肉制品加工中脂肪对甲型副伤寒沙门氏菌rpoE的应激特性
2019-05-18王亚飞李苗云崔文明赵改名柳艳霞高晓平
王亚飞 李苗云 崔文明 赵改名 柳艳霞 高晓平
(1 河南农业大学食品科学技术学院 河南省肉制品加工与质量安全控制重点实验室 郑州450002 2 中检集团中原农食产品检测(河南)有限公司 郑州450002)
甲型副伤寒沙门氏菌是一种无荚膜的革兰氏阴性杆菌,能够引起伤寒症和急性消化道传染病,主要通过受污染的食物和水源暴发流行。据统计,伤寒症导致每年有217 000 死亡数,其中,中南亚和东南亚是主要的疾病暴发地。已报道的几乎80%伤寒病例来自于8 个南亚国家(孟加拉国、中国、印度、印度尼西亚、老挝、尼泊尔、巴基斯坦和越南),发病以儿童、青少年较多[1]。目前,伤寒肠热病仍是全球的重大公共卫生问题之一,尤其在亚洲。食品的杀菌方法很多,包括物理、化学和生物等。热处理是商业应用的标准方法,它是食品工业最经济、最简便和使用最广泛的杀菌方法[2]。为了减少高温对肉制品风味、营养成分的破坏,许多肉制品加工都采用巴氏杀菌的方法来减少细菌病原体[3-4]。
σE调节因子由rpoE 基因编码,是肠杆菌科中重要的σ 因子[5],广泛存在于各种原核细胞中,尤其在响应不同环境应激时发挥重要作用[6]。rpoE 的缺失会导致其对温度胁迫能力大为降低[7-10],且某些细菌在高渗和缺乏碳源的饥饿环境下rpoE 都发挥了重要作用[11-13]。rpoE 也能提高细菌抗氧化的能力[14-15]。肉类产品热加工过程中复杂介质的物理和化学成分对微生物的杀伤力有影响,其中脂肪对菌株的耐热性起重要作用。先前有报道指出脂肪导热性差,水分活度低会导致微生物更高的耐热性[16-18]。基于rpoE 对不同环境的应激能力,脂肪是否会通过调节rpoE 来调高菌株的耐热性尚未有文章报道。
本研究旨在探究肉制品加工中脂肪对甲型副伤寒沙门氏菌rpoE 的应激特性。首先通过PCR检验△rpoE 缺陷株缺陷并稳定表达,探究肉制品加工条件下脂肪对缺陷株和野生株的致死情况,从而进一步利用实时荧光定量PCR 确定脂肪对rpoE 表达水平的影响。
1 材料与方法
1.1 材料与仪器
1.1.1 菌株 试验所用甲型副伤寒沙门氏菌从肉中分离获得,均经过国标法生化鉴定、Vitek 测定以及测序鉴定。缺陷株△rpoE 是前期试验利用此株甲型副伤寒通过Red 同源重组构建获得。
1.1.2 试剂 营养肉汤(Nutrient broth,NB)、木糖赖氨酸脱氧胆盐 (Xylose lysine deoxycholate salt agar,XLD)琼脂,青岛高科园海博生物技术有限公司;氯化钠(分析纯),天津市瑞金特化学有限公司;EASYspin plus 细菌RNA 快速提取试剂盒,北京艾德莱生物科技有限公司;Ezup 柱式基因组DNA 抽提试剂盒 (细菌)、4S Red Plus 核酸染色剂、琼脂糖、第一链cDNA 合成试剂盒(AMV First Strand cDNA Synthesis Kit)(SK2445),上海生物工程有限公司;引物由上海生物工程有限公司合成。
1.1.3 仪器与设备 HVE-50 高压蒸汽灭菌锅,日本HIRAYAMA 公司;SPX-1505H-Ⅱ生化培养箱,上海新苗医疗器械公司;SW-CJ-2F 超级工作台,苏州安泰空气计数有限公司;THZ-C 台式恒温振荡器,太仓市华美生化仪器厂;EasyMix 均质机,法国AES 公司;THZ-82 水浴锅,金坛市杰瑞尔电器有限公司;SW-CJ-1D 洁净工作台,江苏苏洁净化设备厂;HC-2518R 高速冷冻离心机,安徽中科中佳仪器有限公司;DYY-6C 型稳压稳流电泳仪,北京六一;H6-1 微型电泳槽,上海精益有机玻璃制品仪器厂;凝胶成像系统,上海复日科技有限公司;TU-1901 紫外分光光度计,北京普析通用仪器有限公司;PCR 反应扩增仪,德国biometra 公司;StepOne 型荧光定量PCR 仪,美国ABI 公司。
1.2 试验方法
1.2.1 缺陷株△rpoE 鉴定 将野生株和缺陷株从冰箱里取出,复苏,分别接种于营养肉汤(NB)中,置于37 ℃过夜培养,检查生长情况。采用Ezup 柱式基因组DNA 抽提试剂盒(细菌)提取野生株和缺陷株的基因组DNA。根据NCBI 沙门氏菌rpoE基因序列用Primer Premier 5.0 软件设计引物F:GAATCGCGGATCAGGT,R:TGCGGCTTATGGAG TG,缺陷株和野生株在繁殖10 代后进行PCR 鉴定,所扩增片段大小分别为1 261 bp,795 bp,对比野生株和缺陷株PCR 产物,确定缺陷株稳定性。
1.2.2 人工污染介质 挑取野生株和缺陷株甲型副伤寒沙门氏菌单菌落,分别接种于100 mL NB液体培养基中37 ℃摇床培养16 h,达到稳定期。分别用纯瘦肉灌肠和肥瘦肉1 ∶1 灌肠作为介质,采用注射法进行人工污染,人工污染的菌液浓度为108CFU/mL,样品搅碎称取25 g 注射2.5 mL 菌液[19],放入真空袋中压成片状,用真空包装机抽真空,封口,37 ℃培养16 h。
1.2.3 甲型副伤寒沙门氏菌野生株和缺陷株在不同介质下的热处理 水浴锅温度设为55,63,72℃,加热时间从0~180 min 不等。加热时,将培养后的肉样完全浸没在热水中,按照预先设定的时间间隔,定时取出样品进行测定。热处理后的样品应立即放入预先准备好的冰水中阻止其热失活过程。从设定的温度冷却至室温的时间小于10 s。采用稀释涂布XLD 培养基进行活菌计数,37 ℃条件下培养18~24 h。
1.2.4 甲型副伤寒沙门氏菌野生株在不同介质下的热处理rpoE 的表达水平 培养后的肉样进行热处理控制水浴温度55 ℃,热处理30 min,处理后立刻放在冰水中冷却,不进行热处理的肉样为空白对照。取25 g 样品置于无菌均质袋中,加入225 mL 生理盐水,拍打取上清液4 ℃,2 500 g 离心10 min,收集菌体用PBS 缓冲液冲洗2~3次[20]。
采用Real-time PCR 法检测野生株在不同介质中热处理后的残存菌体样本中rpoE 基因相对含量。操作过程:细菌总RNA 提取根据操作说明;利用RNA 逆转录酶合成第一链cDNA;利用cDNA进行PCR 反应合成目的基因和内参基因16S,采用荧光定量PCR 仪对整个过程进行实时定量监控,并对数据进行分析处理。引物序列为rpoE-F:5’CCAGCATAAAGTGGCGAGTC3’,rpoE-R:5’TGACCGCAATACGATACAACC3’;16S-F:5’CC TACGGGAGGCAGCAG3’,16S-R:5’ATTACCGCG GCTGCTGG3’。配备反应体系如表1所示。按照下列条件进行反应:95 ℃,3 min→95 ℃,7 s→57 ℃,10 s→72 ℃,15 s(45 个循环),测60~99 ℃溶解曲线。

表1 PCR 反应体系Table1 PCR systems
1.2.5 数据处理 数据均采用SPSS17.0 进行数据处理与统计分析,所有数据均为3 个平行样品测定3 次。热失活曲线采用Origin 9.0 软件绘制。D 值是指在某一温度下,活菌数死亡90%所需要的时间,及细菌残存曲线经一个对数周期所需要的时间。D 值是细菌死亡率的倒数,D 值越大死亡速度越慢,该菌的耐热性越强。用Linear 模型拟合热致死曲线式中,x 表示加热时间,y 表示细菌活细胞的对数值,可得出该函数的斜率的负倒数为所求的D 值,即D=-1/k。Real-time PCR 结果采用log22-△△Ct方法和SPSS17.0 进行差异比较分析。
2 结果与分析
2.1 缺陷株PCR 鉴定
繁殖10 代后,提取缺陷株和野生株的DNA,然后进行PCR 扩增,可看出野生株可以扩增出约795 bp 基因片段,而突变株扩增出1 261 bp 的基因片段(red 重组敲除工作中未进行抗性基因无痕敲除,因此缺陷株的片段比野生株长),不过从结果看出缺陷株无法正常表达rpoE(图1),证实了缺陷株△rpoE 的基因稳定性,且稳定性极好。
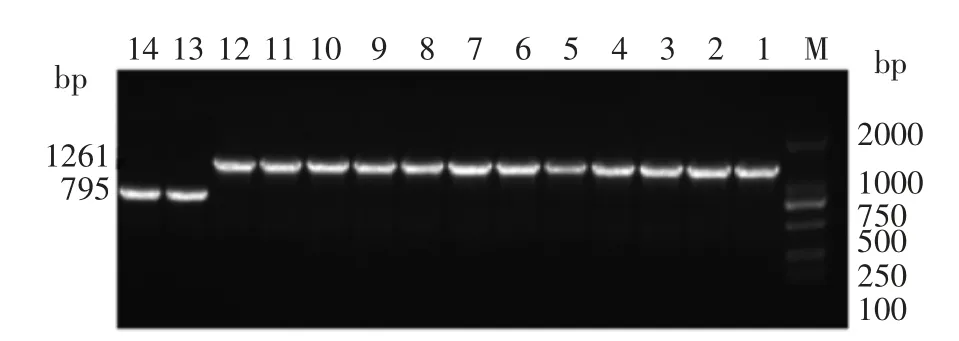
图1 繁殖10 代后野生株和缺陷株凝胶电泳鉴定Fig.1 Verification of the wild and mutant strain after 10 generations of reproduction
2.2 甲型副伤寒沙门氏菌野生株和缺陷株在不同介质下的热失活效果
通过模拟肉制品加工过程中近似巴氏杀菌的条件,本试验选择55,63,72 ℃对纯瘦肉灌肠和肥瘦肉1∶1 灌肠进行热处理。研究结果如图2所示,甲型副伤寒沙门氏菌不同介质中缺陷株和野生株的热失活情况有所不同,肥瘦1∶1 介质缺陷株和野生株的耐热能力强于纯瘦介质,并且随着样品热处理时间的延长,脂肪对菌株的耐热性影响越明显,直至全部死亡。
2.3 甲型副伤寒沙门氏菌野生株和缺陷株在不同介质下的耐热性分析
细菌的耐热性常用D 值表示,如表2所示,55 ℃下,纯瘦肉中缺陷株和野生株的D 值分别为14.0980,10.8933 min,肥瘦1 ∶1 介质中分别为24.9585,13.5506 min。63 ℃下,纯瘦肉中缺陷株和野生株的D 值分别为1.3078,0.3147 min,肥瘦1∶1介质中分别为2.0284,0.4591 min。72 ℃下,纯瘦肉中缺陷株和野生株的D 值分别为0.1740,0.0667 min,肥瘦1∶1 介质中分别为0.3019,0.0856 min。


图2 不同介质下野生株和缺陷株的热失活曲线Fig.2 The survival curves of wild strains and mutant strains indifferent medium

表2 不同介质下缺陷株和野生株的D 值Table2 Heat resistance (expression as D-values in min) for wild strains and mutant strains in different medium at 55,63,72 ℃
横向比较,相同条件下缺陷株和野生株D 值之间存在显著性差异(P<0.05),缺陷株的D 值都明显低于野生株,缺陷株相对于野生株对高温更加敏感。这与先前报道rpoE 耐热功能相一致[7-10],rpoE 基因对甲型副伤寒沙门氏菌的耐热能力也有重要作用。纵向比较,相同温度下缺陷株和野生株分别在肥瘦1∶1 和纯瘦肉介质之间的D 值都存在显著性差异(P<0.05),不管rpoE 基因是否存在,菌株在脂肪含量高介质下的耐热性都高于纯瘦介质,进一步说明脂肪确实对菌株的耐热性起作用,然而脂肪是否会影响rpoE 来调节还不能确定,因此接下来对rpoE 表达水平进行研究。
2.4 甲型副伤寒沙门氏菌野生株在不同介质下热处理rpoE 的表达水平比较
根据2.3 节的D 值结果,选取模拟肉制品加工过程中近似巴氏杀菌的条件为55 ℃,30 min。按照1.2.4 步骤,对rpoE 基因进行实时荧光定量PCR 反应,对比分析肉制品杀菌过程中不同介质下野生株rpoE 基因的表达水平,其结果如图3所示。野生株在介质肥瘦1∶1 和介质纯瘦灌肠中热处理的表达量分别约为对应空白的3.53 倍、3.29倍,结果进一步证明了热激条件下甲型副伤寒沙门氏菌rpoE 的表达上调来提高菌株的生存能力。然而,两种介质条件下的rpoE 表达不存在显著差异(P>0.05),脂肪并不会影响rpoE 表达水平,猜测脂肪对甲型副伤寒沙门氏菌耐热性的影响可能由于物理或其它基因调节产生一个复杂的生物变化过程。

图3 rpoE 基因在不同介质加热处理下的表达水平Fig.3 The relative expression level of rpoE in different medium
3 结论
Red 重组技术适用于甲型副伤寒沙门氏菌基因敲除,稳定性高。肉制品巴氏杀菌近似条件下,rpoE 的表达水平都达到3 倍以上,可提高甲型副伤寒沙门氏菌的耐热性。脂肪对菌株的耐热性起重要作用,并不会通过调节rpoE 的表达来提高耐热性。
