过表达水稻OsSAPP2基因促进转基因拟南芥叶片衰老
2019-04-27张艺函蒋太英崔胜男马殿荣
张艺函,徐 凡,蒋太英,崔胜男,马殿荣
(沈阳农业大学 水稻研究所/农业部东北水稻生物学与遗传育种重点实验室/北方超级粳稻育种教育部重点实验室,沈阳110161)
衰老是植物叶片发育的最后一个阶段,在这个过程中植物利用叶片的衰老来实现其自身营养的循环再利用,从而保证在发育的不同阶段获得足够的营养来供应生长[1]。植物叶片的衰老受到内部发育信号和外部多种环境因子的协调控制[2]。叶片衰老还与糖、钙离子、活性氧含量及植物激素水平等因素相关;最近研究发现植物激素家族的新成员独脚金内酯(Strigolactone,SL)也通过与乙烯信号转导途径互作而参与叶片衰老进程的调控[3]。各因子与发育信号形成交叉复杂的信号通路网络进而对衰老进程进行调控。在农业生产方面,叶片衰老可以降低作物产量,影响作物品质[4]。据有关报道统计,当植物受非生物胁迫时会加快衰老的进程,因非生物因素导致的农作物早熟早衰可使全球粮食主产区的平均产量减少一半[5]。由蛋白激酶(protein kinase,PK)和蛋白磷酸酶(protein phosphatase,PP)所催化的蛋白质可逆磷酸化反应是细胞信号识别与传导的重要环节[6]。蛋白磷酸酶的结构变化较大,2C型蛋白磷酸酶(protein phosphatase 2C,PP2C)是一种丝氨酸/苏氨酸残基蛋白磷酸酶,是高等植物中数量较多的一类。PP2C编码基因的分布十分广泛,除病毒外几乎所有生物均存在。XUE等[7]依据对PP2C蛋白保守催化结构域的分析,分别在拟南芥(Arabidopsis thaliana)和水稻中发现了80个和78个PP2C编码基因;在其他物种中(如玉米、苜蓿、山毛榉和鸢尾等)也发现了PP2C编码基因,它们大多数参与植物对于非生物胁迫的响应,如拟南芥的AtbZIP19/23、玉米(Zea mays)的ABP9和水稻的OsPP2C51作为ABA信号传导的辅助受体,在干旱、寒冷和盐胁迫的ABA依赖性应激反应中起重要作用[8-10];甜樱桃(Prunusavium)的PacPP2C1协同SnRK2蛋白激酶在果实成熟和改善植物对干旱胁迫的耐受性中起关键作用[11];拟南芥的ABI1、ABI2和山毛榉(Fagus)的FsPP2C1、FsPP2C2参与ABA调控的种子休眠/萌发信号途径等。蛋白磷酸酶也在植物叶片调控衰老进程中发挥作用,利用基因芯片技术对大麦(Hordeum vulgare)叶片发育过程中的3个阶段(初衰老期、衰老中期、衰老晚期)进行转录组分析,发现有11个蛋白磷酸酶编码基因在大麦叶片自然衰老过程转录水平发生变化[12];一个定位于拟南芥的高尔基体PP2C,SAG113在叶片衰老过程中参与ABA介导的气孔运动和水分丧失,而且SAG113基因的表达与拟南芥叶片衰老调控重要转录因子AtNAP基因的表达具有表达协同性[13];参与拟南芥叶片衰老负调控的蛋白磷酸酶SSPP,与叶片衰老正调控的蛋白磷酸酶AtSARK相互作用而参与叶片衰老进程的调控[14-16]。目前,在水稻中关于参与叶片衰老进程调控蛋白磷酸酶基因的克隆、功能分析及其调控机理还鲜有报道。本课题组发现并克隆了一个水稻的PP2C编码基因(LOC_06g50380;Os06g0717800),命名为 OsSAPP2(Oryza sativa Senescence-Associated Protein Phosphatase 2,OsSAPP2),OsSAPP2基因CDS全长1179bp,编码393个氨基酸。构建了35S-OsSAPP2双源表达载体,并获得相应的转基因拟南芥。通过表型观察发现,与野生型拟南芥(WT)相比,过表达OsSAPP2基因导致转基因拟南芥叶片变小、叶片数量减少,抽苔开花提前,叶绿素含量下降,叶片早衰等表型;另外,衰老相关转录因子等标志基因的表达量上升。因此可知,OsSAPP2基因是一个参与植物叶片衰老进程调控的关键基因,是水稻叶片衰老的正调控因子。
1 材料与方法
1.1 材料
设计引物扩增OsSAPP2的CDS序列,在上游引物CDSOsSAPP2-F中引入Nco I的酶切位点,在下游引物CDSOsSAPP2-R中引入BstE II的酶切位点。以水稻cDNA为模板,用高保真聚合酶PCR,克隆产物长度1179bp,然后将PCR产物连入T载体(pGEM-T Easy Vector),完成启动子的TA克隆;将所获得的TA克隆和植物双源表达载体pCAMBIA1301基本载体用Nco I/BstE II双酶切,分别回收小片段和载体片段,连接完成35S-OsSAPP2双源表达载体的构建。
引物序列:OsSAPP2-1 为 5’GGATCCATGATAGTGACATTGATGAACTTGTTACGG 3’;OsSAPP2-2 为 5’GAACTCTCACATTTGAGATGCATAAGGTGCA 3’。
拟南芥的培养条件为:光/暗时长为 16h/8h,温度(21±1)℃,光照强度约 90μmol·m-2·s-1;野生型拟南芥(Arabidopsis thaliana),生态型为 Columbia-0。
1.2 方法
1.2.1 拟南芥表型观察 将纯合的转基因拟南芥种子和野生型拟南芥种子用10%安替福民进行表面消毒5min,用无菌蒸馏水冲洗4次,4℃低温处理2d,然后均匀播种于1/2 MS培养基中,放于培养箱培养7~10d。将长势良好的拟南芥植株移植于培养土中培养。在苗龄23d时统计36株拟南芥莲座叶的数目,在苗龄28d时测量18株拟南芥最大莲座叶的长度(单位mm),在苗龄45d时测量18株拟南芥的株高(单位cm)。生长期间使用相机(Nikon D3200)和扫描仪(EPSON Perfection v33)记录拟南芥表型结果。
1.2.2 叶绿素含量的测定 将需检测的拟南芥第5,6片叶子剪下,立即称重0.5~1.0g,迅速剪碎置于2.0mL EP管中,随后快速向EP管中加入2mL 95%无水乙醇。用锡箔纸包裹好,遮光条件下放置1~2d,浸泡至叶片绿色褪去即可,使用多功能酶标仪(购自基因有限公司)测量其叶绿素含量,数据分析参照李合生(2000)的方法[17]。
1.2.3 标志基因表达检测 采用上海Promega公司的Eastep Super总RNA提取试剂盒提取不同叶龄(从移苗第2天开始计算)的转基因拟南芥和野生型拟南芥第5,6片莲座叶总RNA,提取黑暗处理0,1,8d的拟南芥叶片总RNA。采用大连TaKaRa公司的PrimeScriptTMRT Master Mix试剂盒和SYBR®Premix Ex TaqTM试剂盒将总RNA反转录成cDNA,以TIP41-like作为内参进行实时定量RT-PCR,检测衰老标志基因SAG12,衰老关键转录因子NAC2、NAP、WRKY6,叶绿素降解关键酶编码基因ACD1和光合作用关键基因RbcS和RbcL的表达水平。
1.2.4 人工黑暗处理 将长势良好的拟南芥植株移植于1/2MS培养基上培养18~20d,分别在处理0,1,8d时取第5,6片莲座叶置于含有1/2MS液体培养基滤纸的培养皿,于温度为(21±1)℃的暗培养箱培养。
2 结果与分析
2.1 过表达OsSAPP2基因促进转基因拟南芥叶片发生衰老
完成组成型启动子驱动OsSAPP2基因过表达的双源表达载体35S-OsSAPP2,并获得相应的转基因拟南芥。对OsSAPP2基因在转基因拟南芥中的表达情况进行检测,其中line1、line15和line18株系基因属于目的基因过表达株系,line6为目的基因沉默株系(图1A)。选择line1和line18这2个株系为代表进行后续试验。
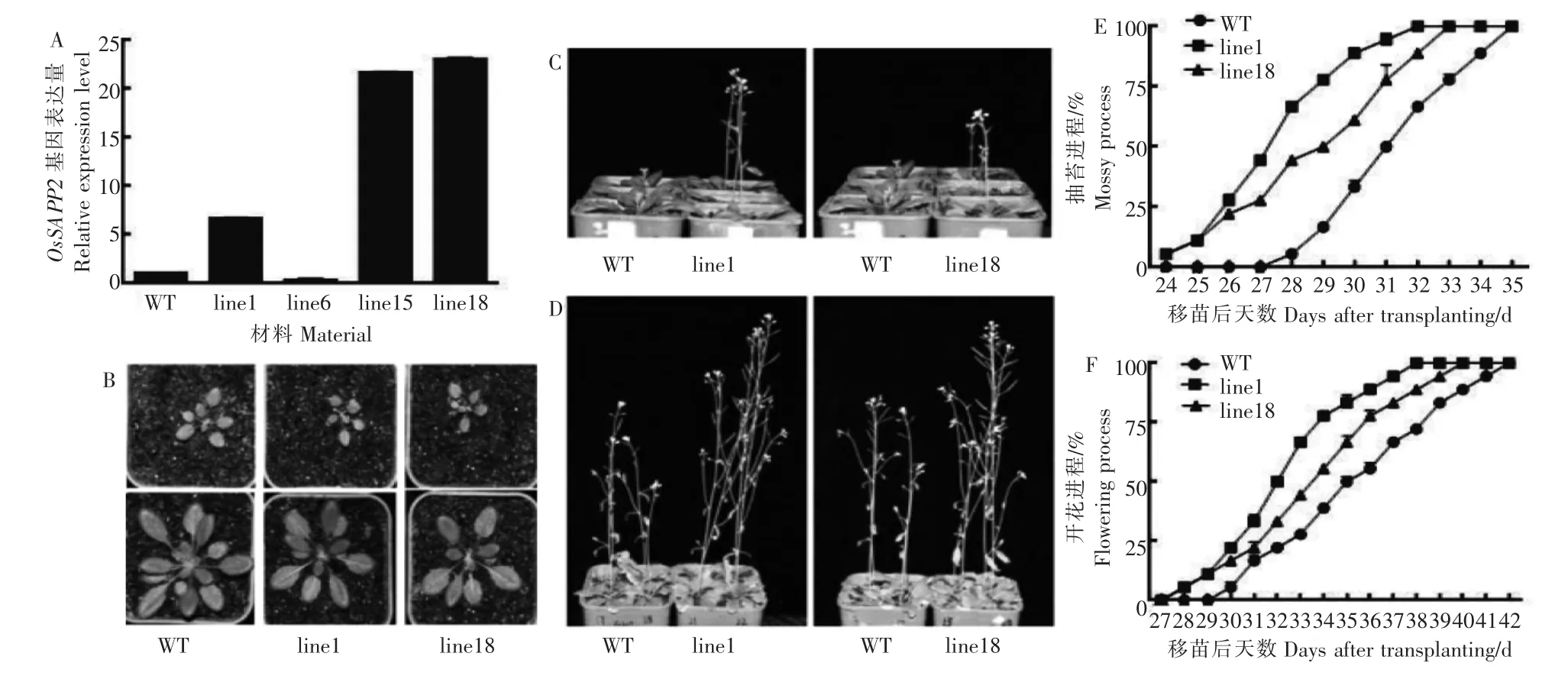
图1 野生型(WT)和35S-OsSAPP2转基因拟南芥植株Figure 1 Growth process of plants in WT and 35S-OsSAPP2 transgenic lines
对野生型拟南芥(WT)和过表达OsSAPP2的转基因拟南芥的莲座叶数目、莲座叶大小、株高、抽苔时间、开花时间等观察发现,过表达OsSAPP2基因可以导致转基因拟南芥叶片变小约15%,莲座叶数目减少2~3片,植株高度增加约20%(表1),统计分析发现差异极显著;同时转基因拟南芥的抽苔和开花时间明显提前(图1)。

表1 35S-OsSAPP2转基因拟南芥株系与野生型拟南芥的表型数据Table 1 The phenotype date of 35S-OsSAPP2 transgenic lines and wild-type Arabidopisis
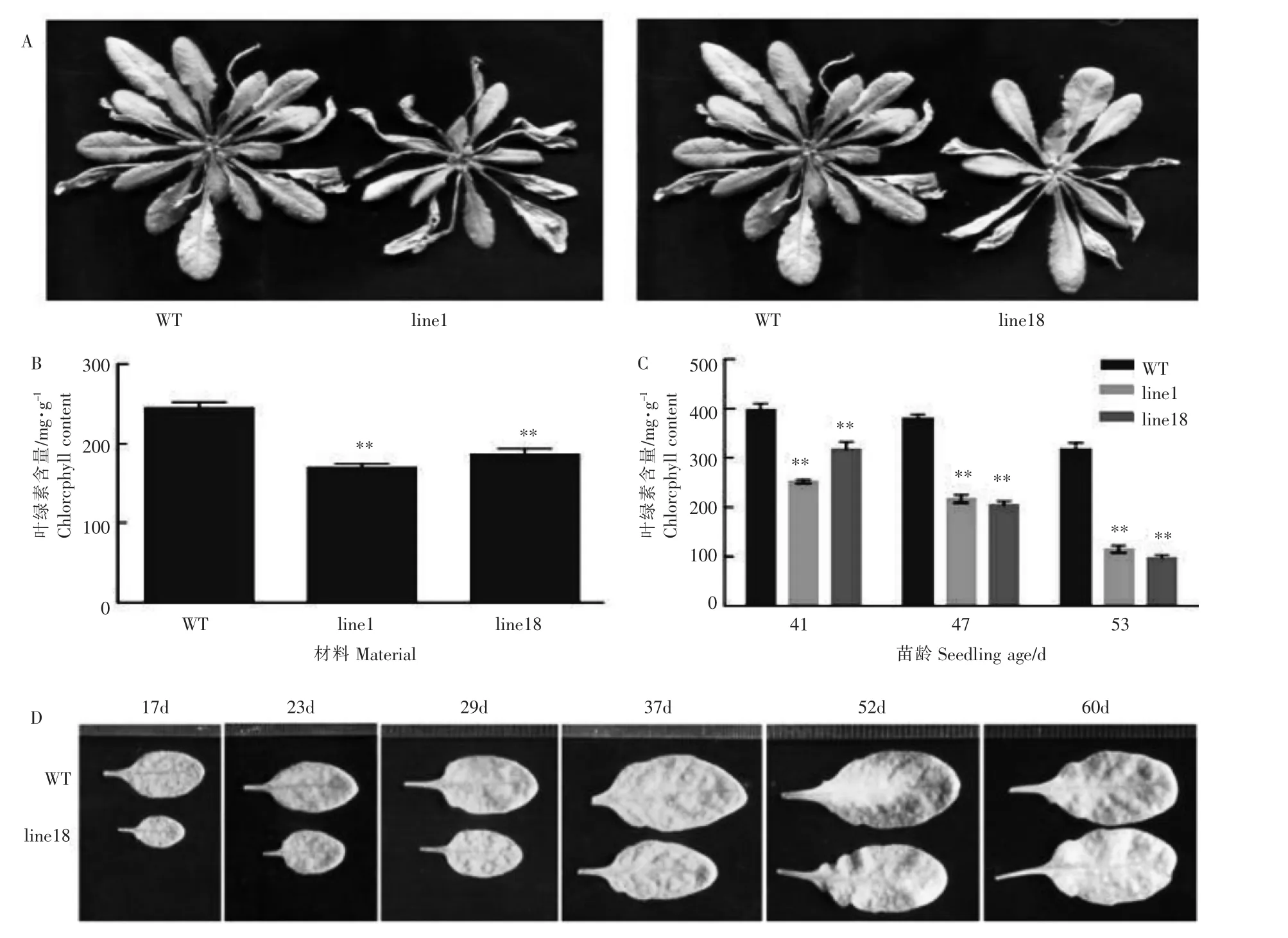
图2 35S-OsSAPP2转基因拟南芥在自然衰老过程中衰老表型及叶绿素含量变化Figure 2 Changes of senescence phenotype and chlorophyll content in 35S-OsSAPP2 transgenic Arabidopsis thaliana during natural aging
为了进一步研究OsSAPP2基因在叶片衰老中的作用,在成熟期对WT和转基因拟南芥进行观察。与WT相比,苗龄69d的35S-OsSAPP2转基因拟南芥莲座叶半径较小、叶色浅、萎蔫严重且衰老死亡数多,具有明显的早衰现象(图2A)。从图2B也可看出,35S-OsSAPP2转基因株系line1、line18莲座叶的叶绿素含量比野生型低。对于WT和35S-OsSAPP2转基因株系line18中不同叶龄的第5片莲座叶来说,随着叶龄的增加,过表达OsSAPP2基因的拟南芥叶片黄化和衰老的程度加速(图2C)。对不同苗龄的拟南芥第5和第6片莲座叶中的叶绿素含量进行测定,发现过表达OsSAPP2基因会导致转基因拟南芥叶绿素含量显著低于WT,且随着植物发育进程,转基因拟南芥叶绿素含量的下降速度要远大于WT(图2D)。
2.2 35S-OsSAPP2转基因拟南芥的衰老标志基因表达分析
在叶片发育的4个时期幼叶片期(YL)、成熟叶片期(NS)、开始衰老叶片期(ES)和衰老中期叶片期(LS),利用实时定量RT-PCR检测WT和转基因株系(line18)中拟南芥衰老标志基因SAG12,衰老关键转录因子NAC2、AtNAP、WRKY6,叶绿素降解关键基因ACD1和光合作用相关基因RbcS和RbcL的表达变化情况。由图3可知,SAG12在WT和line18株系的YL、NS、ES时期几乎检测不到,在LS时期表达量显著上升,且line18中的表达量显著高于WT;NAC2和NAP的表达量随着叶龄的增加而逐渐增加,且过表达OsSAPP2基因会显著促进这2个基因的表达。
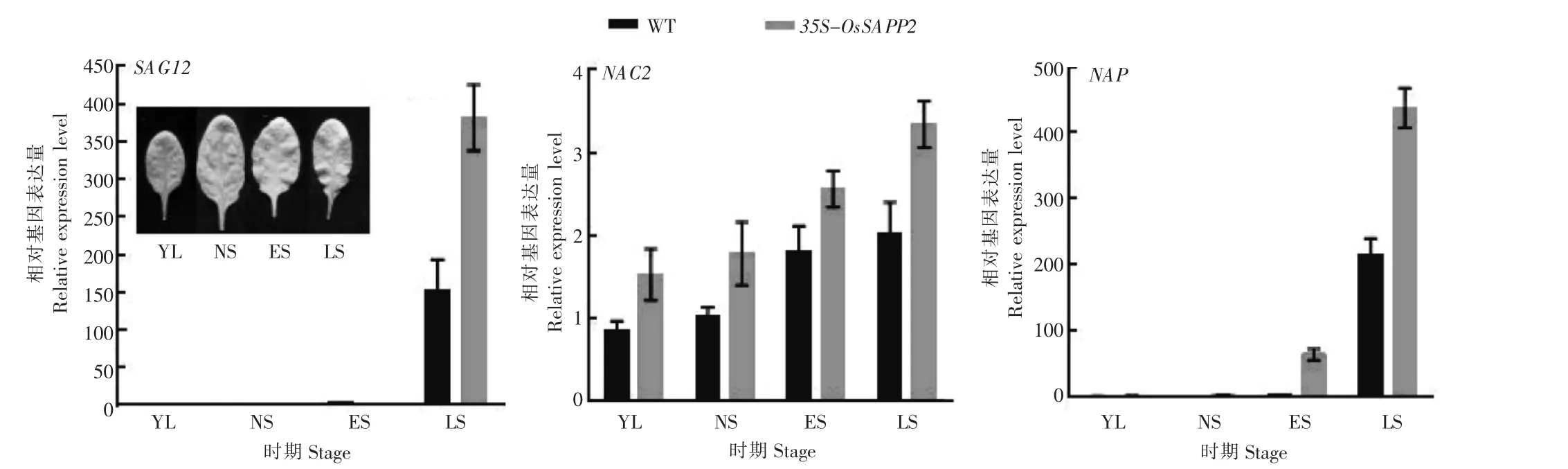
图3 野生型(WT)和35S-OsSAPP2转基因拟南芥在不同发育时期中衰老标志基因(SAG12、NAC2和NAP)表达变化Figure 3 Expression of senescence marker genes(SAG12,NAC2 and NAP)in wild-type(WT)and 35S-OsSAPP2 transgenic Arabidopsis at different developmental stages
WRKY6基因虽然在YL、NS、ES时期line18中的表达量较WT低,但是在LS时期,过表达OsSAPP2基因对WRKY6基因的表达促进作用大幅度上升。随着叶龄的增加,ACD1基因的表达量在WT和line18中逐渐增加,line18中的增加幅度显著高于WT;对于光合作用关键基因RbcS和RbcL来说,在YL和NS时期与WT相比过表达OsSAPP2基因会促进它们的表达,而在衰老开始和衰老进行时期(ES和LS时期),过表达OsSAPP2基因会导致它们的表达量快速下降(图4)。
2.3 黑暗处理促进由OsSAPP2基因过表达造成的拟南芥早衰
人工黑暗处理产生碳饥饿是一种常用的衰老诱导胁迫处理,在黑暗处理情况下对过表达OsSAPP2基因的转基因拟南芥进行表型分析和衰老标志基因表达量变化检测。20d苗龄的转基因拟南芥黑暗处理1d后,无论WT和35S-OsSAPP2转基因拟南芥叶片都有黄化迹象,但是35S-OsSAPP2转基因拟南芥的黄化衰老程度要比WT大;黑暗处理8d后,WT和35S-OsSAPP2转基因拟南芥叶片均出现深度黄化,转基因拟南芥黄化程度也明显大于WT,几乎没有绿色叶片;叶绿素含量测定结果与表型一致,即过表达OsSAPP2基因的转基因拟南芥株系在黑暗处理下,叶绿素含量大幅度降低,且降低速率大于WT(图5A和B)。选取苗龄25d的WT和转基因拟南芥第5片莲座叶,经过4d黑暗处理后,line1和line18转基因拟南芥叶片黄化程度明显大于WT,且叶片中间有萎蔫坏死症状(图5C)。黑暗处理前,35S-OsSAPP2转基因拟南芥第5莲座叶的叶绿素含量与WT无显著差异(p>0.05)。经4d黑暗处理后,35S-OsSAPP2转基因拟南芥叶片的叶绿素含量明显下降,具有极显著差异(图5D)。
利用实时定量RT-PCR检测黑暗处理1d和8d后,NAC2、AtNAP、WRKY6、ACD1、RbcS和 RbcL在 WT和转基因拟南芥line18株系的表达变化情况,随着黑暗处理的时间延长,过表达OsSAPP2基因的转基因拟南芥中,衰老关键转录因子NAC2、AtNAP、WRKY6和叶绿素降解关键基因ACD1的表达量均上升幅度要明显大于WT;由于黑暗处理后导致叶片衰老死亡,所以RbcS和RbcL基因的表达量大幅度降低,且转基因拟南芥中的表达量降低幅度显著高于WT(图6)。
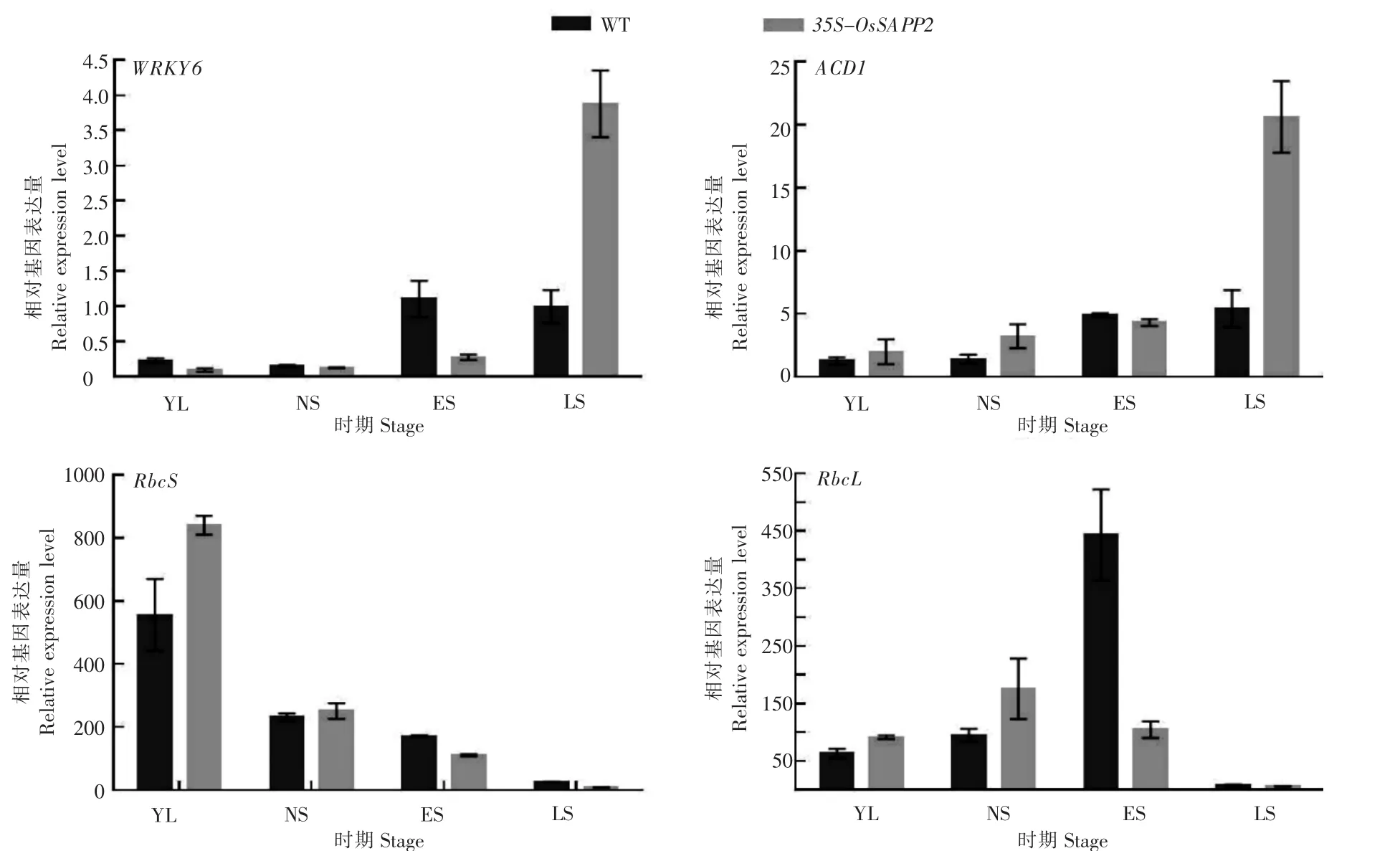
图4 野生型(WT)和35S-OsSAPP2转基因拟南芥在不同发育时期中衰老标志基因(WRKY6、ACD1、RbcS和RbcL)表达变化Figure 4 Expression changes of senescence marker genes(WRKY6,ACD1,RbcS and RbcL)in wild-type(WT)and 35S-OsSAPP2 transgenic Arabidopsis at different developmental stages
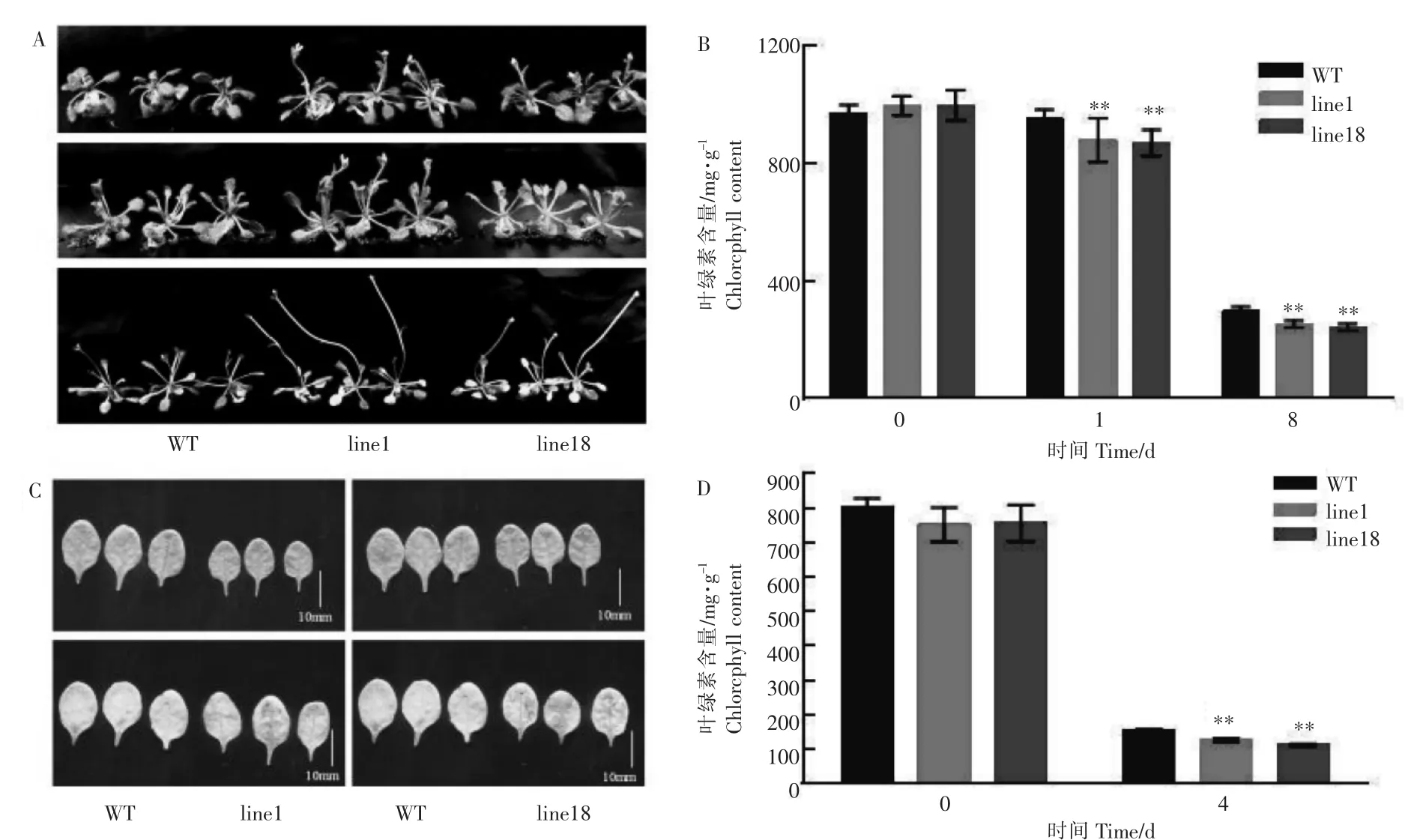
图5 黑暗处理下野生型(WT)和35S-OsSAPP2转基因拟南芥的表型和叶绿素含量变化Figure 5 Phenotypic and chlorophyll contents of wild type and 35S-OsSAPP2 transgenic Arabidopsis thaliana under dark conditions
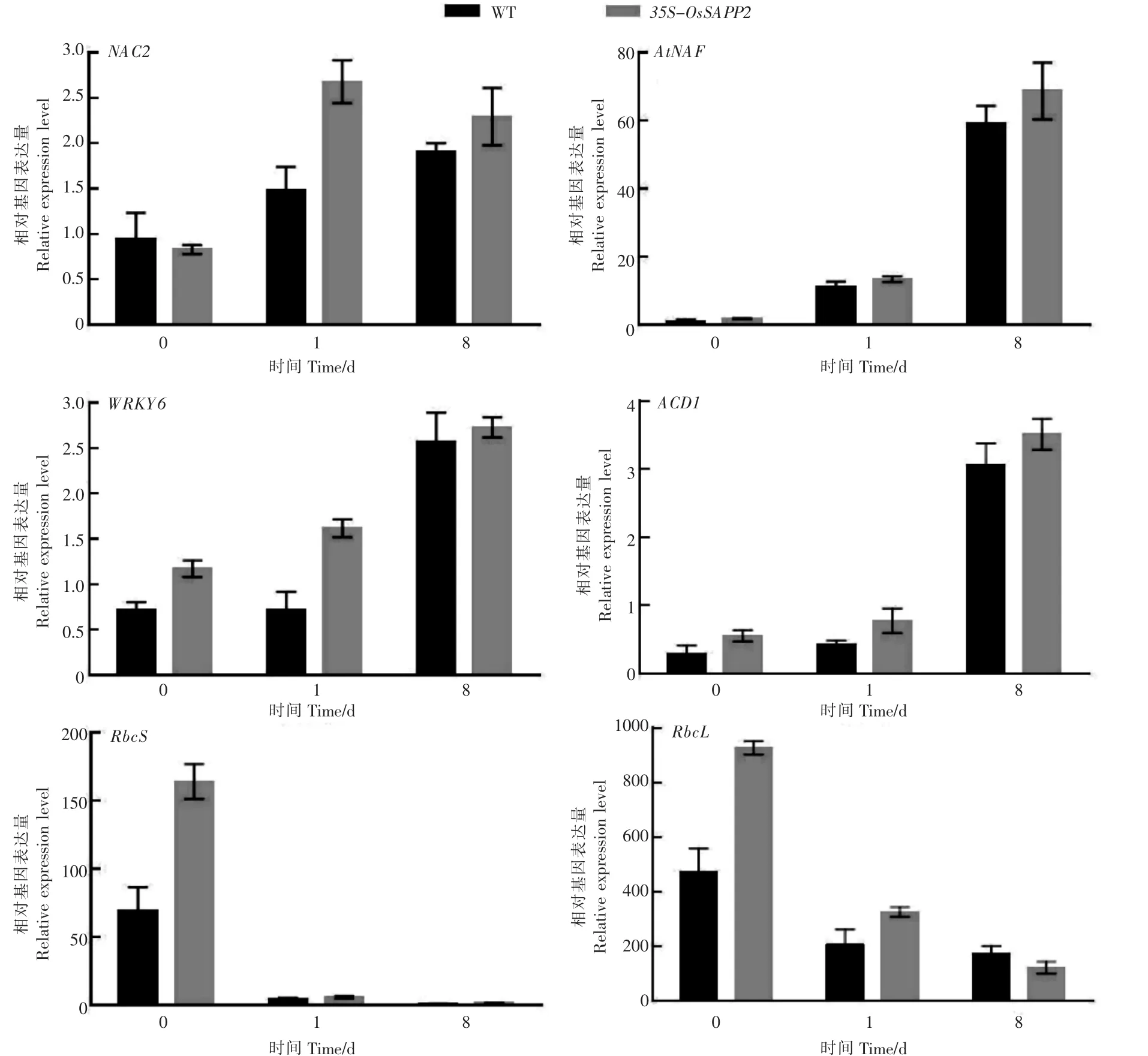
图6 黑暗处理下野生型和35S-OsSAPP2转基因拟南芥中衰老标志基因表达变化Figure 6 Changes of senescence marker gene expression in wild-type and 35S-OsSAPP2 transgenic Arabidopsis under dark conditions
3 讨论与结论
在叶片衰老过程中,叶肉细胞经历复杂的代谢变化和结构变化,叶绿体发生崩解,叶片光合作用能力下降,叶绿素、脂类和核酸等大分子物质发生降解;叶片衰老过程是植株营养物质循环再利用,是新生器官(如新生叶片或种子)营养物质供应的有益补充[18]。这是一个主动地发育过程,有大量基因对这个过程进行严格调控[19]。在农业生产方面,叶片衰老可以降低作物产量,当作物受到非生物胁迫时可以提前发生早衰,这种提前衰老的发生能降低作物一半的产量[5,20-21]。通过转录组学研究发现,在大麦叶片衰老过程中有11个蛋白磷酸酶编码基因的表达发生改变,但是对于叶片衰老过程中蛋白磷酸酶的研究仍然较少,如ZHANG等[13]在2012年从拟南芥中发现了第一个在叶片衰老调控中发挥作用的PP2C,SAG113,氨基酸序列和进化树分析表明SAG113是PP2C的A组成员,与ABI1和ABI2同源性较高;研究发现SAG113作为ABA信号的负调控子,参与叶片衰老过程中失水过程的调控,同时SAG113的表达与衰老重要转录因子AtNAP的表达具有协同性。另外,在拟南芥中还发现了一个对叶片衰老进行负调控的2C型蛋白磷酸酶SSPP,它通过与叶片衰老正调控的蛋白磷酸酶At-SARK互作而对叶片衰老进程进行调控[14-16]。
水稻是我国重要的粮食作物和禾本科功能基因组研究的模式植物,近年来研究已经鉴定出一系列参加水稻叶片衰老进程调控的基因可以调控水稻产量和品质,如Ghd2和Ghd7基因,PGL基因和OsFBK12基因等均通过不同分子途径促进或延缓植物叶片的衰老进程[22-24]。目前,关于水稻中参与叶片衰老进程调控的蛋白磷酸酶还没有报道。本研究发现并克隆的OsSAPP2是一个典型的2C型蛋白磷酸酶编码基因,经过生物信息学软件预测具有典型的PP2C结构域。根据XUE等[7]对水稻PP2Cs的聚类分析研究结果,水稻中PP2Cs共分为11个亚家族,OsSAPP2是亚家族D的成员,D家族成员多数是ABA信号通路的正调控因子;利用PlantCare网站进行在线分析,预测OsSAPP2基因CDS上游区域不仅含有核心元件TATA-box,还具有脱落酸响应元件(ABRE)、光响应元件(AE-box、G-box)和MBS结合位点等其他调控元件。在本研究中发现,过表达OsSAPP2的转基因拟南芥株系莲座叶直径变小、莲座叶数量减少,抽苔和开花时间提前等(图1,图5A,表1);这些结果表明35S-OsSAPP2转基因拟南芥提前从营养生长进入生殖生长,整体植株的衰老进程加速。已有研究证明黑暗诱导会促进叶片衰老,而黑暗诱导也常常用于叶片衰老研究中[25-26]。
在叶龄依赖的自然衰老和人工黑暗处理导致的衰老情况下,过表达OsSAPP2基因的转基因拟南芥与WT相比,叶绿素含量较低,叶绿素降解加快,叶片黄化萎蔫情况严重,且黑暗处理会加速转基因拟南芥的衰老程度(图2、图5),叶绿素含量检测与表型相符,说明OsSAPP2基因响应黑暗胁迫的处理。另外,通过实时定量RT-PCR检测,发现OsSAPP2基因的表达量随着黑暗诱导处理时间的延长而上调,光合作用相关基因RbcS和RbcL在衰老叶片中表达量大幅度下降,而叶绿素降解关键酶编码基因ACD1表达量上升(图6);这也暗示了过表达OsSAPP2基因导致转基因拟南芥的叶绿素降解加速,光合作用能力下降。RbcS和RbcL基因参与光合作用途径,在衰老期表达量下降显著且迅速,推测由于OsSAPP2受衰老调控促进表达从而下调了它们的表达,导致光合作用下降迅速,光合产物形成减少显著,加速衰老。推测OsSAPP2介导的衰老信号也有可能通过促进叶绿素降解而发挥对叶绿素含量的调控作用,同时影响着叶片的光合作用能力。SAG12基因编码半胱氨酸蛋白酶,现有研究证明它仅在只依赖于叶龄发育信号而导致的衰老过程中表达,表达量不受其他胁迫信号的影响,因此SAG12是自然衰老的标志基因。本研究在衰老叶片时期(LS)才能在WT和转基因株系中检测到这个基因的表达,并且过表达OsSAPP2基因会显著促进它的表达量;这个结果暗示着OsSAPP2基因在叶片衰老过程中响应叶龄发育信号。在植物叶片衰老过程中,NAC和WRKY家族成员中大部分的转录因子表达量上调,参与叶片衰老的调控,是叶片衰老的关键转录因子[27];在本研究中,无论是叶龄依赖的自然衰老条件下还是黑暗诱导的衰老条件下,都发现OsSAPP2基因的过表达会显著促进NAC2、AtNAP和WRKY6基因的表达,且在黑暗处理条件下,转基因植物中的表达上升幅度远大于WT;暗示着OsSAPP2基因可能通过调节这些叶片衰老的关键转录因子从而参与叶片衰老的调控。
已有报道显示,改变植物的衰老进程可以影响产量,延缓衰老进程可以增加产量,如有研究者观察到过表达OsFBK12转基因水稻的叶片衰老被延缓,且籽粒大小增加的现象[24];在本研究中发现,与WT相比35S-OsSAPP2转基因拟南芥株系line1和line18的种子形态发生改变,种子长宽比变小,种子的产量和千粒重下降,这种产量的降低可能是由于OsSAPP2基因过表达导致的早衰有关,这与前人研究结果一致。综合以上研究结果,过表达OsSAPP2基因可以导致转基因拟南芥叶片发生早衰,因此该基因参与植物叶片衰老的调控,是正向调节因子。
