Lager啤酒酵母起源的历史足迹及基因组学研究
2019-05-23尹花贺扬侯晓平陈璐董建军
尹花,贺扬,侯晓平,陈璐,董建军
(啤酒生物发酵工程国家重点实验室(青岛啤酒股份有限公司),山东 青岛,266100)
1 悠久的啤酒酿造历史
啤酒是世界上历史最悠久、饮用范围最广的酒精饮料之一,是典型的农业社会发展的产物。啤酒酿造的发展历史与人类社会和文化的发展息息相关,并且受到人类迁徙的影响。据考古学家推测,苏美尔人和巴比伦人早在公元前6 000年开始种植大麦后不久,便开始了原始的啤酒酿造,用于祭祀女神。而公元前3 000年逐步扩展到古波斯、古巴比伦和古埃及等地。在这些社会中,啤酒既是宗教仪式中的重要元素,也是人们日常生活中的常用饮料和药物。而后啤酒传入欧州北部国家,并在每个国家产生了自己特色的品种[2]。斯坦福大学在2016年《美国科学院院刊》(PNAS)的考古研究报道显示,在一个出土于西安米家崖遗址,5 000年前的陶器中发现了用于酿造啤酒的大麦、黍米等作物的残留物[3],表明中国人酿造啤酒的历史同样非常悠久。原始啤酒逐步发展,后来在世界各地酿制出各种各样的啤酒,所采用的原料有所不同,口味也各异。
现代的啤酒酿造技术源于中世纪欧洲的酿造工艺。北欧和英国的温带海洋性气候为种植小麦、燕麦和大麦等谷物提供了理想的条件[4]。谷物中的淀粉在发芽和糖化的过程中能够转变成可发酵性糖(如麦芽糖),随后通过野生酵母的自然发酵生成酒精。但在当时人们对于负责发酵的物质本质仍然难以理解,将其命名为“yeast”。该词来源于德语的“Gischt”或荷兰语的“Gist”,指的是发酵结束时容器顶部的泡沫或絮状物质。酿酒师在每一次发酵结束后将这些“泡沫”刮下,并接种到下一批啤酒的发酵液中。这种在发酵结束后会浮于发酵罐表面的酵母被称为“上面发酵子(top fermenters)”,这实际上就是Ale啤酒(上面发酵啤酒)的酿造[5]。Ale啤酒兴起较早,也被认为和人类早期生产的啤酒比较接近。由于其发酵温度较高,导致产品均一性不好且不易储存,所以啤酒的稳定性和纯净度成了中世纪啤酒酿造的主要问题。
随着啤酒酿造工业的兴起,为控制啤酒的纯净度和价格颁布了一系列的法律法规。1516年在巴伐利亚颁布了《纯酿法》(Reinheitsgebot),规定了啤酒只能在较为寒冷的时间进行酿造(从9月29日的圣米迦勒日到4月23日的圣乔治日之间),且酿造原料只能包括大麦、酒花、水。而法规中并未提及酵母,因为当时酵母被认为是发酵过程的一个副产物而已。而在温度较高的季节,则将啤酒存放在塞满了冰块的山洞中,这一过程则被称为“Lagering”[6],经历了这一储存过程的啤酒则更加稳定。渐渐的人们发现,用这种方式酿造啤酒,在发酵结束后酵母并不会浮在表面,而是沉到发酵罐的底部,因此这些酵母被称为“下面发酵子(bottom fermenters)”。从高温发酵转向低温发酵,并结合低温储存过程,这就是我们今天所熟知的“下面发酵”及Lager啤酒的酿造工艺。
因此,依据发酵工艺和酵母菌种的差异,现代啤酒主要分为两大类:Ale啤酒(上面发酵啤酒)和Lager啤酒(下面发酵啤酒)。Ale啤酒发酵温度较高,口感浓厚,香气丰富,但发酵过程不易控制,产品均一性不好且不易储存,因此在现代的工业啤酒生产中占比不大。与Ale啤酒相比,Lager啤酒发酵温度低,发酵周期长,口味更加淡爽纯净,其发酵过程容易控制,产品均一性好,生产效率高,更适合标准化的工业生产。19世纪中叶,Lager啤酒逐步替代Ale啤酒成为现代工业啤酒的主流,也是全球生产和消量最大的啤酒品类。现代Lager啤酒在19世纪末引入中国,虽然初期发展缓慢,但后期迅速增长,并于2002年超越了美国,连续16年成为全球第一大啤酒生产国。根据啤酒的种类不同,发酵啤酒使用的酵母种类也不同。生产Ale啤酒的酵母即是生物学上重要模式生物——酿酒酵母(Saccharomycescerevisiae,上面发酵酵母,Ale酵母)。而Lager啤酒主要由一种杂交酵母(S.pastorianus、S.carlsbergensis、S.monacensis),统一称为巴氏酵母(S.pastorianus,下面发酵酵母, Lager酵母)发酵而成[7]。
2 Lager酵母起源及遗传学背景
Lager酵母是自然界杂交所产生的异源多倍体菌株。在大规模进行酿酒酵母全基因组测序之前,就已通过单基因序列和微卫星序列的对比分析证实了S.cerevisiae是Lager酵母其中的一个亲本来源[8-9]。
近年来,随着测序技术的发展和测序成本的降低,共有超过1 000株S.cerevisiae的全基因组序列陆续完成测序[10]。通过对工业酿酒酵母菌株(S.cerevisiae)基因组水平的研究,揭示了酵母菌是如何在人类的长期酿造过程中被“驯化”,以及啤酒美味背后的生物学基础。来自比利时鲁汶大学的研究团队搜集了不同来源及工业用途多样的157株工业酿酒酵母菌株,并完成了不同菌株的全基因组测序。通过比较工业酿酒酵母基因组及已发表的酿酒酵母基因组(包括葡萄酒酵母和面包酵母等),进一步构建了157株工业酿酒酵母的系统发生树(图1)[11]。显然,S.cerevisiae可以按照其工业用途(葡萄酒、苹果酒、面包、啤酒、清酒、蒸馏酒、临床、实验室)及地理来源(亚洲、北美、非洲、欧洲等)被区分开。酿造来源的S.cerevisiae形成一个独特的群,其中大部分的啤酒酵母(85.3%)聚集在两个子群(Beer 1和Beer 2),但这两个子群的亲缘关系较远。Beer 1由来自不列颠群岛(英国和爱尔兰)、美国、德国和比利时的S.cerevisiae菌株组成,其中美国来源的菌株与英国菌株在系统发生树上关系较近,表明早期是由英国定居者把酿酒酵母引入了美国。而Beer 2则与葡萄酒酵母的亲缘关系更加密切[11]。
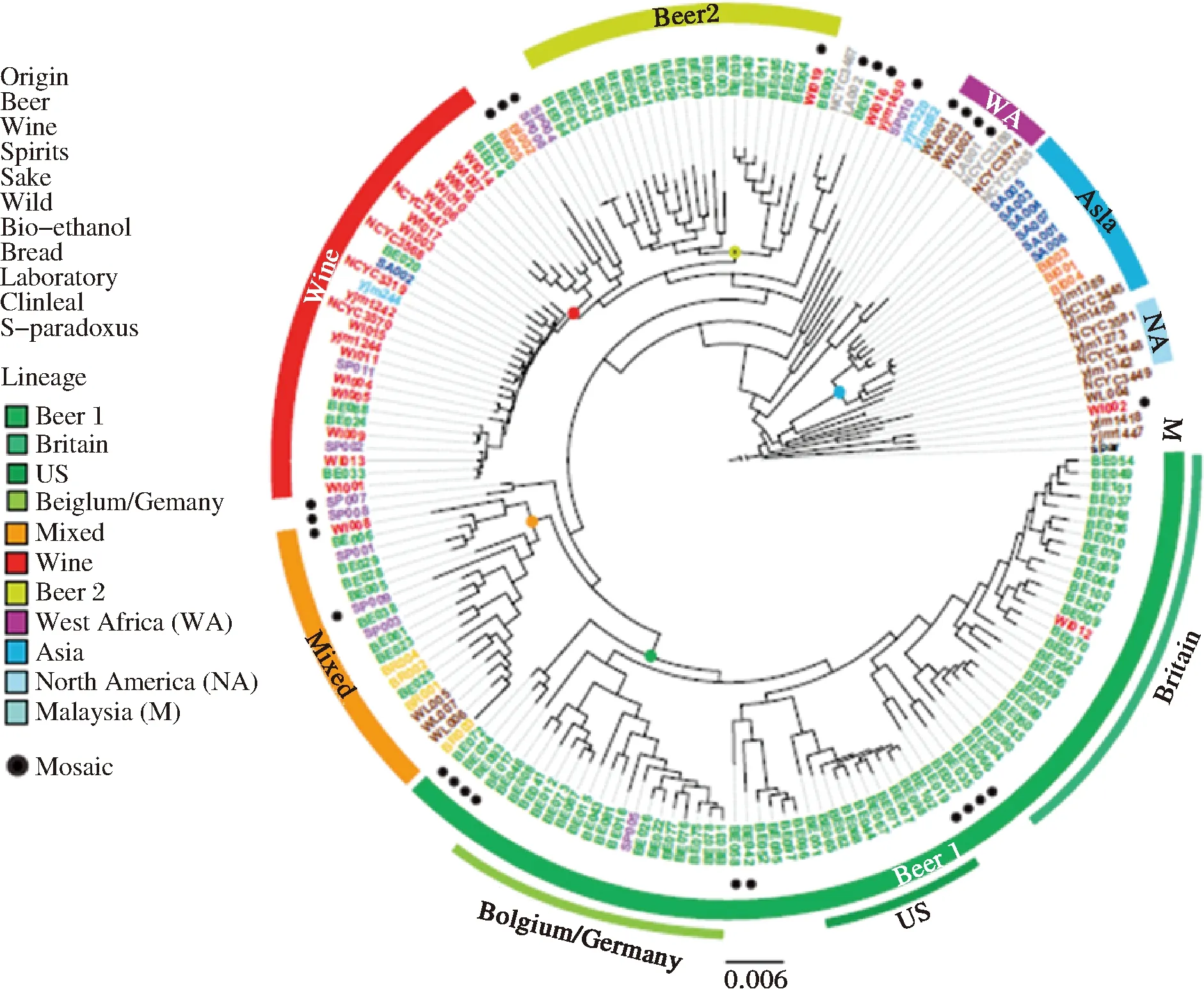
图1 工业酿酒酵母菌株的系统发生树Fig.1 Phylogeny structure of industrial S. cerevisiae strains
比较这两类酵母的分支可以明显看出,啤酒用酿酒酵母,无论是Beer 1还是Beer 2,都有较长的“树杈”,而葡萄酒用酵母的“树杈”则偏短[11]。这表明与葡萄酒酵母相比,啤酒酵母在工业化过程中经历了漫长的驯化,与它们的祖先已经相去甚远。主要表现在工业啤酒酵母的异味生成量降低(4-乙烯基-愈创木酚,4-VG)、麦芽三糖的利用率增加,且在自然条件下的存活率降低。另外,啤酒酵母在长期的工业驯化下几乎都已失去交配能力,而葡萄酒酵母则仍保存了有性生殖的形式[11]。这与两种酒的酿造模式是分不开的。啤酒的酿造条件受季节限制较少,一年四季都可以酿造。人们常常会用上一批啤酒使用过的酵母去酿造下一批啤酒,这无形中就完成了对啤酒酵母的人工驯化。并提出了啤酒酵母的驯化假说:啤酒酵母经历了两次单独的驯化过程,第一次驯化发生在公元1573-1604年间;第二次驯化事件发生在公元1645年至1671年之间[11],这期间正是啤酒酿造从家庭手工业转型成工业化酿造的过程。相比之下,葡萄酒则只能在特定的季节生产。在生产季节外的时间里,尚未掌握微生物学原理和操作技术的中世纪酿造人员只能将酵母留在地窖中,而它们此时则有机会和野生酵母交配,因此展示出的驯化痕迹也较少。
在另一项系统发生研究中,GONÇALVESET等将啤酒S.cerevisiae酵母分为3个进化支:小麦啤酒酵母、英国爱尔兰啤酒酵母和德国Alt-Kolsch啤酒酵母[12]。将Lager酵母S.cerevisiae的亚基因组(Sc-亚基因组)与面包酵母、啤酒酵母、葡萄酒酵母及野外来源的90种S.cerevisiae酵母菌株进行系统发生树分析后,Lager酵母Sc-亚基因组与啤酒S.cerevisiae的亲缘关系更加密切[12]。这些结果都证实了Lager酵母中S.cerevisiae的亲本来源是Ale酵母。
然而,赋予Lager酵母能够在低温进行发酵的非酿酒酵母来源基因序列却一直存在争论。嘉士伯实验室的Morten Kielland-Brand利用单染色体转移的技术将Lager酵母的III号染色体转移至S.cerevisiae中,发现染色体上很大一部分无法与酿酒酵母染色体发生同源重组[13]。这暗示了Lager酵母不是一个简单的酿酒酵母多倍体,而存在除S.cerevisiae外的其他基因来源。直至20世纪末, TAMAI等通过Southern blot杂交技术发现了S.pastorianus有两组不同的染色体,一组跟S.cerevisiae类似,另外一组则与贝氏酵母(S.bayanus)类似[14]。因此长久以来,Lager酵母被认为是S.bayanus与S.cerevisiae杂交的产物[9,15-16]。然而日本三得利的研究学者们在2009年完成的首株Lager酵母全基因组测序结果表明,非S.cerevisiae来源的序列与S.bayanus的同源性仅为92.7%[17],说明Lager酵母可能还存在除真贝氏酵母外的其他亲本来源。此外,研究发现S.bayanus是一个复杂的物种,存在纯合体和杂交体两种形式及至少两种基因型:S.bayanusvar. bayanus和S.bayanusvar. uvarum或S.uvarum[18]。直至2011年首次于南美洲获得了真贝氏酵母(S.eubayanus)后发现,Lager酵母中非S.cerevisiae来源的序列与S.eubayanus的序列相似性达到99.5%[19]。最终明确了工业Lager酵母的亲本来源,是由S.cerevisiae与S.eubayanus通过自然杂交获得。
真贝氏酵母(S.eubayanus)是一种耐寒的野生酵母,主要分布在南美巴塔哥尼亚地区的假山毛榉树和智利南洋杉、亚洲地区的栎树以及北美地区的雪松、红橡树等地,而作为Lager酵母起源地的欧洲则没有发现野生的S.eubayanus[20-22]。S.eubayanus最早(2011年)在南美地区发现,因此提出S.eubayanus是通过欧洲殖民者在新大陆建立的贸易路线传入欧洲的假说[23]。近年来又在中国、北美和新西兰均分离到野生的S.eubayanus菌株[20-24],因此作为Lager酵母亲本之一的S.eubayanus起源地则变得更加复杂。
2014年在中国西藏野生酵母种群及其丰富,涵盖了发现的所有种类,多样性远高于美欧。中国西藏地区发现的S.eubayanus与Lager酵母中非S.cerevisiae来源基因组的序列同源性高达99.8%[23],比来自南美巴塔哥尼亚的S.eubayanus菌株相似性更高。这一结果暗示了来源于西藏的S.eubayanus菌株是Lager酵母中非Ale酵母来源的亚基因组的贡献者。由于Lager发酵起源于1 400年的巴伐利亚,而跨大西洋贸易始于1 500年哥伦布发现新大陆后,因此S.eubayanus从南美通过贸易途径进入欧洲的假说难以成立。而亚欧位于同一大陆,地理位置接近,商贸交往历史悠久。中国古代与欧洲通过丝绸之路的联系一直存在,野生酵母可以通过人员或货物的交流而散播。因此来源于西藏的S.eubayanus被认为是Lager啤酒酵母非Ale基因组真正的野生亲本。通过丝绸之路传播到欧洲,在Lager啤酒低温酿造时与S.cerevisiae进行杂交,赋予Lager酵母耐寒的特性。
3 Lager酵母分型
19世纪的PASTEUR和HANSEN提倡运用纯种发酵来生产Lager啤酒,以避免啤酒的腐败和异味。1909年,LINDNER首次分离了2株不同的Lager酵母菌株,分别以它们的起源地波西米亚(今捷克)Saaz跟德国Frohberg命名,称为Group Ⅰ型(Saaz型)与Group Ⅱ型(Frohberg型)[25]。后来,两种类型的菌株陆续传到欧洲,并最终在世界各地进行使用[26]。Saaz型酵母现主要在捷克地区(如皮尔森啤酒厂)、丹麦的嘉士伯啤酒厂广泛使用;而Frohberg型酵母则在德国(如Weihenstephan啤酒厂)、丹麦的其他非嘉士伯啤酒厂、荷兰地区(如喜力啤酒厂,Oranjeboom啤酒厂)等地进行使用[8]。
这两种菌株在发酵性能上存在显著差异:Group Ⅰ型比Group Ⅱ型更加适合低温发酵,Group Ⅱ型在较高的温度下发酵更快。两种酵母在发酵末期的存活率相当,但Group Ⅱ型菌株产生呼吸缺陷型菌株(娇小突变)的几率更高。芬兰VTT技术中心通过对比在小试发酵条件下温度等对两种类型酵母发酵性能的影响,发现Group Ⅰ(Saaz)型酵母菌株对麦芽三糖的利用能力弱于Group Ⅱ(Frohberg)型,且Group Ⅰ型的酯类生成量较Group Ⅱ低[26]。
在2008年,DUNN和SHERLOCK等利用竞争性基因组杂交技术(CGH)分析了Lager酵母中S.cerevisiae来源、S.bayanus(现定义为S.eubayanus)来源基因的拷贝数及同源染色体的重组位点,结果也从基因组层面验证了Lager酵母的这两个分类(图2)[8]。这两种类型的Lager酵母在基因组层面上的主要区别是Group Ⅰ(Saaz型)型相对于Group Ⅱ(Frohberg型)型整体上缺失了很大一部分S.cerevisiae来源的基因组。大体上,Group Ⅰ型为三倍体,包含单倍体的Sc-亚基因组和双倍体的S.eubayanus亚基因组(Seb-亚基因组);而Group Ⅱ型为四倍体,包含双倍体的Sc-亚基因组和双倍体的Seb-亚基因组[27]。这种基因组水平差异的进化原理目前尚不明确,推测其原因可能是在低温发酵下的人为选择。
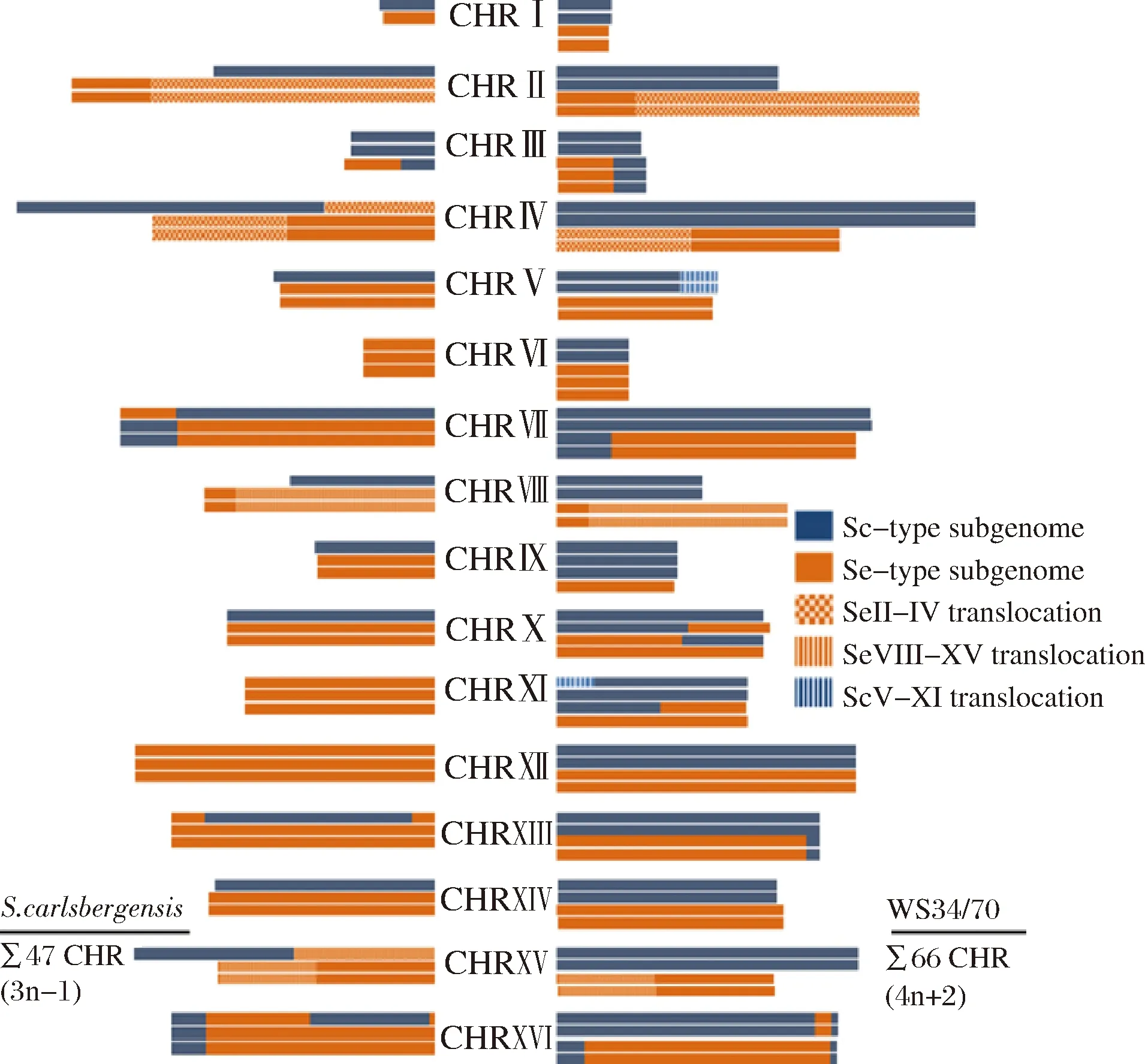
图2 不同类型啤酒酵母(Group Ⅰ和Group Ⅱ)的染色体结构和拷贝数[27]Fig.2 Map of the chromosome structures and copy numbersof different types of Lager yeast (Group Ⅰ and Group Ⅱ)
人们对不同类型Lager酵母明确的祖先和进化过程仍然有争议。目前主要存在3种假说,不同假说都拥有基因组学分析结果的支持[28]。第一种假说提出,Saaz型和Frohberg型的产生是起源于两个完全独立的杂交事件,所涉及的Ale酵母和野生S.eubayanus菌株亲本基因组均存在差异(图3-a),包括:转座子的分布、染色体缺失模式、染色体重排等[14, 18, 29-31]。第二种假说提出,Saaz型和Frohberg型由S.cerevisiae和S.eubayanus一次杂交后分别演化而来。这种假说的依据是在这两种不同类型的Lager酵母中发现了保守的染色体易位[32](图3-b)。也就是说在Saaz型和Frohberg型独自进化之前,在共同的亲本染色体中发生易位,并通过杂交保留在不同类型的酵母中。在后期的进化过程中,Frohberg型酵母中Sc-亚基因组中经由杂合性丢失而降低了同源染色体之间的差异,而Saaz型则丢失了一条S.cerevisiae来源的染色体。第三种假说是结合以上两种假说所提出,认为Saaz型和Frohberg型起源于单倍体S.cerevisiae和二倍体S.eubayanus之间至少一个共有的杂交事件。首先单倍体S.cerevisiae和二倍体S.eubayanus之间的单一杂交事件产生三倍体,进一步分化为Saaz型;而Frohberg型是由三倍体进一步与单倍体S.cerevisiae发生二次杂交所产生的[33](图3-c)。
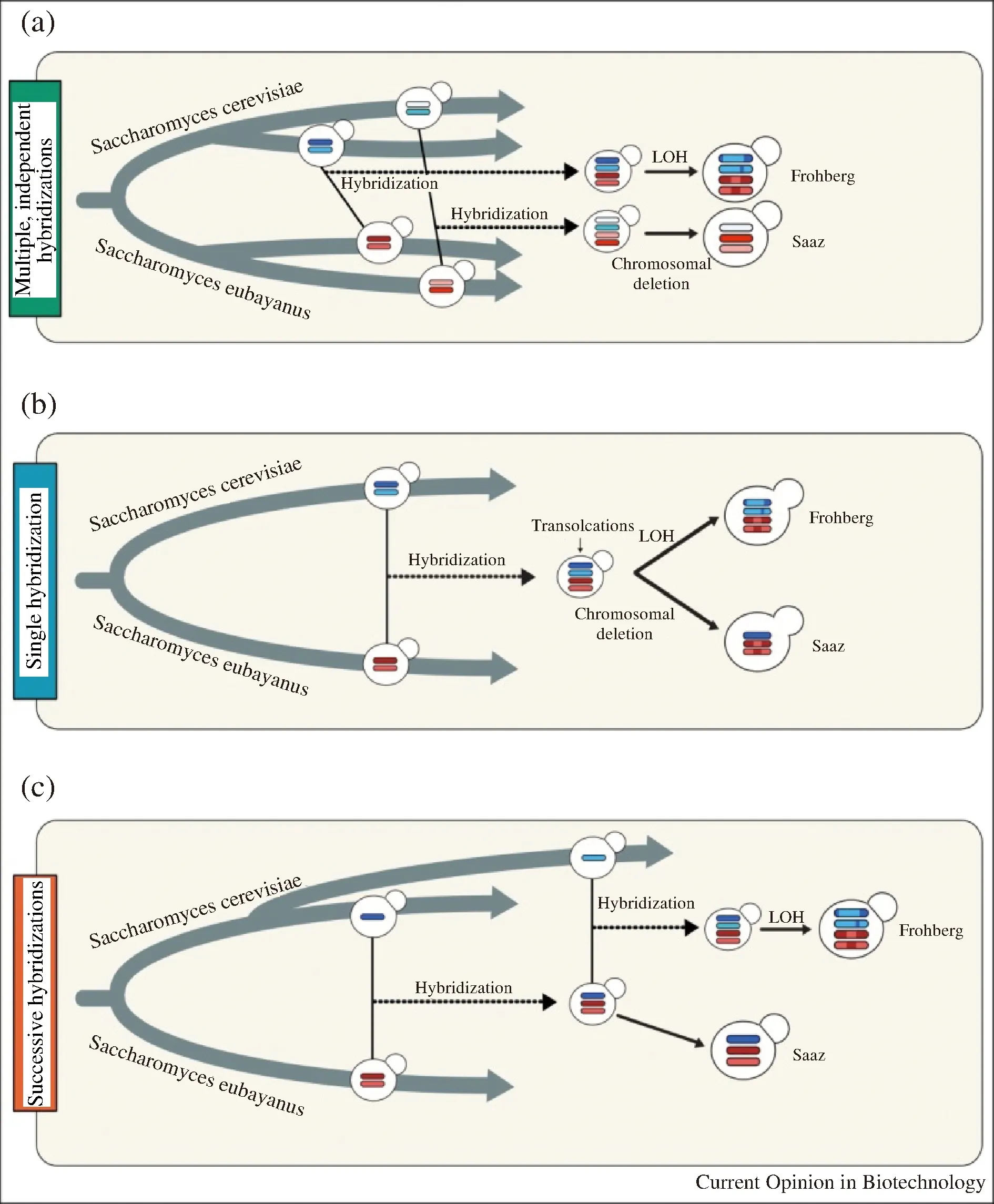
图3 Saaz型和Frohberg型起源的现有模型[28]Fig.3 Current models for the origin of Frohberg andSaaz lineages
4 Lager酵母的基因组结构
第一个完成全基因组测序的Lager酵母Weihenstephan 34/70 (WS 34/70),是Lager啤酒酿造中被广泛使用的Frohberg型菌株,由日本三得利在2009年完成[17]。其多倍体基因组大小为25 Mb,包含36条染色体,分别由酿酒酵母(S.cerevisiae)和贝氏酵母(S.bayanus)两个亚基因组构成,而后来贝氏酵母的亚基因组被确认是来自真贝氏酵母(S.eubayanus)[19]。从DNA含量的预测结果来看,该Lager酵母基因组为四倍体,但实际的染色体数量却较低,这也意味着在其染色体上存在基因的多拷贝现象[17]。其中,Sc-亚基因组与S.cerevisiae模式菌株S-288C的序列同源性高达99.2%,而S.bayanus来源的基因序列与当时唯一完成测序的S.bayanus菌株CBS 7001的序列相似性仅有92.7%。进一步通过分析基因组序列证实了Lager酵母中包含3种类型的染色体:类S.cerevisiae染色体、类S.bayanus染色体和杂合型染色体(来自于前两种的同源重组)[17]。同时还发现WS 34/70的线粒体基因组与S.bayanus菌株CBS 7001同源性很高,表明Lager酵母的线粒体DNA(mt DNA)应该是来源于非S.cerevisiae[17]。
在获得了WS 34/70的基因组序列后,近年来又有多株Lager酵母菌株陆续完成了全基因组测序,重点在于对不同菌株进行比较基因组学分析,并为进一步探索不同类型Lager酵母起源提供了基因组学基础[31-37]。Lager酵母基因组学的研究能够从生物进化的角度来阐明杂交菌株是如何通过基因的融合来实现对环境的适应。通过比较3株Lager酵母的染色体断点位置及序列发现,不同亲本来源的序列大多是在基因编码区发生染色体重组,而不是在基因组的重复元件(Repetitive elements)上。重组所产生的大量嵌合基因会减弱系统发生的信号,这才导致了在Lager酵母起源的研究上存在大量争议[35]。通过对Lager酵母染色体拷贝数的变化研究表明,Group Ⅰ型菌株的染色体组型较为一致,而Group Ⅱ型菌株之间则表现出较大的差异。此外,基因组序列基本一致的Lager酵母菌株在染色体拷贝数上也存在明显的变化[36]。
BAKER等指出,不同类型Lager酵母中Sc-亚基因组在与Seb-亚基因组发生杂交之前就已产生差异,在两个亲本的亚基因组中中性差异位点的数量存在差异。因此支持在Lager酵母进化过程中发生两次独立的杂交事件的假说[29]。与2009年对WS 34/70线粒体基因组分析结果不同的是,OKUNO等通过对Lager酵母mt DNA的系统发生学分析,揭示了Lager酵母的线粒体基因组可以从两个亲本的任意一方获得。此外还发现,Group Ⅰ型菌株和Group Ⅱ型菌株的mt DNA具有相同的来源(西藏)[37]。此外,研究发现在两种类型的Lager酵母基因组上至少存在3处相同的染色体间易位,认为Group Ⅰ型和Group Ⅱ型是由相同的祖先进化而来;但通过SNVs的分析却证明了不同类型Lager酵母基因组中含有不同的Sc-亚基因组,说明Group I型和Group Ⅱ型在其进化史上存在独立的进化起源[37]。因此认为两种类型Lager酵母至少经历了一次共同的杂交事件,进一步证明了第三种进化假说的可信性。
在Lager酵母基因组中,大多数的基因存在Sc-和Seb-两个同源拷贝。但研究发现,在某些基因中,一个来源的基因拷贝由于发生了移码突变或提前终止密码子会导致功能缺失[38-39]。由于有些基因是酵母生存的必需基因,因此推测这种突变是在两个亲本杂交之后产生的[17]。对杂交菌株来说,它平等地继承了亲本双方的基因及其特性,但随时间的推移及杂交菌株所受到的进化压力,一个亲本的基因组则会表现出其主导地位,从而改变菌株的性状[40-44]。这种性状的改变是由杂交菌株的基因组不稳定性所引起。许多研究也正是利用了Lager酵母的这种特性,通过人为引导进化(适应性进化)获得了性能优良的酵母菌株[45-50],包括高浓酿造Lager酵母、抗胁迫Lager酵母等。
5 结束语
在漫长的啤酒酿造历史中,人们无意识地对酵母进行了人工驯化及筛选,并酿造出美味的啤酒。与其他发酵产业不同,大规模啤酒酿造中所使用的Lager酵母菌株仍存在局限性。在生命科学领域早已步入基因组时代的今天,如何利用全新的基因组学知识指导优良酵母菌株的筛选也是当今的研究重点。Lager酵母菌株的单倍体分离及杂交技术现已十分成熟,实现Lager酵母菌株的定制化也不再是梦想。目前,国际许多研究机构都在进行大规模菌株杂交的方式筛选新的酵母菌株,并用来酿造啤酒。利用现代生物学知识和高通量实验技术,通过人工选择获得的酵母菌株与它们的祖先相比是否能够生产出口味更佳、更具竞争力的啤酒呢?还是让我们拭目以待!
