长爪沙鼠CST3序列同源性分析及蛋白体外表达*
2019-05-16杜小燕李长龙霍学云吕建祎陈振文
王 妍 郭 萌 杜小燕 李长龙 路 静 霍学云 吕建祎 刘 欣 陈振文
(首都医科大学基础医学院,北京 100069)
长爪沙鼠(Merionesunguiculatus)又名蒙古沙鼠(Mongoliangerbil),为啮齿类动物家族成员,是源自我国的实验动物资源[1]。由于其独特的解剖结构、生理学和行为学特征,长爪沙鼠在许多研究领域具有特殊的应用价值,如脑神经病研究、寄生虫病研究、微生物学研究、听觉和视觉研究、内分泌研究、代谢研究、肿瘤及其他疾病研究[1-2]。可以说,长爪沙鼠被称为“多功能实验动物”。随着长爪沙鼠线粒体和全基因组测序[2]的完成,以及癫痫易患模型[3]、自发糖尿病模型[4]、脑缺血高发群体[5]等长爪沙鼠动物模型群体的建立,长爪沙鼠逐渐成为相关研究领域的一种“热门”实验动物。近年来,在长爪沙鼠近交系和封闭群培育过程中,我们在证实长爪沙鼠脑底动脉Willis环(circle of Willis,CoW)变异缺失具有遗传性的基础上,通过抑制消减杂交(Suppression Subtractive Hybridization,SSH)方法,发现半胱氨酸蛋白酶抑制剂C (Cystatin C,CST3)的表达变化与长爪沙鼠脑血管畸形相关[6]。为了进一步研究长爪沙鼠CST3的功能和参与血管发育的机制,我们比对了CST3序列同源性,构建了长爪沙鼠CST3蛋白表达载体,对其进行原核表达和鉴定,为制备长爪沙鼠CST3抗体和进行后续的功能研究奠定基础。
1 材料与方法
1.1 实验动物
选取来自首都医科大学实验动物部的封闭群长爪沙鼠1只,雄性,16周龄,饲养于普通环境中(SYXK (京) 2013-0005)。将长爪沙鼠过量麻醉安乐死后,于冰上立即解剖,获取长爪沙鼠新鲜脑组织。组织采集后保存于液氮中。
1.2 主要实验材料
pET28a质粒为实验室保存;克隆载体pMD19-T(Takara,日本);FastQuant cDNA 第一链合成试剂盒、胶回收试剂盒、无内毒素大提质粒试剂盒、E.coliBL21感受态细胞、E.coliDH5a感受态细胞均购自天根公司;TRIzol Reagent(Invitrogen,美国);NcoI和XhoI限制性内切酶(NEB,美国);IPTG、硫酸卡钠霉素、氨苄青霉素均购自索莱宝公司。
1.3 方法
1.3.1总RNA提取及cDNA制备: 取约0.1 g脑组织,加入到1 mL TRIzol试剂中进行组织匀浆,混匀后室温消化10 min。加入200 μL三氯甲烷,颠倒混匀后冰上放置15 min,13 500 r/min 4 ℃离心15 min。将上清液转移至新的离心管中,加入0.5 mL异丙醇,颠倒混匀后冰上放置15 min,13 500 r/min 4 ℃离心15 min,弃上清。加入无核酶冰75%乙醇洗涤沉淀2次,13 500 r/min 4 ℃离心5 min后弃上清。自然风干后无核酶水溶解,获得长爪沙鼠脑组织总RNA。使用Nanodrop 2000c (Thermo scientific, USA) 测量总RNA浓度和纯度,并根据FastQuant cDNA 第一链合成试剂盒说明书要求进行反转录获得cDNA。
1.3.2Cst3基因蛋白质编码区(codingsequence,CDS)克隆和测序:根据NCBI提供的长爪沙鼠Cst3基因CDS区序列(GenBank: KM517575.1)两端设计Cst3基因CDS区克隆的上下游引物,上游引物序列为“5’-ATGGCTAGCCCACTACGATCC-3’”、下游引物序列为“5’-TTAAGCGCTTTTGCAGCTGGA-3’”,并用该引物将Cst3基因CDS区序列从长爪沙鼠脑组织cDNA中扩增出来,PCR扩增体系为50 μL体系,PCR扩增条件为:95 ℃预变性5 min;35个循环95 ℃变性30 s、57 ℃退火30 s、72 ℃延伸30 s;72 ℃ 继续延伸7 min后,4 ℃保存。将PCR产物进行琼脂糖凝胶电泳及胶回收后,将回收产物连接到pMD19-T载体上进行TA克隆测序,以获得长爪沙鼠Cst3基因CDS区序列。
1.3.3基因序列同源性比对:在NCBI上查询小鼠(NCBI Reference Sequence: NM_009976.4)和人类(GenBank: CR542018.1)Cst3基因CDS区序列,用DNAMAN软件将其与长爪沙鼠的Cst3基因序列进行比对,同时进行氨基酸序列比对。
1.3.4质粒载体构建: 去除长爪沙鼠CST3蛋白信号肽区域(1~20 aa)后,对剩余蛋白质全长的对应Cst3基因序列进行密码子优化、在N端加入His标签,并通过化学合成获得对应的基因片段,合成的片段通过NcoI和XhoI位点酶切、连接,将合成序列插入到pET28a质粒载体中。
1.3.5重组蛋白的诱导表达和检测:将pET28a-CST3质粒和pET28a空质粒载体分别转化到E.coliBL21感受态细胞中,涂板、克隆及培养。用30 mL卡那霉素抗性的LB培养基37 ℃扩增菌液,在对数扩增期加入IPTG诱导,使其终浓度为1 mmol/L。收集菌液后取全菌液上清和沉淀,分别进行SDS-PAGE凝胶电泳和Western blotting检测。
1.3.6SDS-PAGE凝胶电泳:将各组蛋白进行15% SDS-PAGE凝胶电泳,电泳条件为:80 V恒压电泳30 min,120 V电泳至结束。随后,用考马斯亮蓝染液染色过夜并进行脱色。
1.3.7Western blotting实验:将SDS-PAGE凝胶使用Bio-Rad蛋白质转膜系统将蛋白质电转至0.22 μm硝酸纤维素膜(nitrocellulose filter membrane,NC膜)上,用0.5%封闭用脱脂奶粉封闭后,使用His抗体(1∶1 000)4 ℃孵育过夜,TBST充分洗涤后,用辣根过氧化物酶(Horseradish peroxidase,HRP)标记抗鼠血清二抗室温孵育1 h。再次充分洗涤后,进行化学发光,用Bio-Rad凝胶图像分析系统进行图像采集分析。
2 结果
2.1 长爪沙鼠Cst3基因CDS区克隆测序分析
为构建CST3原核表达载体,我们首先以长爪沙鼠脑cDNA为模板,扩增了Cst3基因CDS区,并连接到pMD19-T载体上进行克隆测序。利用DNAMAN软件将测序结果与GenBank中长爪沙鼠Cst3基因CDS区序列比对。结果显示,长爪沙鼠Cst3基因CDS区长度为429 bp,我们克隆的长爪沙鼠Cst3基因CDS区序列与GenBank序列仅相差一个碱基(图1),并导致一个氨基酸的差异。
2.2 长爪沙鼠Cst3基因同源性分析
为判断长爪沙鼠Cst3基因序列同源性,我们利用DNAMAN软件分别将长爪沙鼠与人、小鼠Cst3基因CDS区序列进行比对(图2)。长爪沙鼠Cst3基因CDS区序列与人、小鼠的同源性分别为68.90%和79.49%。同时,我们也比较了长爪沙鼠与人、小鼠CST3蛋白氨基酸序列的同源性。结果显示,长爪沙鼠CST3蛋白氨基酸序列与人、小鼠的序列同源性分别为63.51%和71.83%(图3)。这些比对结果显示,长爪沙鼠Cst3基因与人、小鼠序列同源性较低,保守性较差。这提示,在进行长爪沙鼠CST3蛋白研究时,不宜使用小鼠或人类CST3蛋白或抗体。
2.3 长爪沙鼠CST3蛋白表达载体构建
为建立长爪沙鼠CST3蛋白体外表达体系,将长爪沙鼠CST3蛋白与人、小鼠CST3蛋白比对分析,结果表明长爪沙鼠CST3蛋白的1~20氨基酸(amino acid,aa)为信号肽区域,21~142氨基酸为CST3蛋白功能区域。由于长爪沙鼠CST3蛋白编码序列中稀有密码子较多,GC含量偏高,因此,我们在去除序列中的信号肽后,对CST3蛋白编码序列进行了密码子优化,更换了75个同义密码子(图4)。化学合成优化后的编码序列,并将其酶切连接到pET28a蛋白表达载体中构建CST3蛋白体外表达载体,以获得N端携带6X His标签的重组长爪沙鼠CST3蛋白。将质粒转化到E.coliDH5a感受态细胞中,并进行克隆测序,结果显示载体序列与预期一致,带有His标签的重组长爪沙鼠CST3蛋白表达载体构建成功(图5)。预计该重组蛋白总长为143个氨基酸,其中包含N端6X His标签和沙鼠CST3蛋白21~142 aa,分子量大约为15 kDa。

图1 长爪沙鼠Cst3 CDS区克隆测序与GenBank长爪沙鼠Cst3 CDS区序列比对结果Fig.1 The alignment analysis between Cst3 CDS sequences we cloned here andGenBank Cst3 CDS sequences of Mongolian gerbils
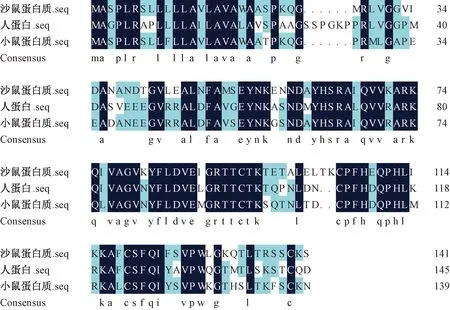
图3 长爪沙鼠与人、小鼠CST3蛋白氨基酸序列比对Fig.3 The alignment analysis of CST3 amino acid sequencesamong Mongolian gerbils, human and mice

图4 长爪沙鼠CST3蛋白编码序列密码子优化方案Fig.4 Codon optimization scheme for CST3 coding sequences of Mongolian gerbils

图5 长爪沙鼠CST3蛋白表达质粒测序结果注:下划线部分为CST3蛋白编码序列,加粗黑字为起始密码子和终止密码子,黄色阴影为T7启动子,红色为His标签Fig.5 The partial sequences of recombinant CST3 protein expression vectoNote: The underlined parts are CST3 protein coding sequences, the bold parts were the start codon and the stop codon, the yellow shade parts were the T7 promoter sequences, and the red font parts were the His tag sequences.
2.4 CST3蛋白体外诱导表达和鉴定
为优化蛋白体外表达条件,我们使用不同浓度和处理时间的IPTG对转入重组CST3蛋白表达质粒的E.coliBL21感受态细胞进行诱导表达。SDS-PAGE凝胶电泳显示,与对照相比,0.2 mmol/L IPTG诱导并无特异性蛋白表达,而1 mmol/L IPTG 37 ℃诱导表达12 h后在分子量约15 KDa 处出现单一目的条带。对蛋白进行Western blotting检测His标签表达情况,结果显示IPTG诱导组有特异性His表达,大小与重组CST3蛋白大小一致,并且His蛋白表达结果也表明1 mmol/L IPTG 37 ℃诱导表达12 h是诱导沙鼠CST3蛋白表达的最佳条件(图6)。综上,我们成功建立了CST3蛋白体外诱导表达系统,获得约15 kDa的重组沙鼠CST3蛋白。
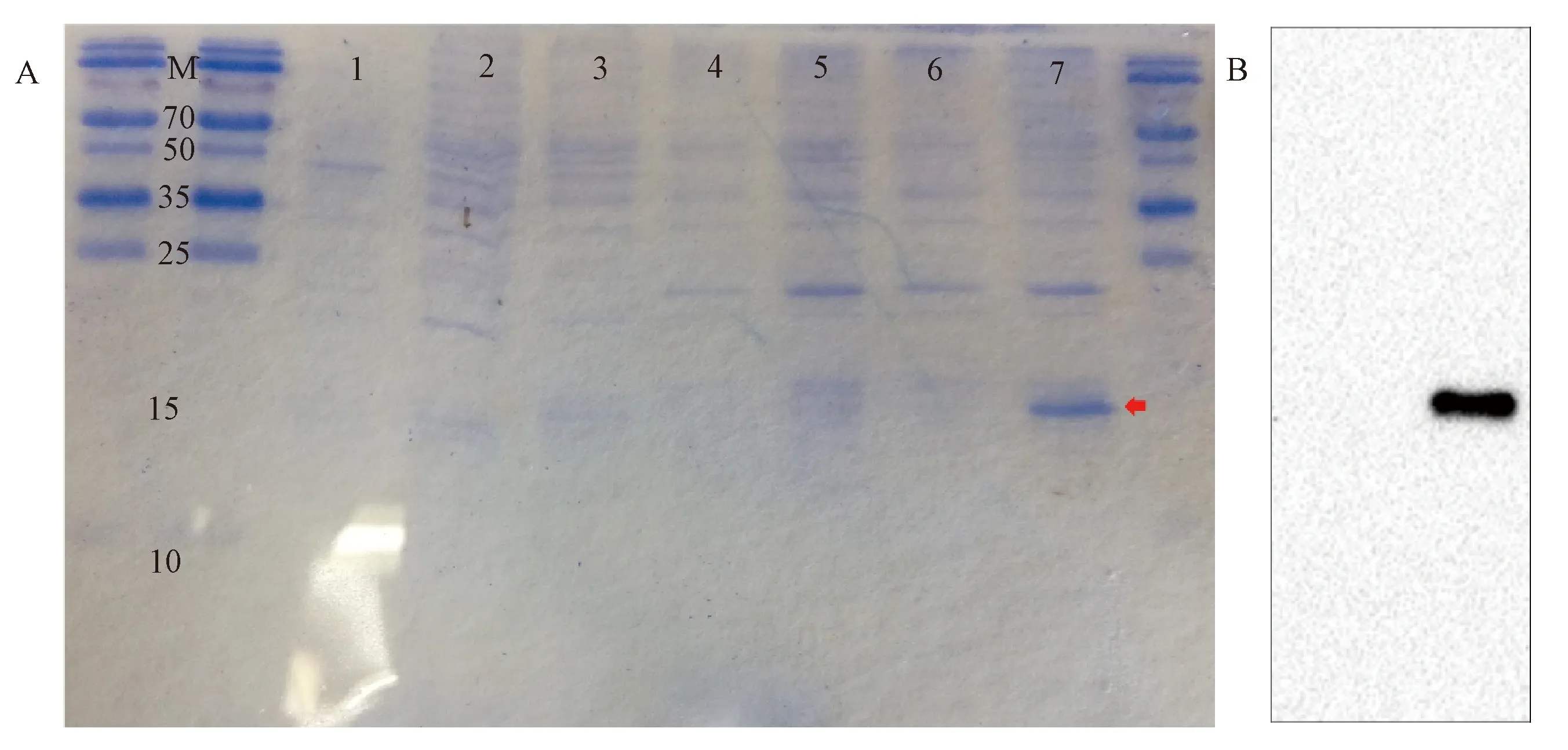
图6 长爪沙鼠CST3蛋白的体外诱导表达注:A:SDS-PAGE凝胶电泳结果;B:Western blotting结果;M:蛋白marker,1为pET28a载体;2、3、4分别为0.2 mmol/L IPTG 37 ℃诱导4 h、8 h和12 h,5、6、7分别为1 mmol/L IPTG 37 ℃诱导4 h、8 h和12 h,红箭头为目的条带Fig.6 Induced expression of gerbil CST3 protein in vitro.Note: A:SDS-PAGE electrophoresis; B:Western blotting;M:protein ladder; 1, pET28a vector; 2~4,induced by 0.2 mmol/L IPTG at 37 ℃ for 4 h, 8 h and 12 h, respectively; 5~7,induced by 1 mmol/L IPTG at 37 ℃ for 4 h, 8 h and 12 h, respectively.Red arrows indicated the target protein
3 讨论
半胱氨酸蛋白酶在多个生物过程,如在蛋白质转运,前蛋白加工,骨重塑,和细胞凋亡等过程中发挥重要作用[7-8],也参与多种病理过程,如心血管疾病和炎症等。因此,半胱氨酸蛋白酶的活性需要被抑制剂严格控制。CST3又称胱抑素C(Cys C),是半胱氨酸蛋白酶的一种重要的内源性抑制剂[9],属于蛋白酶抑制剂的半胱氨酸蛋白酶抑制剂II型超家族,是由Cst3基因编码、分子量为13 kDa的分泌蛋白。CST3是一种非糖基化碱性蛋白质,广泛存在于各种体液及组织的有核细胞中,参与调控细胞内外的蛋白水解酶,保护细胞使其免受细胞内和细胞外蛋白水解酶的水解,是目前发现的对组织蛋白酶B抑制作用最强的蛋白酶抑制剂[10]。血清CST3水平具有一定的诊断价值,是癌症、心血管疾病和炎性肺病等疾病的预后标志之一[11-12]。当肾功能和肾小球滤过率下降时,血清CST3水平显著升高。而循环中CST3水平降低则与动脉粥样硬化斑块形成有关。此外,CST3异常表达也参与肿瘤转移、自身免疫疾病、肝功能障碍和阿尔兹海默症的淀粉样蛋白沉积等疾病的发生发展[13-16]。人类Cst3基因位于第20号染色体短臂13区2上,总长约4.3 kb,包含3个外显子和2个内含子,编码146个氨基酸的CST3蛋白。而小鼠CST3蛋白有140个氨基酸,长爪沙鼠CST3蛋白总长为142个氨基酸。核酸和氨基酸序列比对结果显示,沙鼠CST3蛋白序列与人、小鼠CST3序列同源性都较低。因此,在进一步研究长爪沙鼠CST3基因功能时,特异性抗体和CST3蛋白不宜使用现有人源性或小鼠源性产品,建立长爪沙鼠CST3蛋白的体外表达体系十分必要。
Cst3是一个多态性非常强的基因,在人类CST3蛋白编码区就有223个单核苷酸多态性(single nucleotide polymorphism,SNP)位点。现有研究表明,多种Cst3基因的SNP位点与糖尿病、肥胖、冠心病、阿尔兹海默症和代谢综合征等疾病的发生发展相关[10,17-20]。在建立长爪沙鼠CST3蛋白体外表达体系时,我们克隆的封闭群长爪沙鼠Cst3 CDS区序列与GeneBank中Cst3 cDNA序列存在一个碱基的差异,且这一碱基的不同会导致CST3蛋白质序列的一个氨基酸改变。由核苷酸序列比对可知,这个差异碱基位点恰好对应的是人Cst3基因中的一个SNP位点rs1428071266,这提示长爪沙鼠中的这个差异位点可能是一个SNP位点。但是,目前尚无关于该位点功能的报道。
我们将密码子优化的CST3蛋白编码序列插入pET28a蛋白表达载体中构建了重组长爪沙鼠CST3蛋白表达质粒,并将该质粒转入到大肠杆菌原核表达系统中,通过IPTG诱导获得分子量大约为15 kDa N端携带6X His标签的重组长爪沙鼠CST3蛋白。但是诱导后获得的高效表达的CST3蛋白主要以包涵体形式存在,普通的镍柱层析纯化方法只能获得很少的可溶性长爪沙鼠CST3蛋白[21],不足以进行后期抗体制备等实验。因此,后续若进行蛋白制备,可考虑用变剂溶解,纯化复性后获得CST3蛋白,以用于抗体制备和进一步研究。综上所述,本研究对长爪沙鼠Cst3基因序列同源性进行了分析,并成功构建了重组CST3蛋白表达载体,为长爪沙鼠Cst3基因功能研究奠定了基础。
