一种简化的油菜杂交F1种子的生产方法
2019-05-08杨光圣辛强董发明洪登峰
杨光圣,辛强,董发明,洪登峰
一种简化的油菜杂交F1种子的生产方法
杨光圣,辛强,董发明,洪登峰
(华中农业大学作物遗传改良国家重点实验室,武汉 430070)
安全有效的制种方法是油菜杂交种应用的关键。通过回交转育获得抗(耐)磺酰脲类除草剂的自交不亲和系,将其作为杂交种父本。父本因其具有自交不亲和特性,自交不结实;因其具有抗(耐)磺酰脲类除草剂特性,在用磺酰脲类除草剂进行母本化学杀雄时,不受化学杀雄剂影响。因此,可以将父本与不育系或者化学杀雄的母本材料混合播种,实现杂交种父、母本的混播混收。通过本文提出的杂交制种方法,只要选育出兼有抗(耐)磺酰脲类除草剂和自交不亲和特性的父本,即可进行杂交种的快速组配。该方法将父、母本混播混收,操作简单,节约了劳动成本,提高了杂交种制种效率。
甘蓝型油菜;杂交种;制种;化学杀雄;自交不亲和
0 引言
油菜作为中国第四大农作物、第二大冬季农作物、第一大油料作物,在中国农业生产中占据着举足轻重的地位。菜籽油作为国产食用植物油第一大来源,占中国油料作物产油量的57.2%[1]。近十年来,中国油菜单位面积产量获得持续提升,主要得益于杂交油菜的大面积种植。目前,中国杂交油菜制种应用的授粉控制系统包括波里马细胞质雄性不育三系或两系、S45A和9012AB类型的核不育系统、化学杀雄及自交不亲和等,这些系统在制 种过程中均需要将父、母本按一定行比分行播种,分开收割,不仅费时费工,而且存在出错的风险,影响商品杂交种的纯度。为了提高制种效率,降低杂交种子生产成本,增加杂交种纯度,降低生产风险,笔者提出一种简化的油菜杂交F1种子生产方法。
目前,国内外研究中利用的油菜细胞质雄性不育类型有10多种,生产上应用比较广泛的有萝卜细胞质雄性不育类型(CMS)[2-3]、波里马细胞质雄性不育类型(CMS)[4]等。其中,CMS被认为是国际上第一个具有实用价值的油菜雄性不育系,也是中国唯一在生产上大面积应用的细胞质不育类型。杨光圣等[5-7]1990年在CMS三系的基础上选育出温敏生态型细胞质雄性不育两用系,其在相对高温条件下表现雄性不育,在相对低温条件下表现雄性可育。该不育两用系可能是通过细胞核内的温度敏感基因来调控线粒体基因的转录模式或RNA的加工来实现育性变化的[8]。该系统恢复基因编码一个PPR蛋白,在转录后水平降低的积累量,从而恢复不育系育性[9]。目前,笔者利用该系统已选育出195A、206A、616A等多个品质性状、经济性状和农艺性状均表现优良的生态型雄性不育两用系,培育出华油杂10号等20多个高产优质杂交油菜新品种。
目前,生产上应用的油菜细胞核不育主要有3种:(1)隐性细胞核不育,主要有双隐性重叠基因控制的S45A[10]和2对隐性互作基因控制的9012A[11];(2)显性细胞核不育,如宜3A类型[12];(3)人工创制的转基因细胞核不育系统,如SeedLinkTM系统[13]。在中国生产上利用的主要是隐性细胞核不育系统,其中,由于S45A类型不育系在制种时,需要拔除母本行50%的可育株,极大地限制了它在生产上的应用。9012A类型核不育系统因具有不育性稳定彻底、恢复源广、无负胞质效应、可通过引入临保系产生100%不育群体等优点,被认为是一个具有较大应用潜力的授粉控制系统。后来,国内科研人员对该不育系统的遗传模式进行了修正[14-16],并克隆了其育性控制基因,开发出多个可以应用于育种的分子标记,解析了其雄性不育的机制[1-20]。通过该不育系统的三系配套已选育出多个杂交种,如安徽省农业科学院选育的部分皖油和核优系列组合、上海市农业科学院选育的部分沪油杂系列组合、浙江省农业科学院选育的浙油杂系列组合、华中农业大学选育的华油杂系列组合等。
化学杀雄剂(也叫化学杂交剂)是一类可以诱导雄性不育的化学试剂。目前,油菜生产应用中的杀雄剂主要包括氯代脂肪酸类、无机盐类、激素类、甲基胂酸盐类以及应用最广的磺酰脲类除草剂、氨基磺酸类药物。因其在杂种优势利用时,具有亲本选择选配范围广、育种周期短等优势,并可用于解决细胞质不育系存在微粉等问题,在作物杂种优势利用中获得广泛应用。近十多年来,已审定和登记了一系列杂交油菜品种(如湘杂油1号、湘杂油6号、秦杂油19、华油杂68以及中国含油量第一个超过50%的秦杂油4号等)。近年来,磺酰脲类除草剂是在中国应用最多最成功的化学杀雄剂,其化学杀雄的生物学机制已获得较好的阐释。叶面喷施的低剂量苯磺隆通过极性运输在花药中积累,特异性靶定乙酰乳酸合成酶(acetolactate synthetase,AHAS),抑制支链氨基酸的合成,最终诱导花药自体吞噬型细胞死亡,形成雄性不育[21-22]。
自交不亲和是指植物虽然能产生正常功能的配子,但不能自花授粉或者授粉后不能结实的现象。芸薹属植物中自交不亲和性由一个S位点调控,该位点包含2个复等位基因:一个柱头特异表达的S位点受体激酶(S-locus receptor kinase,SRK)基因和一个位于花粉外壁的受体蛋白SP11(S-locus protein 11或称为S-locus cysteine-rich,SCR)基因[23]。由于S位点的基因几乎不发生重组,而作为一个遗传单位进行传递,因此,将其称为S单倍型(S haplotype)[24]。芸薹属作物中,白菜和甘蓝以及人工合成的甘蓝型油菜表现为自交不亲和,生产上应用的甘蓝型油菜表现为自交亲和。S1300是研究较多的一个甘蓝型油菜自交不亲和系,转育自甘蓝型油菜和白菜杂交育成的自交不亲和系271[25]。研究发现,甘蓝型油菜自交亲和系的S单倍型在C基因组上是S单倍型,A基因组上S单倍型为和;S1300的自交不亲和性由A基因组S单倍型控制,与白菜S单倍型同源,其对应的恢复系和保持系S单倍型分别为A基因组的和[26-31]。虽然甘蓝型油菜自交不亲和以及自交亲和形成机理还未获得完全解析,但这些S位点基因的克隆和序列分析,为育种家提供了遗传转育优良的分子标记,这将加快自交不亲和在油菜杂种优势利用中的应用进程。
虽然上述杂种优势利用的方法在杂交种制种过程中获得广泛应用,但父、母本均需按一定行比分开种植,分别收割,费时费力,同时存在父本混杂的风险。
本研究提出一种简化油菜杂交种生产的方法,利用转育的抗(耐)磺酰脲类除草剂的自交不亲和系为父本,与各种类型不同来源的母本材料按一定比例混合,直接将母本和父本混播混收。该方法不仅操作简单,而且能节约劳动成本和提高种子纯度。
1 构想
首先需要转育获得抗(耐)磺酰脲类除草剂(herbicide resistance,HR)、自交不亲和(self- incompatibility,SI)的父本(恢复系或自交系品种),具体选育方法是通过回交转育和分子标记辅助选择的方法,将自交不亲和基因和乙酰乳酸合成酶突变基因或或+聚合到生产油菜杂交种的父本中,将父本变成抗(耐)磺酰脲类除草剂的自交不亲和系;同时采用同样的方法将自交不亲和保持基因BnS-7和乙酰乳酸合成酶基因或或+聚合到生产油菜杂交种的父本中,选育出抗(耐)磺酰脲类除草剂的自交不亲和同核保持系。这样,就可以用抗(耐)磺酰脲类除草剂的自交不亲和系和抗(耐)磺酰脲类除草剂的自交不亲和同核保持系繁殖出的F1实现杂交种父本的大规模繁殖。同时,还可以通过开花期喷施食盐水的方法繁殖抗(耐)磺酰脲类除草剂的自交不亲和系父本(图1)。无论采用哪种方法,均可以在苗期喷施磺酰脲类除草剂(正常除草浓度)来杀死杂株,以提高繁殖父本的纯度(图1)。
本方法使用的母本材料可以不受雄性育性的限制,根据母本的来源,将母本分为3种类型:(1)类型1,生产上广泛使用的全不育系,例如萝卜细胞质雄性不育系、稳定型CMS不育系、9012A类型的细胞核雄性不育系;(2)类型2,育性有分离或不稳定的不育系统,例如育性不稳定的CMS不育系,在传统制种过程中需要拔除一半可育株的S45A系统不育系等;(3)类型3,常规自交系或常规品种,即在传统制种中通过化学杀雄作为母本的自交系或常规品种。
在播种前,将母本(类型1、类型2或类型3)种子与其对应的同时具有抗(耐)磺酰脲类除草剂和自交不亲和特性的父本种子按一定比例(母本﹕父本=70%—90%﹕10%—30%)均匀混合。将混合的种子播种(人工撒播或条播、机械条播)于种子生产隔离区大田,苗期按常规方法进行田间去杂。对于类型1不育系材料,其不育性稳定且彻底,因此,制种时只需将父、母本种子按比例混匀播种即可。由于父本具有自交不亲和特性,因此不能正常自交结实,所以收获时可以直接混收,不需父、母本分别收割或者收获前砍掉父本(图2-A)。对于类型2和类型3母本,由于其具有可育花粉,需要在现蕾初期喷施含苯磺隆的化学杂交剂,15—20 d后再喷一次同样的化学杂交剂,进行化学杀雄,种子成熟时,将父、母本混收,获得F1杂交种子(图2-B和图2-C)。对于类型1母本材料,为了降低因亲本不纯或田间自生苗对制种种子纯度的影响,也可以参照上述方法,在田间喷施2次同样的化学杂交剂,将杂株和自生苗杀雄(图2-A)。

图1 简化油菜杂交种制种:父本转育及繁殖示意图

A:类型1母本,生产上广泛使用的全不育系。可在现蕾初期喷施含苯磺隆的化学诱导雄性不育剂,将杂株杀雄,提高制种纯度。B:类型2母本,育性有分离或不稳定的不育系统;C:类型3母本,常规自交系和常规品种
2 讨论
2.1 本构想产生的历史背景与研究进展
油菜现有的杂种优势利用途径都有自己的优点和缺点。同时,无论哪条途径,在生产F1杂交种时,需要将父、母本分开种植,分别收割。不仅费时费力,而且容易造成机械或人为混杂。因此,发掘一条更加简便通用的途径,简化油菜杂交种子生产过程,降低种子生产成本,尤其是降低种子生产风险,一直是笔者思考的重要问题。2013年春节,笔者脑海突然产生此构想。经过4年多的努力,于2017年成功选育出7-5STR(图3)等多个同时具有自交不亲和和磺酰脲类除草剂抗性的波里马细胞质雄性不育恢复系。目前,正在将所有重要杂交油菜品种的父本转育成同时具有自交不亲和和磺酰脲类除草剂抗性的材料。且于2017年开始了小规模种子生产试验,2018年已有7-5STR配制2个杂交油菜品种申报国家和湖北省区域试验。
2.2 与现有技术相比,本构想杂交制种方法的优点
(1)播种时,传统杂交种子生产方法是将母本和父本按一定行比分开种植,人工育苗移栽一般是先栽母本,后栽父本。人工直播一般是先播母本再播父本。机械播种要将母本与父本分行播种。这样不仅费工费时,而且常常出错。本构想将母本和父本混合种植,生产程序简单,节约劳动成本,省钱又省力;同时还不易出错。
(2)收获时,传统方法是将母本和父本分开收割,或提前将父本砍掉。提前砍父本,无疑会增加劳动力成本;将父母本分开收割,不仅会增加劳动力成本,而且常常会出错,或人为混杂,导致父本混入母本中,影响种子纯度。本构想直接将母本和父本混收,操作简单,不仅节约了劳动成本;同时由于父本具有自交不亲和特性,不能正常结实,从根本上减少了将父本种子混入F1种子的风险,有利于提高种子纯度。
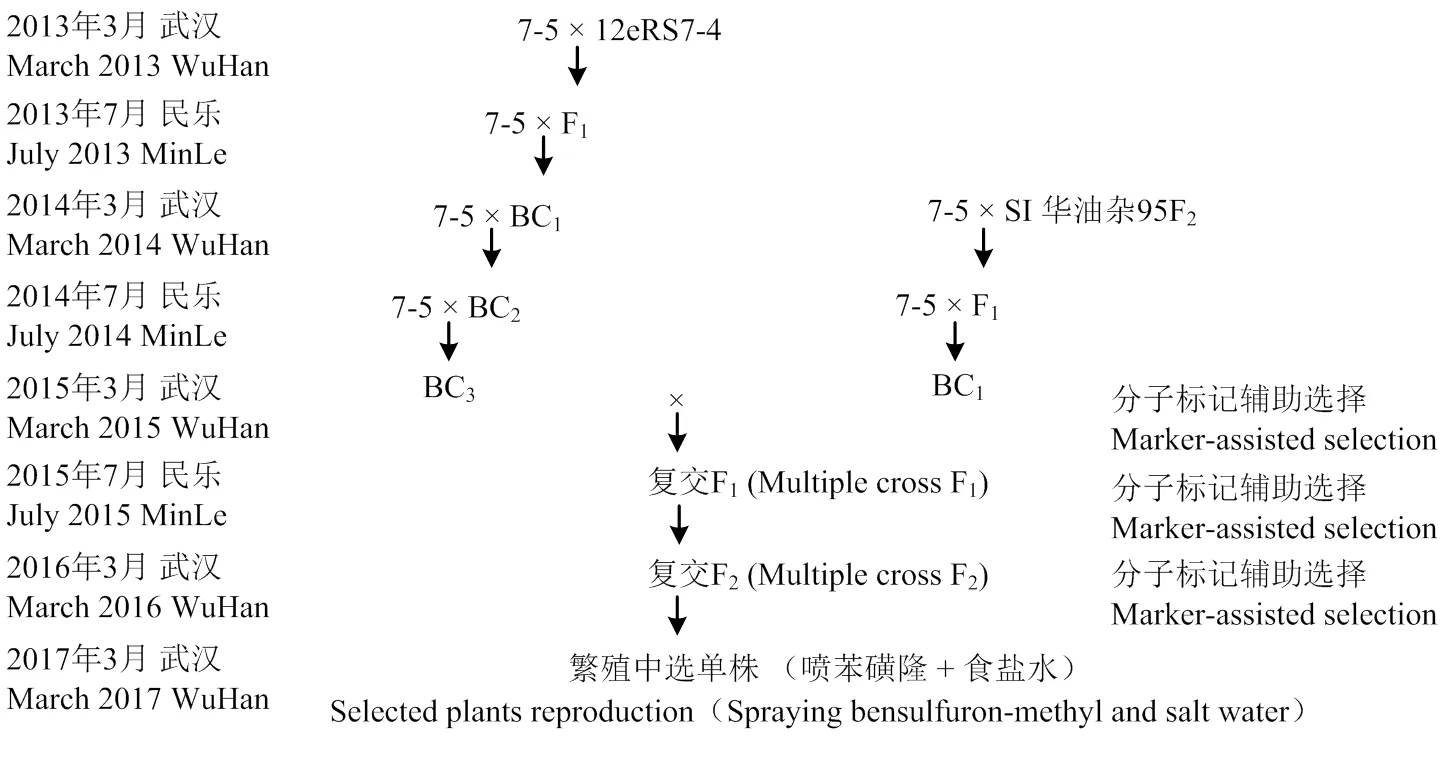
7-5:波里马细胞质雄性不育恢复系;12eRS7-4:抗磺酰脲类除草剂突变体;SI华油杂95F2:自交不亲和杂交品种华油杂95自交F2中的自交不亲和株
(3)与传统方法比较,由于本构想中的父本具有自交不亲和特性,它不能正常结实,开花期会延长,有利于给母本授粉,提高F1种子产量。不仅如此,对于父本开花期早或开花期短的杂交组合,传统的生产方法是将父本迟播或将父本打薹。迟播父本往往生长势弱、开花期变短,导致制种产量下降。将父本打薹,则会导致父本开花期缩短,比母本提前终花,母本后期开的花不能正常授粉结实,也会导致减产。
(4)传统方法中,当利用波里马细胞质雄性不育系制种时,母本行去杂不彻底会导致杂株给不育系授粉,影响杂种纯度;同时波里马细胞质雄性不育系育性易受温度影响,不育系的育性不稳定可能导致自交或兄妹交结实,影响杂种纯度。本构想可以通过喷施杀雄剂将母本行中的杂株和母本同时杀雄,消除了杂株给母本授粉和不育系自交或兄妹交的风险,有利于提高种子纯度,降低生产风险。
(5)传统方法中,化学杀雄制种时,给母本行喷施杀雄剂要避免喷到父本上,否则会将父本杀雄,减少父本花粉量,影响制种产量。而为了避免将杀雄剂喷到父本上,传统方法要采用一定的隔离措施。本构想由于父本带有磺酰脲类除草剂抗性基因,喷施苯磺隆不会影响父本的雄性育性,不需要采取隔离措施。这样不仅可以提高制种产量,而且还可以减少劳动力投入,提高制种效益。
(6)本构想利用的自交不亲和性在遗传上是隐性的,不会影响其杂交F1的自交亲和性。同时,由于隔离不严格或田间自生苗去杂不彻底,自交不亲和父本接受其他材料花粉授粉受精所产生的后代也会表现自交亲和。
2.3 与现有技术相比,本构想的不足
(1)由于要选育具有自交和亲和特性和磺酰脲类除草剂抗性的父本,父本的选育程序比传统方法复杂,且选育周期可能会长一些。为了弥补这一缺陷,可以将分子标记辅助育种和全基因组选择应用到父本的选育过程中。
(2)父本的繁殖纯度及自交不亲和性的稳定性会影响杂交种的制种纯度。要提高父本的繁殖纯度,一是要严格按照亲本繁殖程序进行父本的繁殖;二是可以在苗期通过喷施磺酰脲类除草剂杀死杂株。要保持父本自交不亲和性的稳定性,一是要选育自交不亲和性稳定的父本;二是要严格按照亲本繁殖程序进行父本的繁殖,防止生物学混杂;三是建议采用DH系,减少剩余变异的影响;四是建议通过喷施食盐水繁殖父本,不用保持系繁殖父本。
(3)自交不亲和父本并不是完全自交不结实。不同的自交不亲和父本自交结实能力可能不同,同一自交不亲和父本在不同的制种环境下也可能会有不同程度的自交结实。目前,利用细胞核雄性不育系RG430A与父本7-5STR在人工大棚隔离放蜜蜂辅助授粉的条件下,父本自交结实所产生种子占整个种子的比列低于1%。
(4)对于父、母本开花期差异特别大,尤其是母本开花期特别早的杂交组合,应用该方法进行杂交F1种子会受到一定限制。当遇到父、母本开花期差异较大的组合时,可以通过人工或机械打苔来使开花期相遇。
[1] 范成明, 田建华, 胡赞民, 王珏, 吕慧颖, 葛毅强, 魏珣, 邓向东, 张蕾颖, 杨维才. 油菜育种行业创新动态与发展趋势. 植物遗传资源学报, 2018, 19(3): 447-454.
Fan C M, Tian J H, Hu Z M, Wang J, Lü H Y, Ge Y Q, Wei X, Deng X D, Zhang L Y, Yang W C.Advances of oilseed rape breeding., 2018, 19(3): 447-454.(in Chinese)
[2] Ogura H. Studies on the new male-sterility in japanese radish, with special reference to the utilization of this sterility towards the practical raising of hybrid seeds.,, 1968, 6(2): 39-78.
[3] Charne D G, Grant I, Kraling K, PATEL J D, PRUVOT J C, TULSIERAM L K. Oilseed Brassica containing an improved fertility restorer gene for ogura cytoplasmic male sterility. 2002, US, US6392127.
[4] Fu T D, Yang G S, Yang X N. Studies on three line Polima cytoplasmic male sterility developed in., 1990, 104: 115-120.
[5] 杨光圣, 傅廷栋. 甘蓝型油菜雄性不育-可育两用系选育成功. 中国农业科学, 1990(1): 90.
Yang G S, Fu T D. Success in the breeding of male sterile and fertile lines inL.., 1990(1): 90. (in Chinese)
[6] 杨光圣, 傅廷栋, 杨小牛, 马朝芝. 甘蓝型油菜生态雄性不育两用系的研究: I. 雄性不育两用系的遗传. 作物学报, 1995, 21(2): 129-135.
Yang G S, Fu T D, Yang X N, ma c z. Studies on the ecotypical male sterile line ofL.: I. Heritance of the ecotypical male sterile line.,1995, 21(2): 129-135. (in Chinese)
[7] 杨光圣, 傅廷栋, 马朝芝, 杨小牛. 甘蓝型油菜生态雄性不育两用系的研究: II. 环境条件对雄性不育两用系的影响. 华中农业大学学报, 1997, 16(5): 330-334.
Yang G S, Fu T D, Ma C Z, Yang X N. Studies on the ecotypical male sterile line ofL.: II. The influence of environment on the ecotypical male sterility., 1997, 16(5): 330-334. (in Chinese)
[8] 袁美, 杨光圣, 傅廷栋, 严红艳. 甘蓝型油菜生态型细胞质雄性不育两用系的研究: III. 8-8112AB的温度敏感性及其遗传. 作物学报, 2003, 29(3): 330-335.
Yuan M, YANG G S, FU T D, yan h y. Studies on the ecotypical male sterile-fertile line ofL.: III. Sensitivity to temperature of 8-8112ab and its inheritance., 2003, 29(3): 330-335. (in Chinese)
[9] Liu Z, Yang Z H, Wang X, Li K D, An H, Liu J, Yang G S, Fu T D, Yi B, Hong D F. A mitochondria-targeted PPR protein restores pol cytoplasmic male sterility by reducing orf224 transcript levels in oilseed rape., 2016, 9(7): 1082-1084.
[10] Yi B, Zeng F Q, Lei S L, Chen Y, Yao X Q, Zhu Y, Wen J, Shen J X, Ma C Z, Tu J X, Fu T D. Two duplicate CYP704B1- homologous genes BnMs1 and BnMs2 are required for pollen exine formation and tapetal development in., 2010, 63(6): 925-938.
[11] 陈凤祥, 胡宝成, 李强生, 张曼琳. 甘蓝型油菜细胞核不育材料9012A的发现与初步研究. 北京农业大学学报, 1993, 19(增刊): 57-61.
Chen F X, Hu B C, Li Q S, Zhang M L. Discovery and study of genic male sterility (GMS) material 9012A inL.., 1993, 19(Suppl.): 57-61. (in Chinese)
[12] 李树林, 钱玉秀, 吴志华. 甘蓝型油菜细胞核雄性不育性的遗传规律探讨及其应用. 上海农业学报, 1985, 1(2): 1-12.
Li S L, Qian Y X, Wu Z H. Inheritance of genic male sterility inand its application to commercial production., 1985, 1(2): 1-12. (in Chinese)
[13] Mariani C, De Beuckeleer M, Truettner J, Leemans J, Goldberg R B. Induction of male sterility in plants by a chimaeric ribonuclease gene., 1990, 347(6295): 737.
[14] 陈凤祥, 胡宝成, 李成, 李强生, 陈维生, 张曼琳. 甘蓝型油菜细胞核雄性不育性的遗传研究: I隐性核不育系9012A的遗传. 作物学报, 1998, 24(4): 431-438.
Chen F X, Hu B C, Li C, Li Q S, Chen W S, Zhang M L. Genetic studies on GMS inL.: I. Inheritance of recessive GMS line 9012A., 1998, 24: 431-438. (in Chinese)
[15] 董发明, 洪登峰, 刘平武, 谢彦周, 何庆彪, 杨光圣. 甘蓝型油菜隐性细胞核雄性不育系9012AB遗传模式新释. 华中农业大学学报, 2010, 29(3): 262-267.
Dong F M, Hong D F, Liu P W, Xie Y Z, He Q B, Yang G S. A novel genetic model for the recessive genic male sterility line 9012AB in rapeseed (L.)., 2010, 29(3): 262-267. (in Chinese)
[16] Dong F M, Hong D F, Xie Y Z, Wen Y P, Dong L, Liu P W, He Q B, Yang G S. Molecular validation of a multiple-allele recessive genic male sterility locus () inL.., 2012, 30(2): 1193-1205.
[17] Deng Z H, Li X, Wang Z Z, Jiang Y F, Wan L L, Dong F M, Chen F X, Hong D F, Yang G S. Map-based cloning reveals the complex organization of the BnRf locus and leads to the identification of, a male sterility gene, in., 2016, 129(1): 53-64.
[18] Dun X L, Zhou Z F, Xia S Q, Wen J, Yi B, Shen J X, Ma C Z, Tu J X, Fu T D. BnaC. Tic40, a plastid inner membrane translocon originating from Brassica oleracea, is essential for tapetal function and microspore development in., 2011, 68(3): 532-545.
[19] Li J, Hong D F, He J P, Ma L, Wan L L, Liu P W, Yang G S. Map-based cloning of a recessive genic male sterility locus inL. and development of its functional marker., 2012, 125(2): 223-234.
[20] Xia S Q, Wang Z X, Zhang H Y, Hu K N, Zhang Z Q, Qin M M, Dun X L, Yi B, Wen J, Ma C Z, Shen J X, Fu T D, Tu J X. Altered transcription and neofunctionalization of duplicated genes rescue the harmful effects of a chimeric gene in., 2016, 28(9): 2060-2078.
[21] Zhao L, Jing X, Chen L, Liu Y J, Su Y N, Liu T T, Gao C B, Yi B, Wen J, Ma C Z, Tu J X, Zou J T, Fu T D, Shen J X. Tribenuron-Methyl induces male sterility through anther-specific inhibition of acetolactate synthase leading to autophagic cell death., 2015, 8(12): 1710-1724.
[22] Zhao L, Deng L, Zhang Q, Jing X, Ma M, Yi B, Wen J, Ma C Z, Tu J X, Fu T D, Shen J X. Autophagy contributes to sulfonylurea herbicide tolerance via GCN2-independent regulation of amino acid homeostasis., 2018, 14(4): 702-714.
[23] Kitashiba H, Nasrallah J B. Self-incompatibility in Brassicaceae crops: lessons for interspecific incompatibility., 2014, 64(1): 23-37.
[24] Nasrallah J B, Nasrallah M E. Pollen-stigma signaling in the sporophytic self-incompatibility response., 1993, 5(10): 1325-1335.
[25] 马朝芝, 傅廷栋, 杨光圣, 涂金星, 杨小牛, 但芳. 甘蓝型油菜双低自交不亲和系的选育. 华中农业大学学报, 1998, 17(3): 211-213.
Ma C Z, Fu T D, Yang G S, Tu J X, Yang X N, Dan F. Breeding for self-incompatibility lines with double zero onL.., 1998, 17(3): 211-213. (in Chinese)
[26] Zhai W, Zhang J F, Yang Y, Ma C Z, Liu Z Q, Gao C B, Zhou G L, Tu J X, Shen J X, Fu T D. Gene expression and genetic analysis reveal diverse causes of recessive self-compatibility inL.., 2014, 15(1): 1037.
[27] Gao C B, Ma C Z, Zhang X G, Li F P, Zhang J F, Zhai W, Wang Y Y, Tu J X, Shen J X, Fu T D. The genetic characterization of self-incompatibility in aline with promising breeding potential., 2013, 31(2): 485-493.
[28] Gao C B, Zhou G L, Ma C Z, Zhai W, Zhang T, Liu Z Q, Yang Y, Wu M, Yue Y, Duan Z Q, Li Y Y, Li B, Li J J, Shen J X, Tu J X, Fu T D. Helitron-like transposons contributed to the mating system transition from out-crossing to self-fertilizing in polyploidL.., 2016, 6(5): 337-355.
[29] Tang J Y, Zhang J F, Ma C Z, Tang W, Gao C B, Li F P, Wang X, Liu Y, Fu T D. CAPS and SCAR markers linked to maintenance of self-incompatibility developed from SP11 inL.., 2009, 24(3): 245-254.
[30] Zhang X G, Ma C Z, Fu T D, Li Y Y, Wang T H, Chen Q, Tu J X, Shen J X. Development of SCAR markers linked to self-incompatibility inL.., 2007, 21(3): 305-315.
[31] Zhang X G, Ma C Z, Tang J Y, Tang W, Tu J X, Shen J X, Fu T D. Distribution of S haplotypes and its relationship with restorer-maintainers of self-incompatibility in cultivated., 2008, 117(2): 171-179.
A simplified production method of hybrid F1seeds in rapeseed
YANG GuangSheng, XIN Qiang, DONG FaMing, HONG DengFeng
(National Key Laboratory of Crop Genetic Improvement, Huazhong Agricultural University, Wuhan 430070)
Safe and effective F1seed production method is the key to hybrid production in rapeseed. Father parent lines with tribenuron-methyl resistance (or tolerance) and self-incompatiblity can be developed by means of backcross breeding. In the F1hybrid seed production by using tribenuron-methyl as the male sterility induction agent, the father parent lines have no seedset by self-pollination because of their self-incompatibility, and are not affected by the chemical agent because of their tribenuron-methyl resistance (or tolerance). Therefore, the parental lines can be mixed sowing and the seeds can be mixed harvesting. Using the method we proposed, if the lines with tribenuron-methyl resistance (tolerance) and self-incompatiblity are already developed, hybrid breeding can be carried out immediately. It saves labor costs, improves seed production efficiency and increases seed purity because of the mixed sowing and mixed harvesting.
; hybrid; F1seed production; chemical inducing male sterility; self-incompatibility
10.3864/j.issn.0578-1752.2019.08.004
2018-11-27;
2019-01-20
油菜杂种优势利用技术与强优势杂交种创制(2016YFD0101300)、油菜温敏细胞质雄性不育机理与应用研究(2662017PY020)
杨光圣,Tel:027-87281713;E-mail:gsyang@mail.hzau.edu.cn
(责任编辑 李莉)
