外源氮肥和腐熟剂对小麦秸秆腐解的影响
2019-04-29朱远芃金梦灿马超广敏高敏郜红建
朱远芃,金梦灿,马超,广敏,高敏,郜红建
农田生态保育与污染防控安徽省重点实验室/安徽农业大学资源与环境学院,安徽 合肥 230036
中国秸秆资源可收集量约9.0亿吨,所含的氮(N)、磷(P2O5)、钾(K2O)养分总量分别达到 625.6、197.9、1159.5万吨(宋大利等,2018)。秸秆还田作为培肥土壤的有效方式,可改善土壤物理结构、增加养分储量,提高土壤有机质含量和土壤微生物活性。由于还田秸秆腐解速率低、腐解微生物与作物争氮以及作物病虫害加重等负面效应而引起广泛关注(马骁轩等,2016;丁雪丽等,2008;王爱玲等,2003)。
秸秆腐解过程是影响养分循环和作物利用的主要因素,其腐解速率受秸秆本身的化学组成、温度、通气和水分等外界环境的制约。不同种类的植物秸秆、同种植物不同组织器官或不同株龄,其化学组成存在显著差异,降解速率也不同(Kabuyah et al.,2012)。秸秆腐解过程受秸秆含氮量、C/N和木质素含量等因素的影响(Jing et al.,2016;Li et al.,2017)。秸秆含氮量越高,木质素含量越低,或者C/N和木质素/N越低,越有利于秸秆腐解(Dannehl et al.,2017)。添加适量的无机氮素有利于秸秆腐解(Shahzad et al.,2015),但高浓度的无机氮素会抑制微生物活性,进而影响秸秆腐解(Li et al.,2017)。氮素主要抑制真菌生长,而对细菌影响不显著(Nicolardot et al.,2001;Wang et al.,2003)。研究表明,微生物分解有机物的最适C/N为25∶1,而禾本科作物秸秆的C/N一般都高于最适值,例如小麦茎秆 C/N 可达到 86∶1(Kochsiek et al.,2013)。微生物在腐解秸秆过程中为维持正常的C/N需从外界吸收氮,发生秸秆腐解微生物与植株“争氮”等现象,因此,补充适量氮素可促进秸秆腐解(李昌明等,2017)。比如通过增施外源氮肥,促进了小麦、玉米等秸秆的腐解(高金虎等,2012;黄婷苗等,2017;张经廷等,2018)。外源增氮可提高纤维素酶等水解酶活性,抑制氧化酶活性,进而影响作物秸秆和凋落物等物质腐解(Keeler et al.,2009;Riggs,2016;Zang et al.,2016)。
秸秆腐熟剂是一种由微生物组成、能够快速降解秸秆的微生物菌剂,利用真菌、细菌和放线菌的分解代谢作用将秸秆中的纤维素、半纤维素和木质素分解转化为小分子有机化合物或者产生CO2释放出来。微生物通过分泌胞外酶促进秸秆的矿化分解(Geisseler et al.,2008),其中水解酶和氧化酶在秸秆腐解过程中起重要作用。前人研究多关注不同腐熟剂种类及其配伍对作物秸秆腐解的影响(胡诚等,2016;丁文成等,2016;陈帅等,2016),也有研究关注不同通气、水分、温度等环境条件下腐解剂对秸秆腐解的影响(Zhang et al.,2016;Sandeep et al.,2016)。有关氮素对秸秆腐解影响的研究多集中在氮素形态转化及其对作物氮素有效性等方面,但有关氮素和腐解剂组合对作物秸秆腐解的协同作用机理还缺乏深入研究。
本文采用田间堆腐的方法,研究氮素和腐熟剂组合对小麦秸秆腐解的协同作用,分析小麦秸秆堆腐过程中纤维素酶等水解酶和木质素过氧化物酶等氧化酶活性的变化规律;利用偏最小二乘回归法,分析小麦秸秆腐解速率与小麦秸秆腐解酶活性变化之间的耦合关系,为还田秸秆快速腐解提供理论依据。
1 材料与方法
1.1 试验材料
供试地点:试验地位于安徽省庐江县郭河镇安徽农业大学皖中综合试验站(31°25′N,117°09′E),试验地土壤类型为黄棕壤发育而来的水稻土,土壤pH 5.30、全氮 1.22 g·kg-1、碱解氮 87.55 mg·kg-1、速效磷 12.92 mg·kg-1、速效钾 266.01 mg·kg-1。
供试秸秆,为冬小麦秸秆,全碳含量40.42%、全氮0.82%、全磷0.10%、全钾1.30%、C/N 49.29。氮肥为尿素(46%),购自上海苏懿化学试剂有限公司;秸秆腐解剂由中国农业科学院农业区划研究所研制的有机物料腐熟菌剂(除臭型),有效活菌数0.5×108ind·g-1。
1.2 试验设计与样品采集
秸秆堆腐试验于2017年6-10月在供试地点进行。将收获小麦秸秆烘干后剪成3 cm,取20.0 g放入 100目尼龙网袋,尼龙网袋规格为 30 cm×50 cm,扎口后平摊放置在秸秆条垛中(条垛长3 m、宽2 m、高1 m),条垛四周开有排水沟,接受自然条件下的日晒雨淋。
试验共设置:(1)秸秆自然堆腐(Straw composting,S);(2)秸秆氮肥堆腐(Nitrogen plus straw composting,NS);(3)腐解剂堆腐(Effective microorganisms plus straw composting,ES);(4)氮肥腐解剂堆腐(Nitrogen and effective microorganisms plus straw composting,NESS)等4个处理,每个处理设置3个重复,共埋60个尼龙网袋,每个处理埋12个网袋分5个时间段无放回采集。NS处理添加0.20 g尿素;ES处理添加0.20 g腐熟剂;NESS处理添加0.20 g尿素和0.20 g腐熟剂(李庆康等,2001;潘剑玲等,2013)。在小麦秸秆堆腐试验开始后的7、15、30、70和120 d采集堆腐秸秆,每个小区(5 m×10 m)随机取3个尼龙网袋,装入无菌自封袋中并迅速带回实验室,将样品分成两部分,一部分秸秆烘干称重用于理化性质分析,另一部分秸秆保存在4 ℃冰箱,用于酶活性测定。
1.3 堆腐过程质量损失测定
每次取样时随机从秸秆条垛堆中选取 3个网袋,用水冲洗网袋粘附的泥浆,在75 ℃下烘干(鲍士旦,2002),称重,计算秸秆质量残留率:

式中,Rsr为秸秆残留率;M0为秸秆原始干重(g);Mt为腐解t d后的秸秆干重(g);t为腐解时间(d)。小麦秸秆腐解规律参照 Olson(1963)的理论,利用一阶指数衰退模型进行拟合:

式中,W0为初始秸秆质量(g);Wt为腐解t d后秸秆残余质量(g);k为腐解常数;t为腐解时间(d)。
1.4 酶活性测定方法
1.4.1 β-葡萄糖苷酶(β-Glucosidase,GC)活性的测定方法
β-葡萄糖苷酶活性采用DNS法测定(Martins et al.,2008),利用 β-葡萄糖苷酶分解对-硝基苯-β-D-吡喃葡萄糖苷生成对-硝基苯酚,后者在400 nm有最大吸收峰,通过测定吸光值来计算β-葡萄糖苷酶活性。酶活定义:每毫克组织蛋白每分钟产生1 nmol对硝基苯酚所需的酶量定义为1个酶活性单位,即为 nmol·min-1·mg-1。
1.4.2 秸秆纤维素酶(Cellulase,CL)活性的测定方法
纤维素酶活性采用蒽酮比色法(Martins et al.,2008)测定。分析纤维素酶催化羧甲基纤维素钠降解产生的还原糖的含量。酶活的定义:每毫克组织蛋白每分钟催化产生 1 μg葡萄糖所需的酶量定义为 1 个酶活力单位,即为 μg·min-1·mg-1。
1.4.3 秸秆中性木聚糖酶(Neutral Xylanase,NEX)活性的测定方法
木聚糖酶活性采用DNS法测定(Yang et al.,2015)。中性木聚糖酶在中性环境中催化木聚糖降解成还原性寡糖和单糖,在沸水浴条件下进一步与3, 5-二硝基水杨酸发生显色反应,在540 nm处有特征吸收峰,反应液颜色的深浅与酶解产生的还原糖量成正比,通过测定反应液在540 nm吸光值增加速率,可计算中性木聚糖酶活力。酶活定义:50 ℃,pH 6.0条件下,每毫克蛋白每分钟分解木聚糖产生1 nmol还原糖所需的酶量为1个中性木聚糖酶的活力单位,即为 nmol·min-1·mg-1。
1.4.4 秸秆木质素过氧化物酶(Lignin peroxidase,Lip)活性的测定方法
木质素过氧化物酶活性采用藜芦醇氧化速率法(Yadav et al.,2012)测定。木质素过氧化物酶氧化藜芦醇生成藜芦醛,在310 nm处有特征吸收峰。酶活定义:每毫克蛋白每分钟氧化1 nmol藜芦醇所需的酶量为 1个酶活力单位(U),即为nmol·min-1·mg-1。
1.4.5 秸秆锰过氧化物酶(Manganese peroxidase,Mnp)活性的测定方法
锰过氧化物酶活性采用愈创木酚氧化速率法(Anderson et al.,2010)测定。锰过氧化物酶在Mn2+存在的条件下,将愈创木酚氧化成四邻甲氧基连酚,在465 nm有特征吸收峰。酶活定义:每毫克蛋白每分钟氧化1 nmol愈创木酚所需的酶量为1个酶活力单位,即为 nmol·min-1·mg-1。
1.4.6 秸秆漆酶(Laccase,LA)活性的测定方法
漆酶活性采用ABTS 法(Ling et al.,2015)测定。漆酶分解底物ABTS产生ABTS自由基,在420 nm 处的吸光系数远大于底物 ABTS,测定 ABTS自由基的增加速率,可计算得漆酶活性。酶活定义:每毫克蛋白每分钟氧化1 nmol底物ABTS所需的酶量为 1 个酶活力单位,即为 nmol·min-1·mg-1。
1.5 数据处理
数据的多重比较采用Duncan法分析(P<0.05),差异显著性检验(ANOVA)采用SPSS 19.0软件分析。质量损失与多个酶活性的相关性采用偏最小二乘法(PLS)。
2 结果与分析
2.1 不同处理下小麦秸秆的质量变化
小麦秸秆在不同处理条件下的质量残留量均随培养时间延长呈逐渐降低趋势,且均表现为前期下降快,后期下降慢的规律(图1)。在试验开始的前30 d内,NS、ES和NESS处理的小麦秸秆质量迅速降低,与 S处理相比达到显著性差异水平(P<0.05)。培养至70 d时,NS、ES和NESS处理的小麦秸秆质量残留率分别为 29.84%、32.04%和25.01%,分别比 S处理多降解了3.95%、1.76%和8.78%。培养至120 d时,小麦秸秆在NS、ES和NESS处理的质量残留率分别为25.30%、26.74%和20.84%,即在培养的70-120 d内,ES处理的小麦秸秆腐解快于 NS处理,NESS处理的小麦秸秆腐解率高于NS和ES处理。
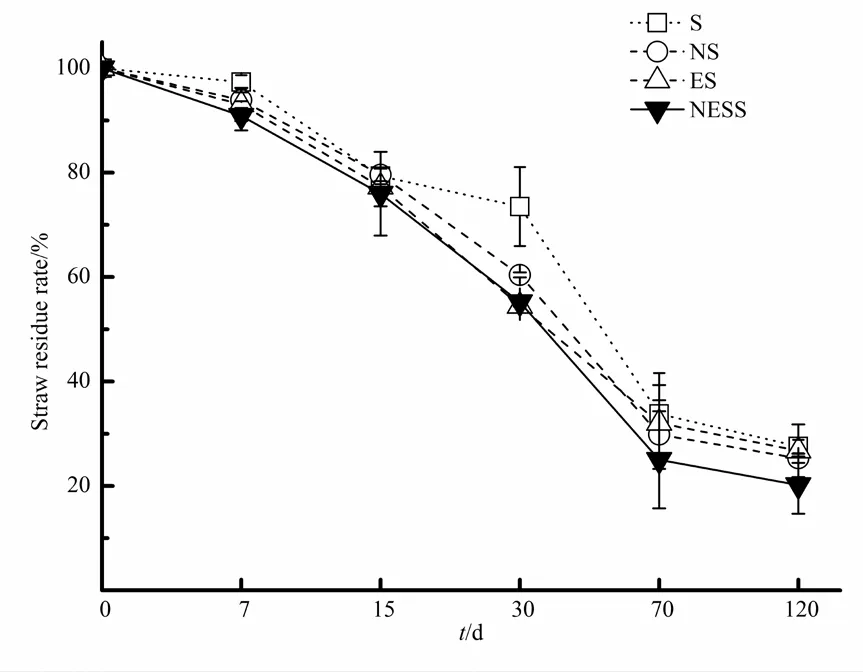
图1 小麦秸秆质量变化规律Fig. 1 Mass change regularity of wheat straw

表1 不同处理条件下小麦秸秆质量残留与堆腐时间关系的拟合Table 1 Regression models of the wheat straw residual quantity and composting time under different treatments
小麦秸秆残留质量随时间变化可用一阶指数衰退模型[Wt=W0×exp(-kt)]进行拟合(表 1),相关性较高(R2>0.80)。堆腐结束时(120 d),NS、ES和 NESS 3种处理条件下秸秆腐解率分别为74.70%、73.26%和 79.83%;腐解速率常数k分别为 14×10-3·d-1、14×10-3·d-1和 17×10-3·d-1。这说明氮肥和腐熟剂同时添加更有利于小麦秸秆腐解(P<0.05)。
2.2 小麦秸秆腐解过程中胞外酶活性的变化
如图2所示,β-葡萄糖苷酶活性在不同处理中均随培养时间延长呈先增加而后降低的趋势。β-葡萄糖苷酶活性在纯秸秆处理中(S)以培养至30 d时活性最高,为15.50 nmol·min-1·mg-1;在 NS 处理中以培养至 7 d时活性最高,为 20.44 nmol·min-1·mg-1;在 ES 处理中以培养至 70d 时活性最高,为 29.69 nmol·min-1·mg-1,分别比 S 和 NS处理时的酶活性提高65.59%和63.45%。NESS处理的β-葡萄糖苷酶活性以培养至70 d时活性最高,为23.49 nmol·min-1·mg-1,分别比 S 和 NS 处理时的酶活性提高56.51%和53.80%,比ES下降了20.88%。培养至120 d时,β-葡萄糖苷酶活性以NS处理的最高,达 14.32 nmol·min-1·mg-1,分别比 S、ES 和 NESS高31.96%、22.17%和51.10%。
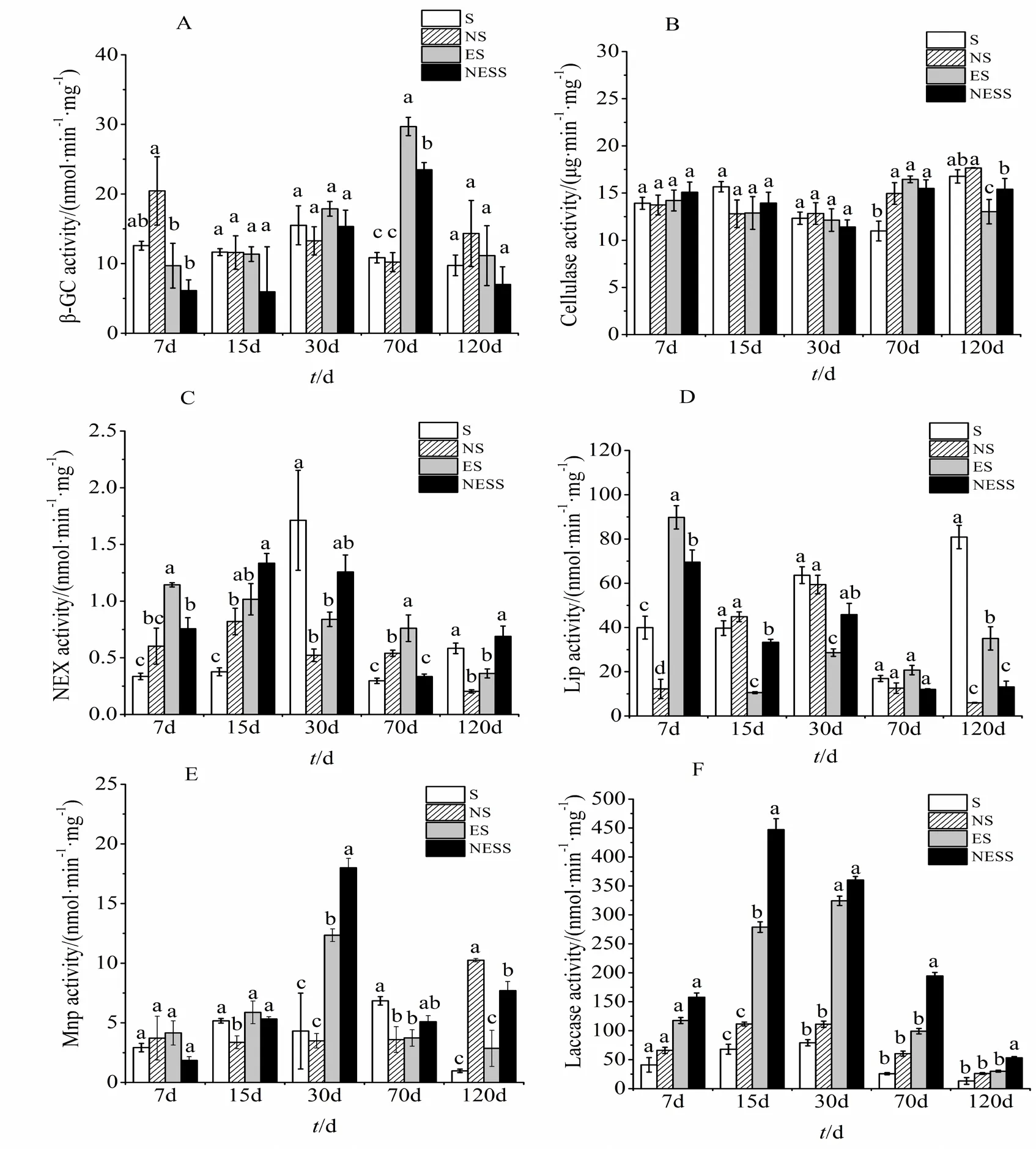
图2 小麦腐解过程中酶活性的变化Fig. 2 Changes of enzyme activity during decomposing
纤维素酶活性在不同处理中均随培养时间延长呈现先降低后升高的趋势。纤维素酶活性在纯秸秆处理中(S)以培养至120 d时活性最高,为16.77 μg·min-1·mg-1;在 NS 处理中以培养至 120 d 时活性最高,为 17.66 μg·min-1·mg-1;在 ES 处理中以培养至 70 d 时活性最高,为 16.46 μg·min-1·mg-1,分别比S和NS处理时的酶活性提高9.12%和33.30%。NESS处理的纤维素酶活性以培养至70 d时活性最高,为 15.49 μg·min-1·mg-1,分别比 S 和 NS 处理时的酶活性提高 3.45%和 29.13%;比 ES下降了5.88%。培养至120 d时,纤维素酶活性以NS处理的最高,达 17.66 μg·min-1·mg-1,分别比 S、ES 和NESS高5.06%、26.23%和12.84%。
中性木聚糖酶活性在不同处理中均随培养时间延长呈现先升高后降低的趋势。中性木聚糖酶活性在纯秸秆处理中(S)以培养至30 d时活性最高,为 1.71 nmol·min-1·mg-1;在 NS 处理中以培养至 15 d 时活性最高,为 0.82 nmol·min-1·mg-1;在 ES 处理中以培养至 7 d时活性最高,为 1.14 nmol·min-1·mg-1,分别比 S 和 NS 处理时的酶活性提高70.62%和47.30%。NESS处理的中性木聚糖酶活性以培养至 15 d时活性最高,为 1.33 nmol·min-1·mg-1;分别比 S、NS 和 ES 处理时的酶活性提高71.80%、38.55%和23.87%。培养至120 d时,中性木聚糖酶活性以NESS处理最高,达0.69 nmol·min-1·mg-1,分别比 S、NS 和 ES 高 15.33%、70.39%和47.42%。
木质素过氧化物酶活性在不同处理中均随培养时间延长呈现逐渐降低的趋势。木质素过氧化物酶活性在纯秸秆处理中(S)以培养至120 d时活性最高,为 80.88 nmol·min-1·mg-1;在 NS 处理中以培养至 30 d 时活性最高,为 59.39 nmol·min-1·mg-1;在 ES处理中以培养至 7 d时活性最高,为 89.76 nmol·min-1·mg-1,分别比 S 和 NS 处理时的酶活性提高55.50%和86.38%。NESS处理的木质素过氧化物酶活性以培养至 7 d时活性最高,为 69.52 nmol·min-1·mg-1;分别比 S 和 NS 处理时的酶活性提高42.55%和82.41%;比ES下降了22.55%。培养至120 d时,木质素过氧化物酶活性以S处理的最高,达 80.88 nmol·min-1·mg-1,分别比 NS、ES和NESS高92.64%、56.68%和83.76%。
锰过氧化物酶活性在不同处理中均随培养时间延长呈现先升高后降低的趋势。锰过氧化物酶活性在纯秸秆处理中(S)以培养至70 d时活性最高,为 6.85 nmol·min-1·mg-1;在 NS 处理中以培养至 120 d 时活性最高,为 10.25 nmol·min-1·mg-1;在 ES 处理中以培养至 30 d时活性最高,为 12.34 nmol·min-1·mg-1;分别比 S 和 NS 处理时酶活性提高65.09%和71.72%。NESS处理的锰过氧化物酶活性以培养至 30 d活性最高,为 17.99 nmol·min-1·mg-1;分别比 S、NS 和 ES 处理时的酶活性提高76.05%、80.60%和31.40%。培养至120 d时,锰过氧化物酶活性以NS处理的最高,达10.25 nmol·min-1·mg-1,分别比 S、ES 和 NESS 高 90.50%、72.13%和24.93%。
漆酶活性在不同处理中均随培养时间延长呈现先升高后降低的趋势。漆酶活性在纯秸秆处理中(S)以培养至 30 d时活性最高,为 79.12 nmol·min-1·mg-1;在 NS 处理中以培养至 15 d 时活性最高,为 111.53 nmol·min-1·mg-1;在 ES 处理中以培养至30 d时活性最高,为 324.28 nmol·min-1·mg-1;分别比 S 和 NS 处理时酶活性提高75.60%和65.70%。NESS处理的漆酶活性以培养至 15 d 活性最高,为 447.33 nmol·min-1·mg-1;分别比S、NS和ES处理时的酶活性提高84.80%;75.07%和37.65%。培养至120 d时,漆酶活性以NESS处理的最高,达 53.15 nmol·min-1·mg-1,分别比 S、NS和ES高75.15%、50.81%和43.41%。
2.3 胞外酶活性与秸秆生物量损失的相关性及偏最小二乘回归分析
小麦秸秆质量损失量和胞外酶活性之间的偏最小二乘法(Partial Least Squares,PLS)回归分析结果显示(表2),不同处理中胞外酶活性对秸秆腐解作用差异较大。在S处理条件下(R2=0.462),纤维素酶(CL)、锰过氧化物酶(Mnp)和漆酶(LA)活性与小麦秸秆腐解速率(质量损失量)呈正相关,回归系数分别为0.39、0.43和0.17;而β-葡萄糖苷酶(β-GC)、中性木聚糖酶(NEX)和木质素过氧化物酶(LiP)活性与小麦秸秆腐解速率呈负相关,其回归系数分别为-0.22、-0.98和-0.34。在NS处理条件下,仅有CL活性与小麦秸秆腐解速率(质量损失量)呈正相关,回归系数为 0.016;其余酶活性均与小麦秸秆腐解速率呈负相关,回归系数在-0.033 (MnP)--0.457 (NEX)之间。在ES处理条件下,仅有CL和MnP活性与小麦秸秆腐解速率(质量损失量)呈正相关,回归系数分别为 0.327和0.085;其余酶活性均与小麦秸秆腐解速率呈负相关,回归系数在-0.048 (β-GC)--0.764 (LiP)之间。在NESS处理条件下,CL和MnP活性与小麦秸秆腐解速率(质量损失量)呈正相关,回归系数分别为1.945和2.096;其余酶活性均与小麦秸秆腐解速率呈负相关,回归系数在-0.365 (β-GC)-2.529(LA)之间。
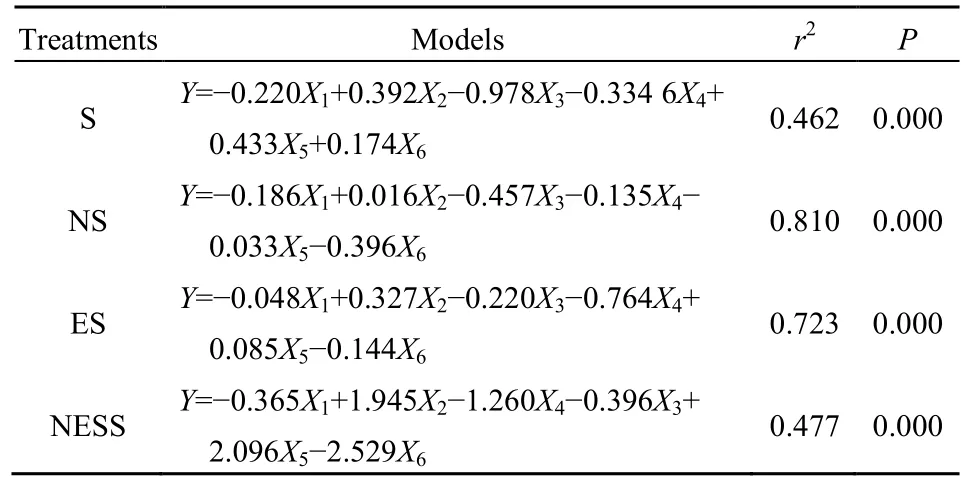
表2 不同处理质量损失量与酶活性的偏最小二乘法回归分析Table 2 Partial least square regression analysis of quality loss and enzyme activity in different treatments
综上所述,在S处理条件下,纤维素酶,锰过氧化物酶和漆酶对秸秆腐解的影响较大,且氧化酶(Mnp,LA)回归系数之和大于水解酶(CL),说明自然堆腐条件下氧化酶比水解酶更能促进秸秆腐解。在NS处理条件下,纤维素酶对秸秆腐解的影响较大,说明氮肥可通过提高纤维素酶活性进而促进秸秆腐解。在ES处理条件下,纤维素酶和锰过氧化物酶对秸秆腐解的影响较大,且水解酶(CL)回归系数大于氧化酶(Mnp),说明腐熟剂主要通过提高水解酶活性促进秸秆腐解。在NESS处理条件下,纤维素和锰过氧化物酶对秸秆腐解的影响较大,且氧化酶(Mnp)回归系数大于水解酶(CL),说明氮肥和腐熟剂混合通过提高氧化酶活性促进秸秆腐解。
3 讨论
3.1 外源微生物菌剂对麦秸腐解的影响
腐熟剂是含有大量的细菌、霉菌、酵母菌和芽孢秆菌等的高效微生物制剂。研究表明,添加秸秆腐熟剂可以促进作物秸秆腐解,缩短腐解转化时间(蔡立群等,2014;耿丽平等,2015)。本试验结果表明,添加腐熟剂显著提升了小麦秸秆腐解速率,这可能与添加腐熟剂后中性木聚糖酶、木质素过氧化物酶和漆酶活性(P<0.05)提高有关。PLS结果表明,纤维素酶和锰过氧化物酶对秸秆腐解的影响较大。这可能是因为腐熟剂中的真菌是降解植物细胞壁的分解者,通过分泌纤维素酶等水解酶和锰过氧化物酶等氧化酶使植株凋落物的微孔结构纤维化(Arora et al.,2002;Wiedermann et al.,2017),从而加速秸秆腐解。纤维素酶包含β-葡萄糖苷酶,木聚糖酶等多种水解酶,通过加快秸秆中多糖以及可溶性碳等组分的分解,可提高秸秆腐解速率(Wiedermann et al.,2017);而锰过氧化物酶可以将Mn2+氧化成更活跃的 Mn3+,Mn3+可以穿透秸秆纤维素内的木质素结构,氧化木质素苯环上的酚醛基团,从而加速木质素的腐解(Wiedermann et al.,2017)。
3.2 外源氮素对麦秸腐解的影响
在小麦秸秆堆腐的120 d内,氮肥能显著促进小麦秸秆腐解,但腐解后期速率减慢(图1),这可能是因为在秸秆腐解前期外源氮肥可弥补秸秆固有氮素的不足,氮肥为微生物提供生长所需要的氮元素,促进微生物生长并分泌秸秆β-葡萄糖苷酶和木聚糖酶,加速了秸秆体内纤维素多糖等可溶性组分的降解。陆地生态系统中,在低氮环境下,微生物在产生胞外酶的过程中需要获得更多的能量来弥补氮的不足(Rinkes et al.,2016),适量的外源氮可以提高胞外酶活性,从而加快植物凋落物腐解。在秸秆腐解后期,可溶性糖等易降解组分相对含量下降,而木质素等难降解组分相对含量升高,且氮素可与秸秆残余物中木质素苯环上的酚羟基结合,形成更加稳定的苯胺类物质,从而减缓了秸秆腐解后期的腐解速率(Fog,2010)。
PLS结果显示,氮肥和腐熟剂同时处理时氧化酶活性与质量损失的相关性也高于腐熟剂单独处理,说明氮肥和腐熟剂具有共同加速小麦秸秆腐解的协同效应。
4 结论
(1)外源氮肥或腐熟剂均能显著加速小麦秸秆腐解(质量损失加快),氮肥和腐熟剂组合可以协同促进小麦秸秆腐解。
(2)外源氮肥主要通过提高水解酶活性加速小麦秸秆腐解,而外源腐熟剂主要通过促进氧化酶活性加速小麦秸秆腐解。
