25年生湿地松家系基本产脂力遗传分析与选择
2019-05-08吴东山杨章旗黄永利
吴东山,杨章旗,黄永利
(1.广西壮族自治区林业科学研究院 国家林业和草原局马尾松工程技术研究中心广西马尾松工程技术研究中心,南宁 530002;2.南宁市林业科学研究所,广西南宁 530107)
湿地松(Pinus elliottii)原产于北美东南部,是美国主要造林树种之一。上世纪70年代开始在我国大面积推广种植,目前已成为我国南方丘陵地区主要造林树种之一[1]。湿地松与本土马尾松(Pinus massoniana)比较,具有高产脂量的优点,这是其引种并大面积推广的重要原因[2]。关于湿地松高产脂的遗传研究也成为了关注的热点[3-7]。吕鹏信等[8]通过产脂力研究湿地松产脂变异时指出,产脂力是由林木内在遗传因素控制的,是松脂产量大小的决定因素,从而避免了生长量及环境因子等因素差异造成的产脂力与生长量之间的弱相关或失去实用意义的情形[9]。因此,产脂力对产脂性状的选育有重要的应用价值。目前国内对湿地松产脂性状的研究多集中在幼林期和中林龄阶段[10-11],对成熟林龄期家系的产脂遗传研究不多,更缺乏对这一阶段产脂力稳定性的系统评价。本研究利用31个湿地松半同胞家系25年生时的采脂测定材料及其对照家系,结合SAS程序中非平衡数据分析方法对家系基本产脂力进行遗传力估算,进行年度内性状相关分析,计算家系基本产脂力的现实增益和遗传增益,为高产脂湿地松家系选育提供参考。
1 材料与方法
1.1 试验材料
试验地位于广西南宁市林科所国家马尾松良种基地(108°00'E,23°10'N),海拔120 m左右,地势平坦。年均温21.5℃,1月均温12.5℃,极端最低温-2.5℃,7月均温29.7℃,极端最高温40.6℃,年均有霜日23 d,年均降水量1 246 mm,年均蒸发量1 613.8 mm,夏湿冬干,属热带北缘季风气候,全年平均相对湿度79%。土壤为第四纪红土发育而成的中壤质厚层赤红壤,土层厚1 m以上,pH值4.5~5.0。
试验材料为广西南宁市林科所湿地松优树子代家系测定林,1991年育苗,1992年采用随机区组设计营造,共80个试验号,包括77个子代家系,3个对照(CK)(103、104、105),单行6株小区,6次重复。选取31个子代家系于2015年进行采脂试验,另取一区组相同家系同分株数同年采脂作为对照。
1.2 试验方法
采脂前测定树高、胸径等指标,在树干1.3 m 处采用下降式双面采脂法[12],采脂测定时间为2015年7—11月,每1 d采割1次,每10 d称量1次。采脂结束后测定树高、胸径、割脂面的水平长度、割面处树干周长、割沟长等。割面宽与采割处周长均在第1刀"V"形底部测量。本研究主要讨论遗传因素对产脂力的作用,为消除树木间生长量及采脂操作带来的不同采割负荷率差异所导致的误差,采用基本产脂力评价湿地松产脂能力,即每天单位水平割面长度(1 cm)的产脂量(g·d-1·cm-1)。
1.3 统计分析方法
半同胞家系测定林中单株观测值的线性模型[13]如下:
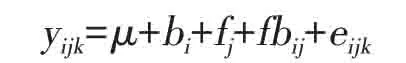
式中:yijk表示i个重复第j个家系的第k个观测值;bi为区组效应;fj为家系效应;fbij为家系与区组的交互作用,eijk为小区内效应。

表型变异系数(CV)和遗传变异系数(GCV)公式:
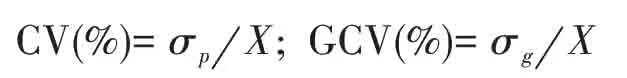
式中,σp、σg、X分别表示表型标准差、遗传型标准差、均值。
遗传增益:∆G=
式中,∆G为遗传增益,S为选择差,h2f为家系遗传力,X为性状群体平均值。
现实增益:∆G实=(xi-x)x
式中,∆G实为现实增益,xi为各家系平均值,x为对照平均值。
所有统计均采用家系均值数据,因试验林小区中家系存在缺株情况,故利用SAS 统计软件中的VARCOMP 过程和限制性最大似然法(REML)对不平衡数据进行方差成分无偏差估算,利用TYPEI类方差进行平方和计算。MODEL方程采用混合模型[14]。
2 结果与分析
2.1 湿地松家系基本产脂力的变异
25年生湿地松处于成熟林龄期,产脂能力达到较高的稳定值,基本产脂力为2.14 g(表1)。基本产脂力变幅为0.92~3.14 g,表型变异系数低于15%,遗传变异系数低于10%,说明25年生湿地松基本产脂力相对稳定,显示出家系间较大的遗传差异和遗传因素对基因与基本产脂力的较强控制。

表1 家系基本产脂力表型变异Tab.1 Phenotypic variation of basic resin productivity in families
家系均方大于区组均方,且基本产脂力仅在家系间表现出极显著差异(P<0.01),说明家系是基本产脂力主要的影响因素(表2)。这为产脂力的家系选择提供了可能。遗传方差分量结果也印证了方差分析的结论。产脂力家系和单株遗传力分别为0.601 7和0.393 4,家系遗传力高于单株遗传力52.95%,从一定程度上反应出成熟林龄期湿地松产脂在遗传控制上的家系主导特性。

表2 家系基本产脂力方差分析和遗传估算Tab.2 Analysis of variance and genetic estimation of basic resin productivity in families
2.2 基本产脂力的协方差分析
对31个家系水平及个体单株基本产脂力的相关性进行协方差分析,所选择的模型效应极显著(P<0.001),回归关系极显著(P<0.001)(表3)。
25年生时,除了家系对基本产脂力的影响达到极显著(P<0.01)外,单株的影响也达到显著水平(P<0.05),其原因可能是采脂的间隔期过长,割口的机械创伤已经愈合,不能对松脂的持续分泌产生刺激效果(表3)。这表明,在成熟林龄的一定阶段,在单株个体水平进行高产脂力优良材料的选择也是可行的。

表3 25年生湿地松基本产脂力的协方差分析Tab.3 Covariance analysis of basic resin productivity at 25-year-old Pinus elliottii
2.3 相同家系不同采割时间基本产脂力的比较
为了解采脂割口愈合后,再次进行采脂时基本产脂力与未采脂林木是否存在差异,以及2种类型林木对基本产脂力的影响,取另一区组相近树体条件下的未采脂相同家系同分株数量作为对照,进行采脂试验,对应比较基本产脂力均值、总产脂量(表4)。
间隔3年后再次进行采脂与同立地条件下同家系新采脂对照比较,家系间基本产脂力和总产脂量有明显的差异,29%的家系(3、15、31、56、68、78、85、89和106号)基本产脂力提高幅度在2.72%~27.36%之间,其余家系的基本产脂力均存在不同程度的下降,降幅在-1.08%~-54.43%之间;松脂总产量与基本产脂力的表现不一致,19%的家系(12、18、29、69、83和87号)基本产脂力下降,但总产脂量高于同家系对照。这是因为总产脂量是基于潜在产脂力条件计算的,与胸径、材积等生长性状和冠幅等形质性状有关。与3个CK 比较,全部家系的基本产脂力及总产脂量均高于CK。

表4 相同家系不同采割类型基本产脂力及总产量比较Tab.4 Comparison of basic resin productivity and total yield of different tapping types in the same family
2.4 湿地松基本产脂力增益及高产脂家系选择
基本产脂力方差分析及同家系不同采割类型基本产脂力比较表明,在家系水平,基本产脂力存在极显著差异,且基本产脂力显著大于对照,这为基于基本产脂力的湿地松高产脂家系选择提供了可能。为了解湿地松在成熟林龄初期和中期产脂力增益的变化,选取湿地松在21年及25年生时的产脂数据进行分析,2年间家系基本产脂力现实增益及遗传增益的家系排序不一致,25年生时家系平均基本产脂力是21年生时的1.17 倍,按遗传增益从大到小排序,21年生时基本产脂力遗传增益大于10%的家系有31、68、15、32、17、56、69和12号,25年生时基本产脂力遗传增益大于10%的家系有54、55、87、17、24、13、12、8、32、31、69和68号(表5)。按现实增益从大到小排序,21年生时基本产脂力现实增益大于10%的家系有31、68、15、32、17、56、69、12、54、89、55、53和87号,25年生时基本产脂力现实增益大于10%的家系有54、55、87、17、24、13、12、8、32、31、69、68、86、20、56、53、73和91号。
综合2个年度的遗传增益及现实增益,以2年的平均现实增益20%以上、平均遗传增益10%以上为标准,表现优良的家系为31、54、17、55、32、68、12和69号家系,占全部参试的25.81%。2年间其家系平均现实增益分别为34.48%、32.35%、31.86%、30.98%、25.76%、25.66%、23.17%和22.26%;家系平均遗传增益分别为19.31%、16.93%、16.91%、14.29%、12.43%、12.95%、14.31%和13.70%。以基本产脂力为基础选择出的这8个家系可作为高产脂材料进行高世代繁育。
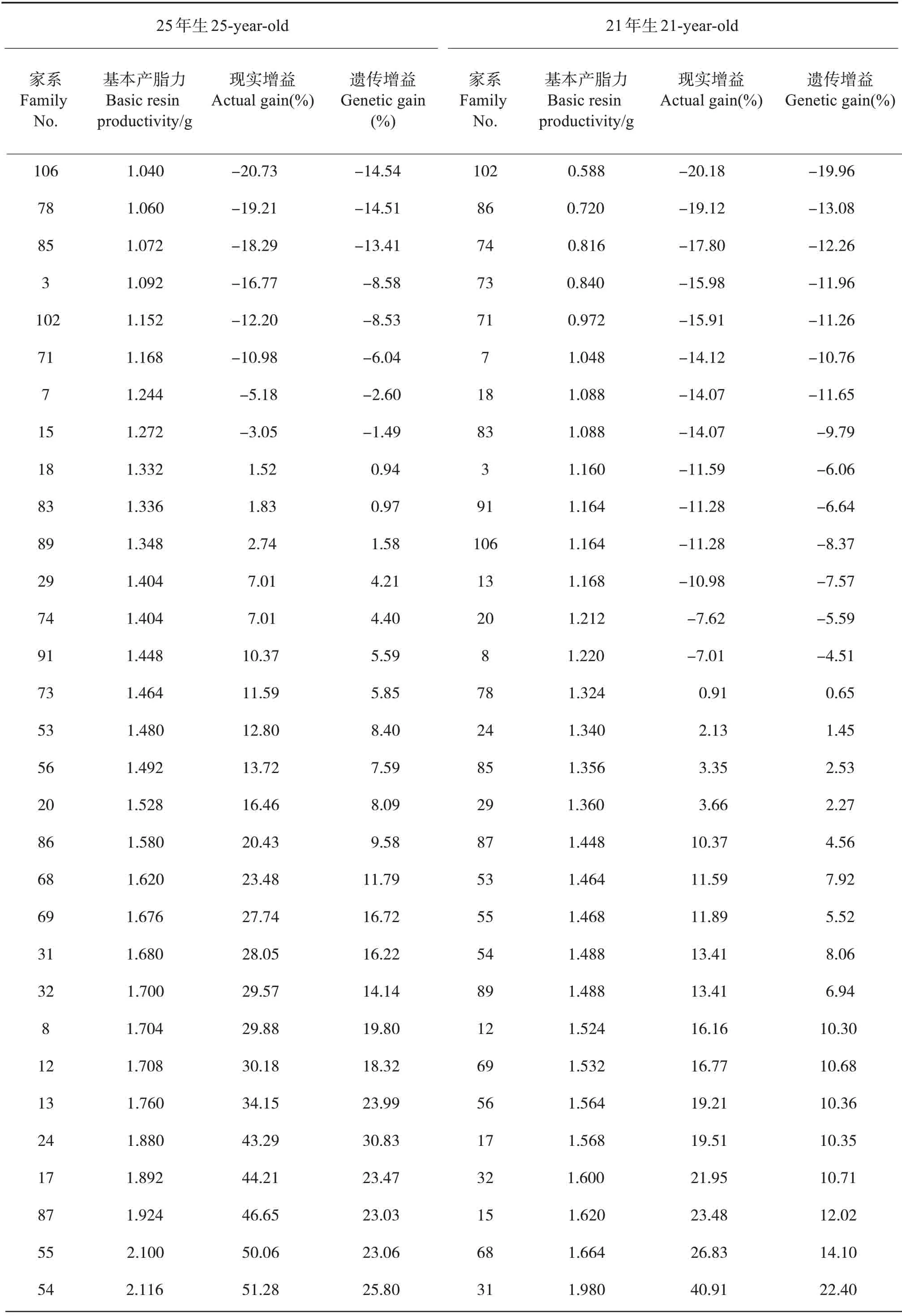
表5 年度间家系基本产脂力现实增益及遗传增益Tab.5 Actual gain and genetic gain of family basic resin productivity in two years
3 结论与讨论
在国内,湿地松基本产脂力研究早期较为典型的为吕鹏信等[8]研究3~5年生湿地松幼龄子代产脂力试验,得出树木的潜在产脂力主要由基本产脂力决定、基本产脂力受较强遗传因素控制的结论。本文以成熟林龄期湿地松家系为研究材料进行基本产脂力的分析,方差分析表明,基本产脂力在家系间的变异极显著,且从家系遗传方差分量及家系遗传变异系数占总变异系数百分比来看,成熟林龄基本产脂力的家系遗传效应显著,家系遗传力处于中等以上水平(0.601 7),这一结论与吕鹏信等[8]的研究结果一致。由于材料中存在不足,特别是在进行引种湿地松高产脂育种群体的早期选择评价时,缺乏美国学者们[16]的完整测定数据,使研究价值有所降低。
在基本产脂力的年度内协方差分析上,本文得到的结论与张建忠等[9]的结论有所不同。张建忠等[9]对15~16年生中林龄湿地松进行连续2年采脂,年度间产脂量极显著相关,本文研究的成熟林龄期湿地松是间隔3年再次进行采脂,可能是割口的机械损伤刺激减弱,年度内的产脂力为显著相关,结论不同可能与本文的研究对象是基本产脂力,未考虑林木生长、形质等指标的综合因素有关。潘显强[15]在研究年度内的产脂量相关性时侧重于总产量与月份产量的紧密性,本文从基本产脂力出发,指出在成熟林龄的一定阶段内,在家系水平上,家系效应是影响基本产脂力的主要效应。家系内单株对基本产脂力的影响不显著,而年度内家系遗传力及单株遗传力的差异比张建忠等[9]研究中中林龄湿地松的数值要大。
本文初步讨论了成熟林龄期湿地松间隔采脂基本产脂力的表现,在同家系同分株数量的同等条件下比较新采割与再次采割2种树体类型的基本产脂力及总产脂量,发现仅有29%的家系基本产脂力高于对照,19%的家系基本产脂力低于对照但总产脂量却高于对照,一定程度上印证了本文协方差分析的结论。初步的结论为在家系水平上间隔3年再次进行采脂,基本产脂力及总产脂量均有所降低。进一步印证需进行多年连续采脂补充材料说明。
由于本文仅讨论遗传因素控制较强的基本产脂力性状,为此在进行高产脂家系选择时,侧重考虑产脂力的现实增益及遗传增益表现,并对2个年度内的2种增益进行分别排名。基本产脂力现实增益与遗传增益均表现优良的家系为31、54、17、55、32、68、12和69号家系。通过计算现实增益与遗传增益,可知现实增益稍高于遗传增益。选出的家系均具有较高的遗传增益,这为高产脂改良提供了良好的基础材料。
