉在柱头表面萌发且有部分形成花粉管穿过柱头乳突细胞(图4-b)。相同S单倍型自交不亲和系花期杂交授粉,如F416(SRK3)×H1(SRK3)、G1(SRK36)×P1(SRK36)可见大量花粉被吸附在柱头表面,但是几乎看不到萌发的花粉形成花粉管穿过柱头乳突细胞(图4-c、d)。然而,属于不同S单倍型的自交不亲和系花期杂交授粉后,如F416(SRK3)×E3(SRK36)、H1(SRK3)×H2(SRK45)则可以观察到大量的花粉被吸附在柱头表面萌发并形成花粉管穿过柱头乳突细胞进入花柱组织(图4-e、f)。上述花粉管萌发的荧光检测结果支持了根据SRK序列所确定的不同甘蓝自交不亲和性的单倍型划分结果。
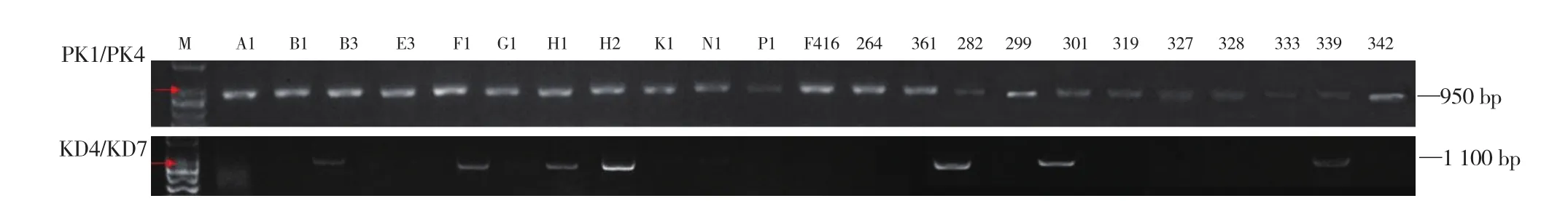
图3 23份甘蓝自交不亲和系ClassⅠ型以及ClassⅡ型自交不亲性鉴定

图4 相同S单倍型以及不同S单倍型甘蓝自交不亲和系花期杂交花粉原位萌发荧光显微观察结果
2.3.2 相同S单倍型以及不同S单倍型甘蓝自交不亲和系花期杂交结荚结籽情况 田间花期授粉35 d后,F416花期自交授粉结荚结籽情况(图5-a~c)与 相 同 S单 倍 型 G1(SRK36)×P1(SRK36)、F416(SRK3)×H1(SRK3)自交不亲和系花期杂交授粉后结荚结籽情况一致,表现为所结种荚短小、干瘪,几乎看不见籽粒突起,空荚占大多数且掉荚现象较为普遍(图5-g~l)。而不同S单倍型F416(SRK3)×E3(SRK36)、H1(SRK3)×H2(SRK45)自交不亲 和系花期杂交授粉后所结种荚长、饱满,籽粒突起明显且种子发育饱满,空荚、死荚极少(图5-m~r),最多种荚结籽数达到34粒,且结籽数量明显优于各S单倍型蕾期自交授粉结荚结籽情况(图5-d~f)。上述结果表明,根据SRK基因序列确定的相同S单倍型自交不亲和系之间花期授粉后不能结籽,表现不亲和;而不同S单倍型自交不亲和系之间花期授粉后结籽量多且饱满,亲和性高。
2.3.3 相同S单倍型以及不同S单倍型甘蓝自交不亲和系杂交组合的花期亲和指数 一般认为,花期亲和指数小于0.5或1的材料属于强自交不亲和系;介于1~3之间为弱自交不亲和系;而大于10的被认定为自交亲和系(方智远 等,1983)。从表3可以看出,自交不亲和系F416花期自交杂交亲和指数为0.09,而蕾期自交平均亲和指数为1.73,属于严格的自交不亲和系。20个杂交组合中,相同S单倍型自交不亲 和系共配制8个杂交组合, 花期杂交亲和指数均小于0.5,其中E3(SRK36)×G1(SRK36)亲和指数最高,但也仅为0.26;且相同S单倍型自交不亲和系之间花期正反交不影响其亲和指数,如E3(SRK36)和G1(SRK36)正反交亲和指数无显著差异。而分属12个不同S单倍型自交不亲和系之间花期杂交结籽多,亲和指数高,最低亲和指数为8.10〔E3(SRK36)×H1(SRK3)〕,最高亲和指数达到21.76〔F1(SRK7)×E3(SRK36)〕。 另 外, 不同S单倍型自交不亲和系之间花期杂交亲和指数差异较大,如H1(SRK3)作为母本分别与H2(SRK45)、E3(SRK36)杂交时,花期亲和指数差异不显著;但E3(SRK36)作为母本分别与H1(SRK3)、F1(SRK7)、N1(SRK8) 杂 交 时,E3(SRK36)×H1(SRK3)的亲和指数显著低于另外两个组合;F1(SRK7)作为母本分别与H2(SRK45)、E3(SRK36)杂交时,亲和指数差异达显著水平。同一自交不亲和系作为母 本与不同单倍型材料杂交后出现结籽数量差异大的现象,可能与来自父本中花粉决定因子SCR/SP11的亲和性有关。上述结果显示,根据SRK基因序列确定的相同S单倍型自 交不亲和系之间花期杂交确实不亲和,而不同S单倍型自交不亲和系之间的花期杂交是亲和的,表明基于SRK基因序列分析确定单倍型可以用于指导不同甘蓝自交不亲和系的杂交组合设计。

图5 相同S单倍型以及不同S单倍型甘蓝自交不亲和系花期杂交结荚结籽情况
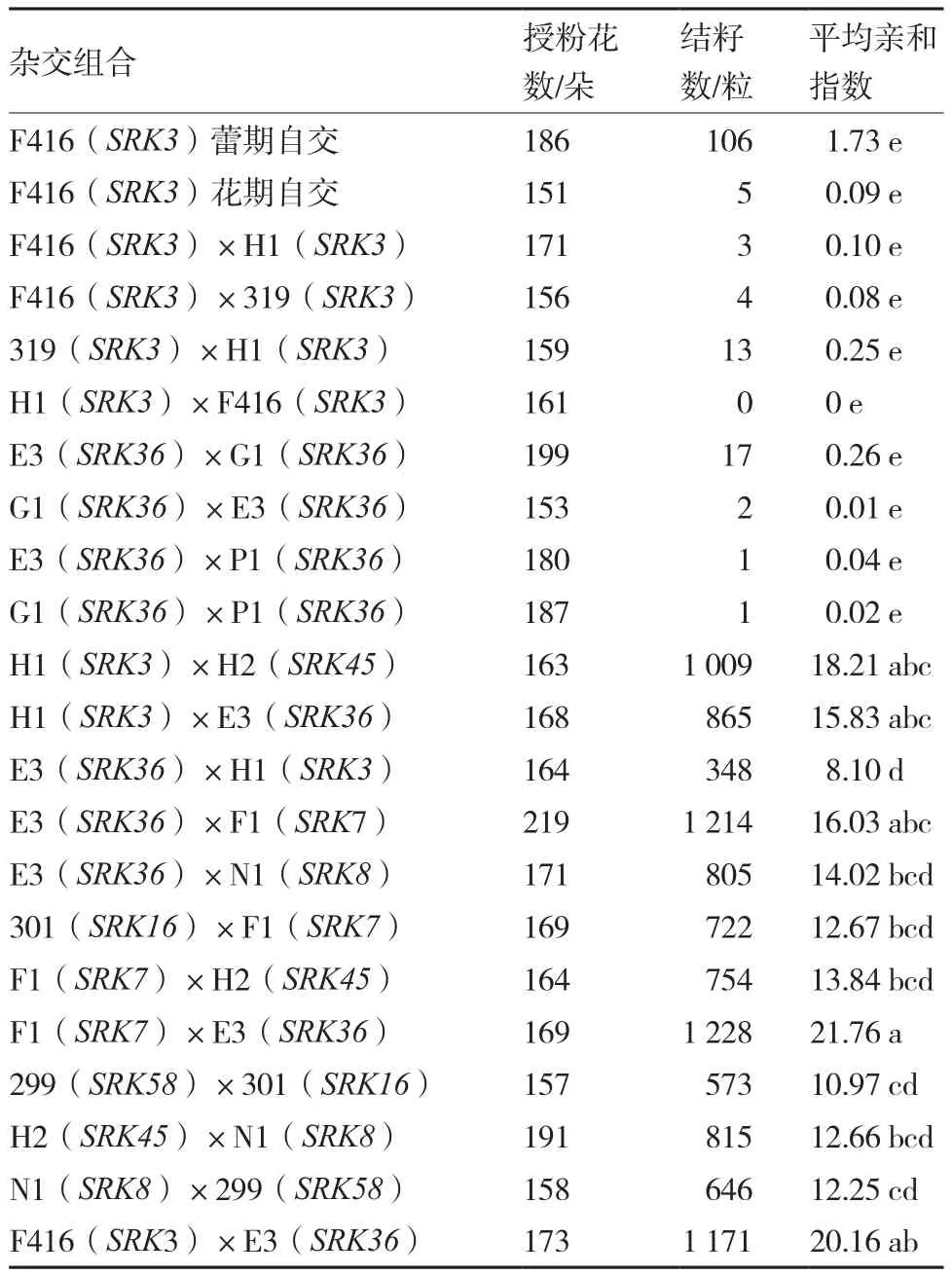
表3 不同甘蓝自交不亲和系杂交组合的花期亲和指数
3 结论与讨论
芸薹属自交不亲和性可分为ClassⅠ型和ClassⅡ型两大类,其中ClassⅠ型为花粉显性,表现强自交不亲和;ClassⅡ型为花粉隐性,表现弱自交不亲和(Nasrallah & Nasrallah,1993)。利用SRK基因特异引物,Tian等(2013)对111份结球甘蓝自交系的单倍型进行鉴定,发现56份属于ClassⅠ型(PK1/PK4引物对),55份属于ClassⅡ型(KD4/KD7引物对);然而,基于花粉管荧光染色鉴定,仅4份属于SRK15单倍型的ClassⅡ型材料表现为花期亲和。在甘蓝育种中,两种类型自交不亲和系均被广泛用于杂种配制(苗雯雯 等,2013)。本试验中,PK1/PK4引物在23份甘蓝自交不亲和系中均扩增获得大小为950 bp的条带,KD4/KD7引物在7份材料(B3、G1、H2、K1、282、301、339)中扩增获得大小为1 100 bp的条带,显示这7份材料在S位点上是杂合的,属于杂合体。甘蓝自交不亲和系存在S位点杂合现象在早期的研究中也曾报道(田磊 等,2011)。然而,通过SRK基因序列分析,未能在这7份杂合材料中鉴定出属于ClassⅡ型的SRK2、SRK5以及SRK15单倍型序列(Tian et al.,2013)。另外,本试验将S位点杂合的G1(鉴定为SRK36单倍型)分别作为父本和母本与同一S单倍型的E3以及P1花期杂交后均表现为典型的自交不亲和特性。说明,即使在S位点上出现ClassⅠ型和ClassⅡ型杂合,仍不影响其花期杂交的不亲和性。
通过特异引物扩增仅能将甘蓝自交不亲和系分为ClassⅠ型和ClassⅡ型两大类,但不能确定其所属单倍型(田磊 等,2011)。李霞等(2015)对4份不结球白菜的SRK及SLG基因序列进行克隆分析,初步确定了其所属单倍型,但未进行相应的亲和性试验。基于SRK基因序列分析,Tian等 (2013)对111份结球甘蓝单倍型进行了鉴定,并利用柱头花粉管荧光鉴定法重点验证了归属于相同S单倍型材料之间亲和性。本试验中,23份甘蓝自交不亲和系分属于10个不同的单倍型,未检测到属于ClassⅡ型的SRK2、SRK5以及SRK15单倍型,但不同单倍型的分布频率与Tian 等(2013)在111个结球甘蓝自交系上的鉴定结果接近;F1材料被鉴定为SRK7单倍型,与高启国等(2011)针对该材料SRK基因和SCR基因同时鉴定结果一致。说明利用本试验中的简并引物扩增SRK基因序列进行单倍型的确定是可靠的。本试验还对归属于相同S单倍型材料以及不同S单倍型材料之间的亲和性,同时采用柱头花粉管荧光鉴定法以及亲和指数测定法进行了验证,获得了基于SRK基因序列鉴定相应自交不亲和系所属单倍型与田间鉴定相吻合的直接证据。这些结果支持基于SRK基因序列分析确定对应甘蓝自交不亲和系所属单倍型是一种简单可靠的鉴定方法,可用于指导甘蓝自交不亲和系之间的杂交组合设计。
陈世儒,雷建军,李成琼,郭余龙.1990.甘蓝自交不亲和性的荧光染色快速测定.中国园艺学会成立六十周年纪念暨第六届年会论文集.北京:万国出版社.
方智远,孙培田,刘玉梅.1983.甘蓝杂种优势利用和自交不亲和系选育的几个问题.中国农业科学,(3):51-61.
高启国,韦静宜,朱利泉,王小佳.2011.基于SRK和SCR序列的甘蓝自交不亲和系单倍型分析.西南大学学报:自然科学版,33(2):86-90.
李霞,胡侦华,周国林,汪爱华.2015.四个不结球白菜自交不亲和S单元型分子鉴定.湖北农业科学,(16):3948-3951.
卢军,李乐玉,陈晓丹,朱利泉,王小佳.2007.芸薹属自交不亲和功能因子及分子机制研究进展.广西农业科学,(6):601-605.
苗雯雯,田磊,庄木,刘玉梅,杨丽梅,张扬勇,方智远.2013.甘蓝一代杂种S单元型的分布.中国蔬菜,(5):23-28.
田磊,庄木,刘玉梅,杨丽梅,张扬勇,方智远.2011.甘蓝自交系S单元型的快速分类鉴定.园艺学报,38(S):2545.
王关林,方宏筠.2014.植物基因工程.北京:科学出版社.
张爱芬,李英,高素燕,史公军,侯喜林.2008.不结球白菜自交不亲和S单元型的鉴定研究.西北植物学报,28(9):1720-1727.
张恩慧.1989.用荧光显微法测定甘蓝自交不亲和性.陕西农业科学,(1):6-7.
朱利泉,王小佳.2000.利用S位点多态性快速测定甘蓝自交不亲和性及其S等位基因系.植物学报,42(6):595-599.
Boyes D C,Nasrallah J B.1993.Physical linkage of theSLGandSRKgenes at the self-incompatibility locus ofBrassica oleracea.Mol Gen Genet,236:369-373.
Chen C H,Nasrallah J B.1990.A new class of S-sequences defined by a pollen recessive self-incompatibility allele ofBrassica oleracea.Mol Gen Genet,222:241-248.
Kandasamy M K,Paolillo D J,Faraday C D,Nasrallah J B,Nasrallah M E.1989.The S-locus specific glycoproteins ofBrassicaaccumulate in the cell wall of developing stigma papillae.Dev Biol,134:462-472.
Kusaba M,Matsushita M,Okazaki K,Satta Y,Nishio T.2000.Sequence and structural diversity of the S-locus genes from different lines with the same self-recognition specificities minBrassica oleracea.Genetics,154:413-420.
Nasrallah J B,Kao T H,Goldberg M L,Nasrallah M E.1985.Erratum:a cDNA clone encoding an S-locus-specific glycoprotein fromBrassica oleracea.Nature,326(6112):523.
Nasrallah J B,Nasrallah M E.1989.The molecular genetics of selfincompatibility inBrassica.Ann Rev Genet,23:121-139.
Nasrallah J B,Nasrallah M E.1993.Pollen-stigma signaling in the sporophytic self-incompatibility response.The Plant Cell,5(10):1325-1335.
Nishio T,Hinata K.1978.Analysis of S-specific proteins in stigma ofBrassica oleraceaL.by isoelectric focusing.Heredity,38(3):391-396.
Ockendon D J.2000.The S-allele collection ofBrassica oleracea.Acta Hort,539:25-30.
Okamoto S,Sato Y,Sakamoto K,Nishio T.2004.Distribution of similar self-incompatibility(S)haplotypes in different genera,RaphanusandBrassica.Sexual Plant Reproduction,17(1),33-39.
Schopfer C R,Nasrallah M E,Nasrallah J B.1999.The male determinant of self-incompatibilty inBrassica.Science,286:1697-1700.
Stein J C,Howlett B,Boyes D C,Nasrallah M E,Nasrallah J B.1991.Molecular cloning of a putative receptor protein kinase gene encoded at the self-incompatibility locus ofBrassica oleracea.Proceedings of the National Academy of Sciences of the United States of America,88(19):8816-8820.
Takasaki T,Hatakeyama K,Suzuki G,Watanabe M,Isogaik A,Hinata K.2000.The S-receptor kinase determines selfincompatibility inBrassicastigma.Nature,403:913-916.
Tantikanjana T,Nasrallah M E,Nasrallah J B.2010.Complex networks of self-incompatibility signaling in theBrassicaoleracea.Plant Biol,13:520-526.
Tian L,Miao W,Liu J,Fang Z,Liu Y,Yang L,Zhang Y,Zhuang M.2013.Identification of s haplotypes in cabbage inbred lines(Brassica oleraceavar.capitataL.).Scientia Horticulturae,164:400-408.
