嗅觉学习对橘小实蝇寄主气味选择行为的影响
2019-05-23于金鑫方超周佳怡姚园园曾鑫年刘家莉
于金鑫,方超,周佳怡,姚园园,曾鑫年,刘家莉
(华南农业大学农学院/广东省昆虫行为调控工程技术研究中心,广州 510642)
学习是生物体获取信息的一个重要手段[1]。在对昆虫学习能力的研究中已发现,其可通过嗅觉[2]、视觉[3]等的学习去寻找和定位寄主[4]。昆虫的嗅觉系统具有高度专一和极其灵敏的特性,是昆虫识别和感知环境的重要感觉器,因此,有关昆虫嗅觉学习的研究也较深入且主要集中在联系性嗅觉学习上。西方蜜蜂(Apis mellifera)[5]可将花朵的气味与花蜜或花粉联系,从而提高搜寻蜜源植物的效率;德国小蠊(Blattella germanica)[6]和双斑蟋(Gryllus bimaculatus)[7]通过联系学习薄荷精油和蔗糖溶液而变得更趋向于薄荷精油;沙漠蝗(Schistocerca gregaria)[8]学习气味刺激和食物间的联系进而更偏好选择学习的气味;寄生蜂(Parasitoids)[9]也可通过过往的经历来提高对学习寄主的选择率。由上述研究可知,联系性嗅觉学习在昆虫的寄主选择中扮演着重要的角色。
经典性条件反射(classical conditioning)因其可精确化控制条件刺激(conditioned stimuli,CS)和非条件刺激(unconditioned stimuli,US)的作用时间和频率而被广泛应用于联系性嗅觉学习的研究中,昆虫的条件性伸喙反射则是其中的一种经典模式,多被应用于蜜蜂、果蝇、实蝇等具有长喙的昆虫研究中。在训练昆虫学习时,将条件刺激和非条件刺激重叠给予,若试虫成功进行联系性学习,则其对非条件刺激的伸喙反射行为也可由条件刺激激发,结果表现为对条件刺激的伸喙反射率升高[10]。然而,经典性条件反射中的伸喙反射行为并不能完全反应自由活动状态下昆虫的行为反应,因此,在学习后的记忆持续性测试中,观察昆虫对学习后气味的行为反应将更有利于反映学习对昆虫嗅觉偏好的影响。
橘小实蝇(Bactracera dorsalis)又名柑橘小实蝇、东方果实蝇、黄苍蝇、果蛆等,属双翅目(Diptera)、实蝇科(Tephritidae)、寡鬃实蝇亚科(Dacinae)、果实蝇属(Bactrocera),寄主遍及250多种果蔬,是一种重要的检疫性害虫[11]。我们在前期研究中采用伸喙反射的经典性条件训练法已发现橘小实蝇具有较强的联系性嗅觉学习能力,可将薄荷精油与蔗糖溶液联合,学习后对薄荷精油的伸喙反射率显著升高。甜橙是橘小实蝇的寄主之一,其对甜橙挥发物的学习能力及学习后的行为变化目前尚不十分清楚。本研究拟采用甜橙精油为条件刺激,运用伸喙反射的经典性条件训练法调查橘小实蝇对甜橙精油的学习能力,并进一步探索学习后橘小实蝇对甜橙精油的选择偏好性是否发生变化,以明确嗅觉学习对其寄主选择的影响。
1 材料与方法
1.1 材料
1.1.1 供试虫源
供试橘小实蝇为广东省昆虫行为调控工程技术研究中心2010年建立的室内种群,在光周期为14 h光照∶10 h黑暗、温度26~28℃、湿度60%~80%的实验室条件下饲养。将人工产卵器[12]内壁涂抹果汁,放入饲养笼内供成虫产卵,收集到的卵置于人工饲料(含香蕉、酵母、玉米粉、蔗糖、纤维、苯甲酸钠、盐酸、蒸馏水)中饲养,待幼虫老熟后,转到含水量为60%的细沙中待其化蛹。
羽化后的成虫放入30 cm×30 cm×30 cm的饲养笼内饲养,笼内放入人工饲料[m(酵母)∶m(蔗糖)=1∶3]和水供成虫取食,将羽化后2~3 d的雄虫从笼内隔离出单独饲养,选用16~19 d的未交配雄性成虫作为供试虫源。
试虫前处理:从实验种群中随机挑选20~30头雄成虫,置于30 cm×30 cm×30 cm的饲养笼内饥饿12 h(仅提供水,不提供饲料),以提高其对蔗糖溶液的嗜好性。试验当天,供试昆虫被固定于一塑料管(长3.5 cm,直径8 mm)中,并保证其头和前足可在管外自由活动[12]。试验30 min前使用蔗糖溶液测试,仅选用对蔗糖溶液有伸喙反应的个体继续试验,每头试虫仅用1次。据报道,橘小实蝇成虫每天活跃最高峰在8:00—11:00 a.m.[13-14],所以所有试验的时间固定为每日9:00—11:00 a.m.。在训练过程中死亡的个体被剔除(死亡率低于5%)。
1.1.2 供试药品
薄荷精油、甜橙精油(广州嘉媚乐天然用品连锁公司);蔗糖(分析纯,天津市大茂化学试剂厂);甲醇(一级色谱纯,天津四友精细化学品有限公司);正己烷(分析纯,上海润捷化学试剂有限公司)。
1.1.3 供试仪器
气泵(RESUNAIR-2000)、烧瓶、通风罩等。
1.1.4 气味刺激装置
由气泵产生的气流经活性炭除杂后再经过装有蒸馏水的烧瓶,使气流保持一定的湿度,气流速率由流量计调节,调节后的气流(2 L/min)通过装有气味源的烧瓶,以使气味混入气流中来提供气味刺激。刺激时气流末端对准试虫触角并保持1 cm的距离(图1)。此外,在训练位置后设有一个排气罩,不仅用于稳定气流且可及时将训练产生的气味排出室内[12]。

图1 嗅觉学习气味刺激装置Fig.1 Odor delivery system
将50 μL甜橙精油滴于“W”型滤纸(4 cm×4 cm)上,滤纸置于锡箔纸上,随后放置在通风橱内干燥10 min后备用。
1.2 试验方法
1.2.1 橘小实蝇的联系性嗅觉学习
1.2.1.1 经典性嗅觉条件训练法
本试验参考陈小燕等[12]所用的经典性嗅觉条件反射方法,选用甜橙精油作为条件刺激,因甜橙为橘小实蝇的寄主之一,用10%蔗糖溶液作为非条件刺激,联合对橘小实蝇进行奖赏性的嗅觉学习训练。训练时,首先将橘小实蝇放置于训练位置适应10 s,然后先给予气味刺激3 s,并记录其是否伸喙,接下来的3 s同时给予条件刺激和非条件刺激。给予非条件刺激时使用蘸取蔗糖水的牙签碰触橘小实蝇的喙,待橘小实蝇伸喙后使其取食3 s。每头试虫训练5次,训练的时间间隔为10 min,分别记录5次训练时试虫对甜橙精油的伸喙反射率。接受此训练法的实验组即为对称组。
1.2.1.2 非对称性训练对照组
在非对称训练组中,橘小实蝇共接受9次训练,即5次气味单独刺激和4次食物单独刺激(训练所用气味刺激为50 μL甜橙精油原液,食物刺激为10%蔗糖溶液),2种刺激以每5 min的间隔交替刺激试虫。在气味刺激训练时,给予气味6 s并记录气味释放前3 s时试虫对气味产生的伸喙反射;在食物刺激时,给予食物3 s。2种刺激间隔5 min是为了保证测定气味刺激诱导的伸喙反射间隔与实验组嗅觉训练的时间间隔一致,即为10 min。
1.2.1.3 气味刺激试验组与食物刺激试验组
试验中用于条件刺激和非条件刺激的嗅觉和味觉刺激也可能通过习惯性反应、敏感性反应或是其他非联系性学习来影响橘小实蝇的伸喙反射。为了明确非联系性学习的影响,设置了气味刺激试验组和食物刺激试验组。在气味刺激试验组中,仅用甜橙精油刺激橘小实蝇6 s,刺激前3 s时测试试虫对气味的伸喙反应,每头试虫刺激5次,每次间隔10 min。在食物刺激试验组中,喂食10%蔗糖溶液5次,喂食时间间隔设为10 min,最后一次喂食10 min后测试试虫对甜橙精油的伸喙反应。
1.2.1.4 记忆时长的测定
橘小实蝇接受5次联系性嗅觉训练,每次间隔10 min,训练方法同1.2.1.1。3组试虫分别在最后一次训练结束后的10 min、3 h和6 h后被测定对条件刺激甜橙精油的伸喙反应。
1.2.2 嗅觉学习对橘小实蝇选择偏好性的影响
1.2.2.1 橘小实蝇气味选择性测定
本试验选用Y型嗅觉仪(图2)测定橘小实蝇对气味的选择偏好性。Y型管主臂长20 cm,两臂长20 cm,两臂夹角为60°。在Y型管前端10 cm处放置光源(76 lx),并罩白纸以减弱光的强度,利用趋光性来提高橘小实蝇的出管选择率。由气泵产生的气流经流量计调节流量(500 mL/min)后,再经活性炭除杂,蒸馏水加湿后进入放有气味源的烧瓶,最后载有气味的气流进入Y型管来测试橘小实蝇对两臂气味的选择性。试验时,在Y型管直臂接入一定量的试虫数,10 min后观察进入两臂2/3处的试虫数。试虫仅测试一次,每次测试后更换滤纸,调换气味源方向,更换Y型管。测试后的Y型管用甲醇清洁干净后备用。本试验选用的气味源A为甜橙精油,气味源B为薄荷精油,测试时,吸取各精油50 μL滴于4 cm×4 cm的滤纸上,晾置10 min后置于气味源烧瓶中。
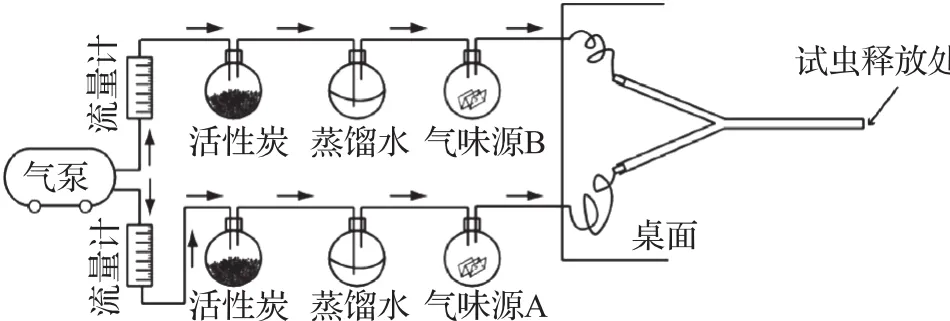
图2 Y型嗅觉仪Fig.2 Y-type olfactometer
1.2.2.2 嗅觉学习训练前后橘小实蝇的气味选择偏好性测定
将未经训练的试虫固定于塑料管内,固定时间与5次训练所需时间一致,固定时间结束后,试虫从管内被释放至一干净养虫笼内,10 min后用于选择性测试。测试总虫数不低于30头,测定后计算选择率。试虫在接受联系性训练(方法见1.2.1.1)、非对称训练(方法见1.2.1.2)、气味单独刺激和食物单独刺激(方法见1.2.1.3)后由固定管中释放出来,置于干净养虫笼内10 min后用于测试,测试所用方法见1.2.2.1。
1.2.3 嗅觉学习对橘小实蝇化学趋风性(chemoanemotaxis)的影响
1.2.3.1 橘小实蝇化学趋风性测定
本试验选用自制微型风洞(图3),长50 cm,直径3 cm,由气泵产生的气流经流量计调节流量(500 mL/min)后,再经活性炭除杂,蒸馏水加湿后进入放有气味源的烧瓶(气味源为滴于4 cm×4 cm的滤纸上的甜橙精油)中,最后载有气味的气流进入风洞以测试橘小实蝇的化学趋风性。每次测试1头,每头试虫仅测试1次,每次测试后更换滤纸,并用甲醇清洁风洞,每次测试时间5 min,统计记录到达风洞气味源的试虫数。

图3 微型风洞装置Fig.3 Mini wind tunnel equipment
1.2.3.2 训练前后橘小实蝇对甜橙精油化学趋风性的影响
将未经训练的橘小实蝇固定于试管内,固定时间与5次训练所需时间一致,固定时间结束后,试虫从管内被释放至一干净养虫笼内,10 min后测试其对甜橙精油的趋性,测试总虫数不低于30头,测定后计算趋性率。试虫在接受联系性训练、非对称训练、气味单独刺激和食物单独刺激后由固定管中释放出来,10 min后测试其对甜橙精油的趋性,测试总虫数不低于30头,测定后计算趋性率。
1.2.4 数据记录与统计分析
学习后条件反射率(conditioned response):统计每次训练中进行伸喙反射的橘小实蝇个体占比(发生伸喙反射的个体占总测试虫数的百分率)。用科克伦试验(Q)[Cochran's test(Q)]分析5次连续训练对伸喙反射的影响。用列联表G检验分析某一训练次数下不同训练组间伸喙反射率的差异。多组间伸喙反射率差异采用χ2多重比较,显著性经Brunden矫正,矫正α´=α/2(k-1),k为样本数。
学习表现力(learning performance):统计每头试虫在5次训练中总的伸喙次数(0~5次)。用Kruskal-Wallis(KW)和 Scheffé事后多重比较(post hoc tests)分析多组间橘小实蝇的学习表现力,用Mann-WhitneyU检验比较2组间橘小实蝇的学习表现力[15]。
Y型管选择率:对条件气味甜橙精油的选择率=(进入甜橙精油气味管臂试虫数/做出选择的试虫数)×100%。
化学趋风性:对甜橙精油的趋向率=(到达气味源的试虫数/供试虫数)×100%。
各试验组间选择率和趋向率的比较采用χ2多重比较法,统计分析所使用软件为SPSS 9.0。
2 结果与分析
2.1 甜橙精油作为条件刺激的嗅觉学习
2.1.1 对称性与非对称性训练下的嗅觉学习
橘小实蝇对甜橙气味的伸喙反射率随5次连续训练显著增加(Cochran试验,Q=299.28,df=4,P<0.001;图4),由第1次的8.1%增加至第5次的75.7%。在所有训练试虫中,仅有11.9%的试虫在所有5次训练中均未伸喙。当以甜橙精油和蔗糖溶液交替刺激试虫各5次时,伸喙反射率随训练次数增加无显著变化(Q=1.62,df=4,P=0.81)。对称组与非对称组在第2次训练中所测得的伸喙反射率已存在显著差异(G=33.54,df=1,P<0.01)。在随后的训练次数中仍保持着差异(第3次训练,G=62;第4次训练,G=68.4;第5次训练,G=63.6;所有df=1,P<0.01)。对称组与非对称组间的学习表现力也存在显著差异(Mann-Whitney试验,U=2 245,P<0.001)。
2.1.2 气味或食物单独刺激对嗅觉学习的影响
在气味刺激试验组(n=57)中,橘小实蝇在第1次训练中的伸喙反射率即自发伸喙反射率为6.3%。经随后4次刺激后,伸喙反射率无明显增加(Q=1,df=4,P=0.91),仅有1头伸喙4次,4头伸喙1次,伸喙率保持为1.5%~3.0%。
在食物刺激试验组(n=60)中,取食过蔗糖溶液5次后的橘小实蝇伸喙反射率为11.6%,并与自发伸喙反射率(4.3%)进行比较表明,两者间无显著差异(χ2=2.47,df=1,P=0.12)。
2.1.3 橘小实蝇记忆时长测定结果
分别于训练后的10 min、3 h和6 h测定橘小实蝇对甜橙精油的伸喙反射率。经不同时间后,橘小实蝇的伸喙反射率仍显著高于训练前(10min,G=89.9;3 h,G=61.55;6 h,G=47.69;df=1,P<0.01;图5)。对比不同时间段的伸喙反射率发现,随训练后时间的推移,伸喙反射率逐渐降低(G=9.63,df=2,P<0.01)。6 h后的伸喙反射率显著低于10 min后的伸喙反射率(10 min与6 h:χ2=9.54,df=1,α´=0.012 5,P<0.05),而10 min和3 h及3 h和6 h的伸喙反射率间无差异(10 min与3 h,χ2=1.73;3 h与6 h,χ2=2.94;df=1,α´=0.012 5,P>0.05)。
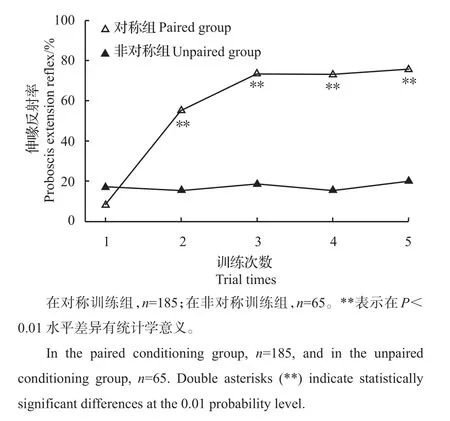
图4 橘小实蝇对甜橙精油的伸喙反射率Fig.4 Proboscis extension reflex of B.dorsalis to sweet orange essential oil
2.2 橘小实蝇嗅觉学习对甜橙精油选择性的影响
未经训练的试虫经Y型管测试,其对甜橙精油的选择率为35.3%,经5次对称训练后其对甜橙精油的选择率为63.7%,极显著高于训练前的选择率(χ2=6.9,P<0.01;图6)。橘小实蝇经5次非对称训练后其对甜橙精油的选择率为13.7%,显著低于经5次对称训练后其对甜橙精油的选择率(χ2=25.1,P<0.01;图6)。
橘小实蝇经5次甜橙精油气味单独刺激后,其对甜橙精油的选择率为16.4%,经5次蔗糖溶液单独喂食后,其对甜橙精油的选择率为24.5%,2组对甜橙精油的选择率均显著低于对称组训练后对甜橙精油的选择率(气味刺激组与对称组,χ2=17.3,df=1,α′=0.012 5,P<0.05;食物刺激组与对称组,χ2=11.8,df=1,α′=0.012 5,P<0.05)。
2.3 橘小实蝇嗅觉学习对甜橙精油趋性的影响
将未经训练的橘小实蝇放入微型风洞,仅有8.3%的试虫表现为正相趋风性(positive anemotaxis),即趋向于气味源甜橙精油。经5次对称训练后的橘小实蝇放入微型风洞后,有52.5%的橘小实蝇趋向于气味源,趋向率显著高于训练前的橘小实蝇(χ2=14.47,P<0.01)。经5次非对称训练后,仅有10%的橘小实蝇趋向于气味源,趋向率显著低于对称训练后的橘小实蝇(χ2=16.8,P<0.01)。橘小实蝇经甜橙精油单独刺激5次后,其对甜橙精油的趋向率为24.3%,显著低于对称训练后的橘小实蝇(χ2=6.4,P<0.05);而喂食蔗糖溶液5次后,其对甜橙精油的趋向率为12.8%,也显著低于对称训练后的橘小实蝇(χ2=14.1,P<0.01)(图7)。
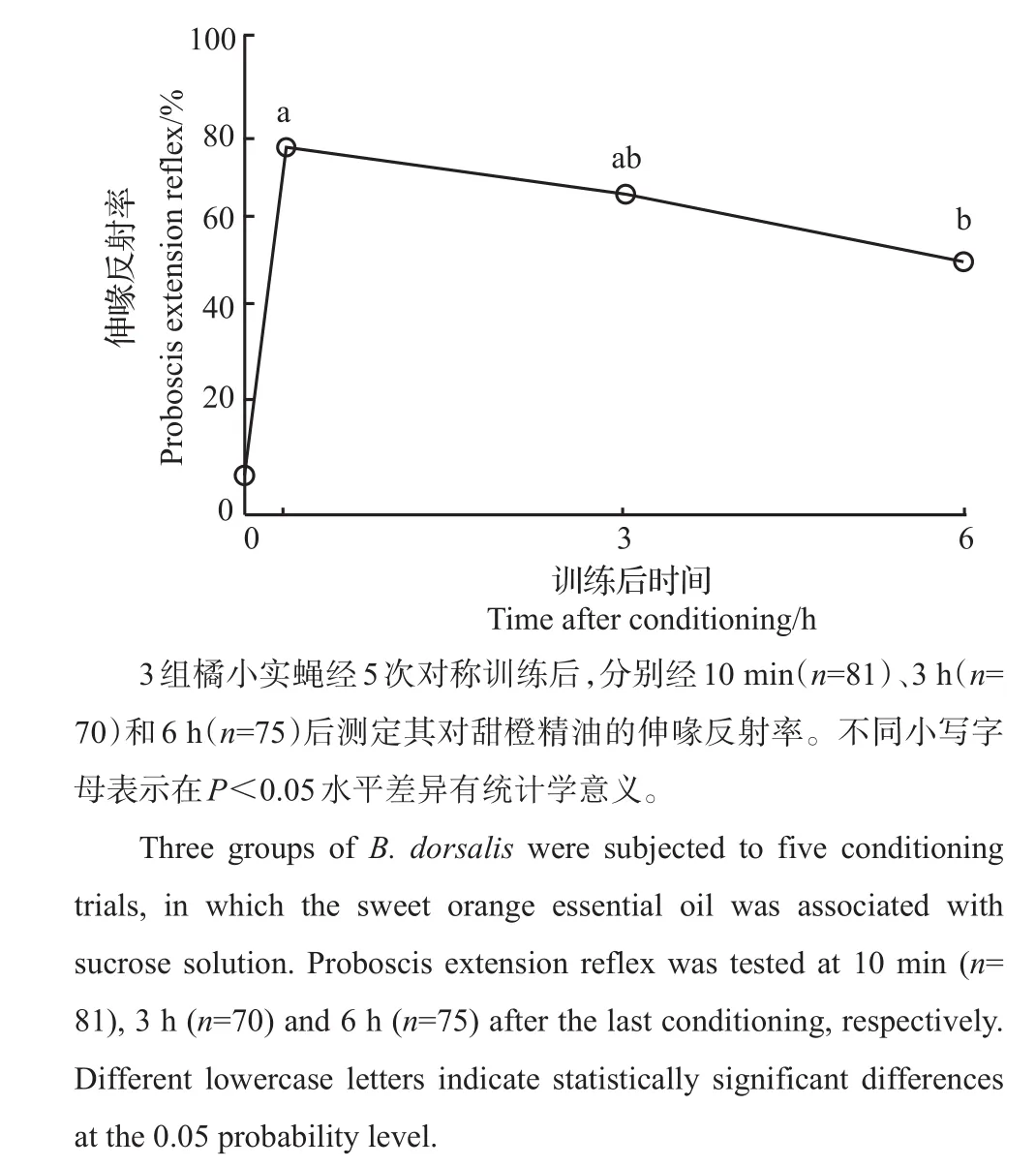
图5 橘小实蝇对甜橙精油的记忆时长Fig.5 Retention of olfactory memory of B.dorsalis to sweet orange essential oil

图6 橘小实蝇Y型管选择率测定Fig.6 Y-tube selection rate test in B.dorsalis

图7 橘小实蝇训练前后的化学趋风性Fig.7 Chemo-anemotaxis of B.dorsalis before and after training
3 讨论
3.1 橘小实蝇的奖赏性嗅觉学习
通过经典性嗅觉条件训练法可成功训练被固定于试管中的橘小实蝇联合甜橙精油与蔗糖溶液,学习后的橘小实蝇对甜橙气味的伸喙反射率显著提高,这一结果表明甜橙气味经学习后已成为一种信号,这一信号预示了食物的存在。通过对比对称训练组和非对称训练组可知,条件刺激的气味和非条件刺激的食物的短暂联合是学习形成的必要条件,即两者同时刺激时,时间上的交错是引起伸喙反射率增加的原因,而若时间上无交叉则不能引起伸喙反射率的增加。这种因条件刺激和非条件刺激的对称性联合而产生的学习行为在很多昆虫中均有证明,如西方蜜蜂[5]、烟芽夜蛾(Heliothis virescens)[16]、沙漠蝗[8]等。
在对称训练时,训练的结果极有可能受条件刺激或非条件刺激产生的习惯性反应或敏感性反应的影响,因此,本文也测定了气味单独刺激及食物单独喂食后是否对橘小实蝇的伸喙反射率产生影响。结果表明,气味或食物的单独多次刺激并不使橘小实蝇对训练气味的伸喙反射率产生任何影响。但在我们的前期研究中曾发现橘小实蝇取食过蔗糖溶液后,其对薄荷精油的伸喙反射率有轻微升高,这种升高有可能由敏感性反应所引起[12]。因取食蔗糖而引起的敏感性反应在西方蜜蜂中亦有发现:西方蜜蜂在取食一次蔗糖溶液后,其对气味的伸喙反射率可迅速增加2倍,并在2 min内降回至最初的水平[1]。
3.2 奖赏性嗅觉学习对橘小实蝇气味偏好性的影响
在经典性条件训练法中获得对条件刺激的条件反射并不能明确反应嗅觉学习对昆虫选择行为的影响,因此,嗅觉学习训练后选择合适的行为测试可研究嗅觉学习对昆虫行为的影响。在对黑腹果蝇(Drosophila melanogaster)[17]、昆士兰果实蝇(Bactrocera tryoni)[18]、西 方 蜜 蜂[19]、美 国 大 蠊(Periplaneta Americana)[20]、德国小蠊[6](Blattella germanica)、双斑蟋(Gryllus bimaculatus)[7]及沙漠蝗[8]的研究中均发现嗅觉学习可引起昆虫随后对气味偏好性的改变。在本试验中也发现嗅觉学习可引起橘小实蝇气味偏好性改变,经奖赏性训练后,橘小实蝇对学习过的甜橙精油的选择率相较于训练前明显增加,且通过与非对称组及气味刺激和食物刺激组比较可知,此选择率的增加是联系性的奖赏性嗅觉训练所引起的,这一结果表明橘小实蝇可能更倾向于选择曾经有过取食经历的寄主,同时,研究结果也表明橘小实蝇可将学习后对甜橙精油的条件反应转化为选择行为。
3.3 橘小实蝇对学习气味的定位机制
在大多爬行或飞行昆虫中,橘小实蝇对气味源的定位机制已被证实为由气味改变的趋风性,即化学趋风性[21-24]。VET等[25]和PAPAJ等[9]曾通过使用昆虫运动轨迹记录仪研究了一种寄生蜂Leptopilina heteroma的产卵经历对其搜寻定位行为的影响,发现在某一寄主产卵后的寄生蜂对该寄主的趋性增强,这一趋性表现为化学趋风性的增强,即风上移动时间缩短,移动轨迹更趋于一致。嗅觉学习对趋性强度的影响还表现在趋化性上,在研究秀丽隐杆线虫(Caenorhabditis elegans)[26]的学习中发现,厌恶性学习可降低线虫的趋化性,从而影响其对食物的选择。本研究的微型风洞结果显示,经联合对称训练后向风上方向移动趋向于气味源的橘小实蝇显著增多,表现为正向趋风性的增强。而这一趋性的增强仅由对称性训练引起,而非单一刺激和非对称训练,表明了联合性学习是引起橘小实蝇定位气味源的必要条件。在复杂的生境中,为了快速高效搜寻到取食寄主,橘小实蝇可能通过学习以记忆食物资源的气味,在今后的搜寻中可对记忆的气味快速反应并趋向定位到气味源。而学习后橘小实蝇是否因嗅觉反应变化从而导致行为发生变化尚不明确,这也将是未来研究的一部分。
4 结论
由本文的实验结果可知,橘小实蝇具有较强的联系性嗅觉学习能力,经与蔗糖溶液联合训练可学习甜橙精油的气味,学习后其对甜橙精油的伸喙反射率明显升高,且这种升高的必要条件为对称性训练而非非对称训练引起的敏感性反应和习惯性反应。经对称训练后,其对甜橙精油的伸喙反应可转移为行为上的选择偏好性和趋向性,表现为在选择性试验中,与训练前相比,试虫更倾向于选择学习过的甜橙精油;在趋性试验中,与训练前相比,试虫对学习过的甜橙精油的趋性增加。
