闽北不同海拔湿地松生物量模型构建及比较分析
2017-08-31魏年锋
魏年锋
(福建省林业勘察设计院,福建 福州 350001)
闽北不同海拔湿地松生物量模型构建及比较分析
魏年锋*
(福建省林业勘察设计院,福建 福州 350001)
对福建省政和县石屯镇、杨源乡10年生湿地松幼龄林建立多个生物量相容模型,并对不同海拔的湿地松幼龄林生物量比较研究。结果表明:不同海拔的湿地松幼龄林生长差异明显,胸径、树高差异显著,300m海拔区比900m海拔区的平均树高、胸径大;其分别建立的900m海拔区和300m海拔区生物量相容性模型决定系数R2>0.99;300m海拔区10年生湿地松单位面积生物量比900m海拔区高97%。说明海拔对湿地松生物量影响显著,较低海拔区更利于湿地松生长。
海拔;生物量模型;湿地松
湿地松(Pinuselliottii)原产于美国东南部,于20世纪30年代引种到我国,由于其具有抗旱耐涝、生长快、出材质量好、松脂量丰富等特性,已成为我国南方丘陵山区的主要造林树种之一,特别是以丘陵山区为主的闽北地区。生物量作为一种量化指标,对植物的生长评判、立地条件的反应等方面有重要作用[1]。目前国内外对湿地松的生物量做了较为丰富的研究,如Gonzalez[2]、刘海[3]等研究了湿地松的生物量分布特征,肖兴翠[4]、Burkes[5]等研究了不同林分密度的湿地松幼龄林生物量,曾伟生等[6]建立了湿地松的生物量相容性模型;李大岔等[7]、王舒凤等[8]研究了湿地松混交林的生物量组成情况;郑丽垠[9]对不同坡位的湿地松生物量做了分析。但是不同海拔对湿地松幼龄林生物量的影响及比较研究相对较少。湿地松幼龄林时期的生长对未来采脂、出材等都极为重要,其生物量的存储及变化情况受到多方面的制约与促进,比如土壤条件、水热条件、地形因子等。本文以闽北地区10年生湿地松幼龄林为研究对象,重点关注不同海拔对生物量的影响,研究结果对湿地松的人工种植在海拔的选择上有重要指导意义。
1 材料及方法
1.1 研究区概况
研究区位于福建省政和县的石屯镇和杨源乡。石屯镇位于政和县中西部,平均海拔<300 m,年均降雨量1 468 mm,年均温19.1 ℃;杨源乡位于政和县东南部,平均海拔>900 m,年均降雨量1 639 mm,年均温15.5 ℃。300 m海拔区样地位于石屯镇,900 m海拔区样地位于杨源乡。
1.2 研究方法
1.2.1 样地布设与样本选取
于900 m海拔区和300 m海拔区分别选取两块湿地松纯林设置样地,所选样地坡位、坡度等地形情况基本一致。两区湿地松均为2008年造林,造林密度2 500 株/hm2,苗木情况基本一致。在两片林分内各设置2块20 m×20 m的样地,并于2017年4月对样地内的湿地松进行每木检尺(表1)。

表1 样地内湿地松基本情况
在两片不同海拔的林分内各选取5株平均标准木,测量树高、胸径。同时按照“分层切割”进行现场取样,按根、枝、叶、干称取鲜重并分别保存。根部采用“全挖”称鲜重,其余各部分按照1%的比例称重带回实验室,采用80 ℃恒温烘干至恒重,计算不同样品的含水率及干重,获得标准木生物量。
1.2.2 数据处理
国内外学者提出了许多高精度的生物量经验模型[10-12]。曾伟生等[5]采用非线性二元联立方程组建立湿地松生物量相容性模型,决定系数超过0.9。不论是传统回归模型还是生物量与蓄积量相容模型,公式中都含树高、胸径[13]。本次试验选择非线性二元立木生物量方程计算生物量:
M=aDbHc
(1)
式中:M代表生物量总量,a、b、c为系数,D是胸径,H是树高。
利用外业采集的树高、胸径以及实验室实测的生物量数据,修正生物量方程系数,获得10年生湿地松幼龄林的生物量模型。
2 结果与分析
2.1 不同海拔立木生物量
通过对平均标准木的测量分析,得到不同海拔区树种的树高胸径信息,并对结果做显著性检验(表2)。比较发现闽北不同海拔的10年生湿地松幼龄林树高、胸径都存在显著差异(P<0.01),其中300 m海拔区湿地松的平均树高比900 m海拔区的高1.8m;300 m海拔区湿地松的平均胸径比900 m海拔区的大1.9cm。但300 m海拔区的湿地松树高、胸径的标准差也相应大于900 m海拔区湿地松,说明湿地松在300 m海拔区长势较好,而900 m海拔区的湿地松长势较为均衡。

表2 不同海拔标准木树高胸径对比
注:**为P<0.01水平显著
通过实验得到的样本含水率、生物量计算获得不同海拔的立木生物量,并对结果做显著性检验(表3)。
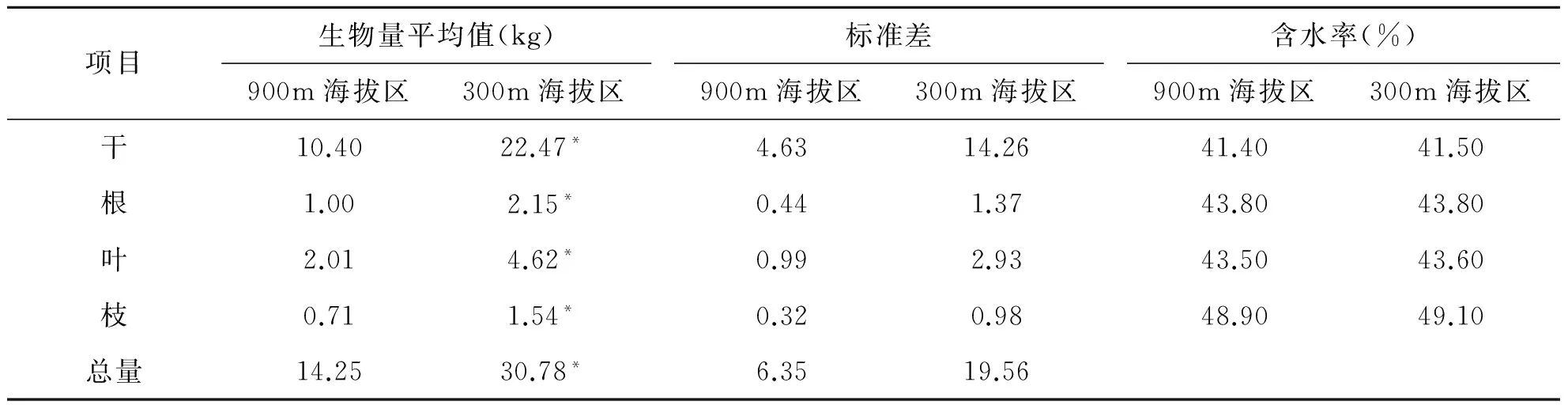
表3 不同海拔生物量及含水率
注:*为P<0.05水平显著
通过称量、烘干、统计发现300 m海拔区的10年生湿地松立木生物量平均值均高于900 m海拔区湿地松生物量,其中涵盖不同部分之间的生物量平均值与总量的生物量平均值,且300 m海拔区与900 m海拔区之间的10年生湿地松单株立木生物量呈现显著差异(P<0.05)。不同海拔区叶部分的生物量差异最明显,900 m海拔区的10年生湿地松叶部分生物量为300 m海拔区处的43.5%,其余部分都>46%;300 m海拔区10年生湿地松样本的生物量标准差大于900 m海拔区湿地松的样本;不同海拔之间的干部含水率最低,枝部分含水率最高。从表2、表3中可以得出,不同海拔间的树高、胸径、生物量都存在显著差异,因此,生物量相容性模型应该按照不同海拔研建。
2.2 不同海拔湿地松生物量模型建立及比较
通过采集的树高、胸径、生物量按不同海拔区分,在Origin软件内拟合非线性二元函数,最终获得政和10年生湿地松以及其不同海拔10年生湿地松生物量方程如下:
MT=0.06119D2.47724H0.2634(R2>0.99)
(2)
ML=0.0898D2.22595H0.3735(R2>0.99)
(3)
MH=0.09302D2.21868H0.31927(R2>0.99)
(4)
其中:式(2)、(3)、(4)中MT、ML、MH分别代表该地区10年生湿地松生物量、300m海拔区10年生湿地松生物量、900m海拔区10年生湿地松生物量,D代表胸径,H代表树高。
不同海拔的生物量模型决定系数均为R2>0.99,整体高于曾伟生等[5]地下部分0.90,地上部分0.97的研究结果。
结合式(2)、(3)、(4)与表1情况,计算不同海拔湿地松幼龄林生物量,所得政和10年生生湿地松生物量为46.81 t·hmm-2,其中900 m海拔区10年生湿地松生物量为31.69 t·hmm-2,300 m海拔区10年生湿地松生物量为61.37 t·hmm-2。900 m海拔区的10年生湿地松生物量为政和10年生湿地松生物量值的68%,300 m海拔区10年生湿地松生物量比政和10年生湿地松生物量值高31%,300 m海拔区10年生湿地松生物量比900 m海拔区10年生湿地松生物量高97%。
3 结论与讨论
不同的海拔决定和影响着环境中的许多因子,如降水、气温、土壤等,同样影响着湿地松的生长。对300 m与900 m海拔处的10年生湿地松幼龄林生物量,及相容性模型的建立进行研究,发现不同海拔的湿地松有明显的生长差异。由于不同的海拔造成的土壤养分、积温、降水等方面的差异,使得300 m海拔处的环境更适合种植湿地松,但该区域湿地松立木之间长势的差异增大。
在前人研究的基础上,重点研建了10年生温地松幼龄林的整体生物量模型与不同海拔间的生物量模型,结果发现三者的系数差异较大,说明不同海拔的10年生湿地松长势存在较大差异,造成了生物量模型差异较大。模型的决定系数极高,究其原因,本次研究采用同一县域内的10年生幼龄林,其生长环境比较接近,同时龄级一致。通过调查分析,将不同海拔的湿地松生物量分别建模,最终提高了决定系数。
就10年生湿地松幼龄林单株立木而言,300 m海拔区的单株立木平均生物量大于900 m海拔区,并且300 m海拔区10年生湿地松的各部分生物量都大于900m海拔区。标准差中,900 m海拔区的湿地松生物量较小,结合调查数据结果计算得到900 m海拔区10年生湿地松生物量为31.69 t·hmm-2,这与曾伟生等[14]建立的马尾松生物量模型计算结果接近(32.00 t·hmm-2),证明研究区900 m海拔区湿地松生长情况正常。300 m海拔区10年生湿地松生物量为61.37 t·hmm-2,比900 m海拔区10年生湿地松生物量高出97%,同时其林分保留密度也较大,说明300 m海拔区条件更适合湿地松生长。300 m海拔区的湿地松含水率高于900 m海拔区样本,但是900m海拔区的10年生湿地松长势更平均。
综合分析表明,300 m海拔区的环境中10年生湿地松的平均树高、平均胸径、每公顷生物量、存活率等指标较900m海拔区更大,湿地松在闽北更适应300m海拔区环境。后期将就不同海拔湿地松生物量差异驱动力、定量分析海拔对湿地松生物量差异影响等方向做深入研究。
[1] 田大伦,项文化,闫文德.马尾松与湿地松人工林生物量动态及养分循环特征 [J].生态学报,2004,24(10):2207-2210.
[2] Gonzalez-Benecke,Carlos A. Jokela, Eric J. Cropper,et al. Parameterization of the 3-PG model for Pinus elliottii stands using alternative methods to estimate fertility rating, biomass partitioning and canopy closure[J]. 2014,327:55-75.
[3] 刘海,叶功富,朱祥锦,等.马尾松、湿地松幼树生物量及养分积累 [J].福建林业科技,2014(01):49-54.
[4] 肖兴翠,李志辉,唐作钧,等.林分密度对湿地松生物量及生产力的影响 [J].中南林业科技大学学报,2011(03):123-129.
[5] Burkes,Will, Barron-Gafford, et al. Biomass partitioning and growth efficiency of intensively managed Pinus taeda and Pinus elliottii stands of different planting densities[J]. FOREST SCIENCE,2003,49(2): 224-234.
[6] 曾伟生,姚顺彬,肖前辉.中国湿地松立木生物量方程的研建 [J].中南林业科技大学学报,2015(01):8-13.
[7] 李大岔.福建柏与杉木、湿地松混交林分结构和生物量的研究 [J].福建林业科技,2004(04):51-53.
[8] 王舒凤,郑郁善,陈雄,等.木麻黄湿地松混交林生物量研究 [J].福建林学院学报,2001(02):153-156.
[9] 郑丽垠.长乐沿海山地不同坡位湿地松生物量分析 [J].亚热带农业研究,2013(02):94-97.
[10] Parresol. Additivity of nonlinear biomass equations[J]. Canadian Journal of Forest Research 2001,31(5).:865-878.
[11] Huiquan B,John Turner, Marcia J. Lambert. Additive biomass equations for native eucalypt forest trees of temperate Australia[J]. Trees - Structure and Function. 2004,18(4): 467-479
[12] 曾伟生,唐守正. 东北落叶松和南方马尾松地下生物量模型研建 [J].北京林业大学学报,2011(02):1-6.
[13] 吕常笑,邓华锋,王少杰,等.马尾松不同区域相容性立木材积和地上生物量模型 [J]. 浙江农林大学学报,2016(05):790-797.
[14] 曾伟生,肖前辉,胡觉,等.中国南方马尾松立木生物量模型研建 [J].中南林业科技大学学报,2010(05):50-56.
魏年锋(1963-),男,福建罗源人,工程师,长期从事人工速生丰产林、特种用材林、珍稀阔叶林、经济林的培育管理工作,(E-mail)weinianfeng88@163.com。
S791.246
A
1004-2180(2017)02-0061-04
