微小RNA在tau蛋白过度磷酸化中的作用
2019-04-22刘佳惠徐丹宋波廖华强应明徐慧文郭超钦黎玲玲罗友根
刘佳惠,徐丹,宋波,廖华强,应明,徐慧文,郭超钦,黎玲玲,罗友根*
(1井冈山大学医学院神经退行性疾病与衰老研究中心,吉安,343009;2吉安市吉州区疾病预防控制中心,吉安,343000)
阿尔茨海默病(Alzheimer’s disease, AD)是一类常见的神经退行性疾病,主要临床表现为学习记忆障碍、失语、失忆、失认、视空间技能损害,抽象思维改变等[1]。其主要病理表现为老年斑(senile plaque, SP)和神经元纤维缠结(neurofibrillary tangles, NFTs)及进行性突触损伤与神经元丢失[2]。老年斑由β-淀粉样蛋白(β-amyloid protein, Aβ)沉积而成,过度磷酸化的tau蛋白是神经纤维缠结的重要组成成分。目前关于AD发病机制的主要学说包括:淀粉样肽级联反应、微管相关tau异常、氧化应激、炎症介质、胆碱功能降低等学说。
微小RNA(microRNA, miRNA)是由内源基因编码的一类长度在20~24个核苷酸之间的单链非编码RNA分子,能负向调控靶mRNA表达,最终导致mRNA降解或翻译抑制[3]。研究表明MiRNA与AD发病密切相关[4]。本文就微小RNA在tau蛋白过度磷酸化中作用的相关研究进行简单叙述。
1 Tau蛋白过度磷酸化在AD发病中的作用
细胞内tau蛋白过度磷酸化是AD最早出现的病理改变之一。Dermaut[5]等研究发现,NFTs数量与AD痴呆程度呈正相关。NFTs中的主要成分成对螺旋丝的亚单位是过度磷酸化的tau蛋白。Tau蛋白是一种神经元微管相关细胞骨架蛋白,存在于正常神经元内,主要集中于脑内神经元轴突之中,其主要功能是与微管相关蛋白结合聚合形成微管,维持微管稳定性,并在神经元可塑性中发挥重要作用。Tau蛋白磷酸化水平由催化磷酸化反应的蛋白激酶和催化去磷酸化反应的蛋白酯酶的活性共同调节。当tau蛋白磷酸化/去磷酸化出现平衡紊乱时,tau蛋白过度磷酸化形成双螺旋丝(paired helical filaments, PHF)及NFTs,导致细胞骨架稳定性遭到破坏,在脑中沉积而使神经元变性[6]。研究表明,过度磷酸化的tau蛋白促微管组装的生物学活性丧失,对蛋白水解酶抗性提高,从而产生神经毒性[7]。
2 MiRNA在AD发病中的作用
MiRNA在中枢神经系统内分布广泛,可调节神经发育、分化、成熟[8]。Schonorock等研究发现,AD患者脑组织中存在miRNA失调[9]。AD患者脑组织内异常表达的miRNA作为重要的基因转录后负性调节因子参与Aβ聚积、神经纤维缠结等神经退行性病理变化的形成[10]。AD患者脑内miR-132-3P表达水平降低和miR-125b过表达均能诱导tau蛋白过度磷酸化[11,12]。有研究显示,可溶性泛素羧基末端水解酶 -1(ubiquitin C-terminal hydrolase L1, UCHL1)的降低会导致AD,其机制与诱导tau蛋白过度磷酸化有关[13]。MiR-922是UCHL1的负性调控因子,可通过与UCHL1的3’-UTR结合抑制其表达,从而促进tau蛋白过度磷酸化[14]。MiR-146a在AD中过表达,通过抑制ROCK1蛋白的翻译,导致神经元PTEN磷酸化的降低,进而引起tau蛋白过度磷酸化[15]。
3 MiRNA调控tau蛋白表达
进行性核上性麻痹(progressive supranuclear palsy, PSP)是一种tau蛋白相关神经系统疾病。PSP患者脑组织中miR-132表达下调,使靶基因神经元多聚嘧啶序列结合蛋白2(polypyrimidine tract-binding protein 2, PTBP2)表达上调,从而调控tau蛋白外显子10的剪接[16]。同样,miR-124和miR-9通过调控特定剪接调节因子,对内源性tau蛋白外显子10的剪接进行调节[16,17]。Dickson等[18]在人类神经母细胞瘤细胞系M17D和SH-SY5Y中采用双荧光素酶报告基因技术发现tau蛋白3’-UTR中存在miR-34a结合位点,miR-34a与tau 蛋白3’-UTR特异性结合使tau mRNA和蛋白表达下调。
4 MiRNA调控磷酸激酶的活性
根据蛋白激酶催化tau蛋白磷酸化反应序列特点可将其分为脯氨酸依赖性蛋白激酶(PDPK)和非脯氨酸依赖性蛋白激酶(non-PDPK)两大类。其中PDPK大致可分为三类:①有丝分裂原激活的蛋白激酶(mitogen-activated protein kinase, MAPK)及其三种主要亚型:细胞外信号相关蛋白激酶(extracellular signal-regulated kinase, ERK)、c-Jun氨基末端激酶(c-Jun N-terminal kinase, JNK)、p38MAPK。②周期蛋白依赖性激酶(cyclin dependent kinase, CDK),如CDK5;③糖原合成酶激酶-3(glycogen synthase kinase-3, GSK-3)。Non-PDPK包括:钙/钙调素依赖性蛋白激酶Ⅱ(calcium/calmodulin-dependent protein kinase Ⅱ, CaMKⅡ)、蛋白激酶A、蛋白激酶C、Ser/Thr蛋白激酶B及酪氨酸激酶1、酪氨酸激酶2等[19]。
4.1 MiRNA调控MAPK的活性
ERK1是一种直接的tau蛋白激酶。现有资料显示miR-15a在AD患者脑内表达下调。进一步研究证实miR-15a对ERK1具有靶向调控作用,从而影响tau蛋白磷酸化[20]。帽结合蛋白4EHP是哺乳动物miRNA诱导沉默复合物的关键组成部分,介导基因沉默。研究发现双特异性磷酸酶6(dual specificity phosphatase 6, DUSP6)mRNA被帽结合蛋白4EHP和miR-145所抑制,促进ERK1/2磷酸化和细胞生长,减少凋亡[21]。据该研究推测,miR-145可能在AD样tau蛋白过度磷酸化中起到了一定的调控作用。MiR-125b的靶标之一,DUSP6,也称为丝裂原活化蛋白激酶磷酸酶3,是ERK 1/2的负反馈调节因子,通过去磷酸化使ERK 1/2失活[22]。在AD患者脑内miR-125b通过负调控DUSP6激活ERK1/2,导致tau蛋白过度磷酸化[11]。
4.2 MiRNA调控CDK5的活性
CDK5是调节tau蛋白磷酸化最重要的蛋白激酶之一[23]。Aβ低聚物促进Ca2+内流,通过钙调蛋白酶激活启动CDK5/p35向CDK5/p25转化,且CDK5/p25复合物的积累引起CDK5长时间激活,导致脑中tau蛋白过度磷酸化[24]。
视网膜母细胞瘤蛋白(retinoblastoma protein,RB1)是一种主要的肿瘤抑制因子,通过调节细胞周期G1信号通路进而影响tau蛋白磷酸化水平。AD患者脑内miR-26b表达增加,其与RB1信使RNA直接结合并减少RB1表达,促进RB1的磷酸化,进而使CDK5活性增加,导致tau蛋白过度磷酸化[25]。另有研究表明,miR-125b可以下调DUSP6诱发ERK1/2的激活最终使CDK5活化,导致tau蛋白出现多位点磷酸化[11]。Sun等研究发现miR-195可以与CDK5R1 3’-UTR结合通过抑制p35表达来降低CDK5活性,从而降低神经元tau蛋白磷酸化水平[26]。Wang等研究结果表明miR-132通过NOS1信号间接抑制CDK5活性,使tau蛋白磷酸化水平下降[27]。
4.3 MiRNA调控GSK-3β的活性
在AD病理过程中,GSK-3β是催化tau蛋白过度磷酸化的主要激酶[28]。在AD模型中miR-138表达增高,过表达miR-138可以通过靶向调控视黄酸α受体(Retinoic acid receptor alpha, RARA)激活GSK-3β,从而促进tau蛋白的磷酸化[29]。在AD细胞模型中表达下调的miR-124-3p通过抑制Caveolin-1/PI3K/Akt信号通路激活GSK-3β,致使tau蛋白异常过度磷酸化,诱导神经细胞凋亡[30]。转染miR-124-3p模拟物能逆转tau过度磷酸化,减少细胞凋亡,从而发挥神经保护作用。另有研究报道GSK-3β为miR-132-3p的靶基因,与GSK-3β直接结合下调后者的表达从而降低tau蛋白磷酸化水平[12]。
5 MiRNA调控磷酸酯酶的活性
磷酸酯酶可催化tau蛋白去磷酸化。哺乳动物丝/苏氨酸磷酸酯酶有5种,即PP1、PP2A、PP2B、PP2C、PP5。除PP2C外其它4种均能导致tau蛋白去磷酸化,其中PP2A是最重要的酯酶,其次为PP1、PP5、PP2B,其活性分别占人脑tau磷酸酯酶总活性的71%、11%、10%、7%[31]。蛋白磷酸酯酶2A(protein phosphatase 2A , PP2A)是人脑中调节tau蛋白磷酸化的主要酯酶,可以作用于人脑tau蛋白多个磷酸化位点,其活性与tau蛋白磷酸化水平呈负相关。Gong等研究发现在大鼠脑切片中PP2A抑制剂冈田酸引起tau蛋白过度磷酸化[32]。
已有研究提示雌激素对AD发生发展产生一定影响[33]。雌激素受体分为ERα和ERβ。研究表明,ERα通过上调miR-218来抑制其靶蛋白酪氨酸磷酸酶(protein tyrosine phosphatase, PTPA)的表达,导致PP2A失活而GSK-3β激活,促进tau蛋白磷酸化;相反,ERβ抑制miR-218表达水平从而降低tau蛋白磷酸化水平[34]。Banzhaf等发现miR-125b能够使蛋白磷酸酯酶1催化亚基α(protein phosphatase 1 catalytic subunit α isoform, PPP1CA)表达下调从而诱导tau蛋白磷酸化[11]。Smith等采用免疫印迹、RTPCR、荧光素酶报告基因研究技术在三转基因AD小鼠(3xTg-AD)模型中发现,miR-132/212缺乏使PP2B表达下调而GSK-3β被激活,引起tau蛋白过度磷酸化,从而参与AD发病[35]。
6 总结与展望
MiRNA已被证实在神经系统中广泛分布,参与中枢发育、神经元分化和突触塑形等过程,通过对突触可塑性的调节参与形成学习记忆。Tau蛋白过度磷酸化被认为是AD发病主要机制之一。大量数据证实miRNA与AD发病密切相关。AD患者脑内miRNA表达异常,后者通过调控蛋白激酶和/或蛋白酯酶活性影响tau蛋白磷酸化水平参与AD发病。由此推断,miRNA有望成为AD诊断与防治的新的生物标记物和靶点。本文就AD患者或模型中异常表达的miRNA对tau蛋白磷酸化的作用及机制进行了总结(表1)。
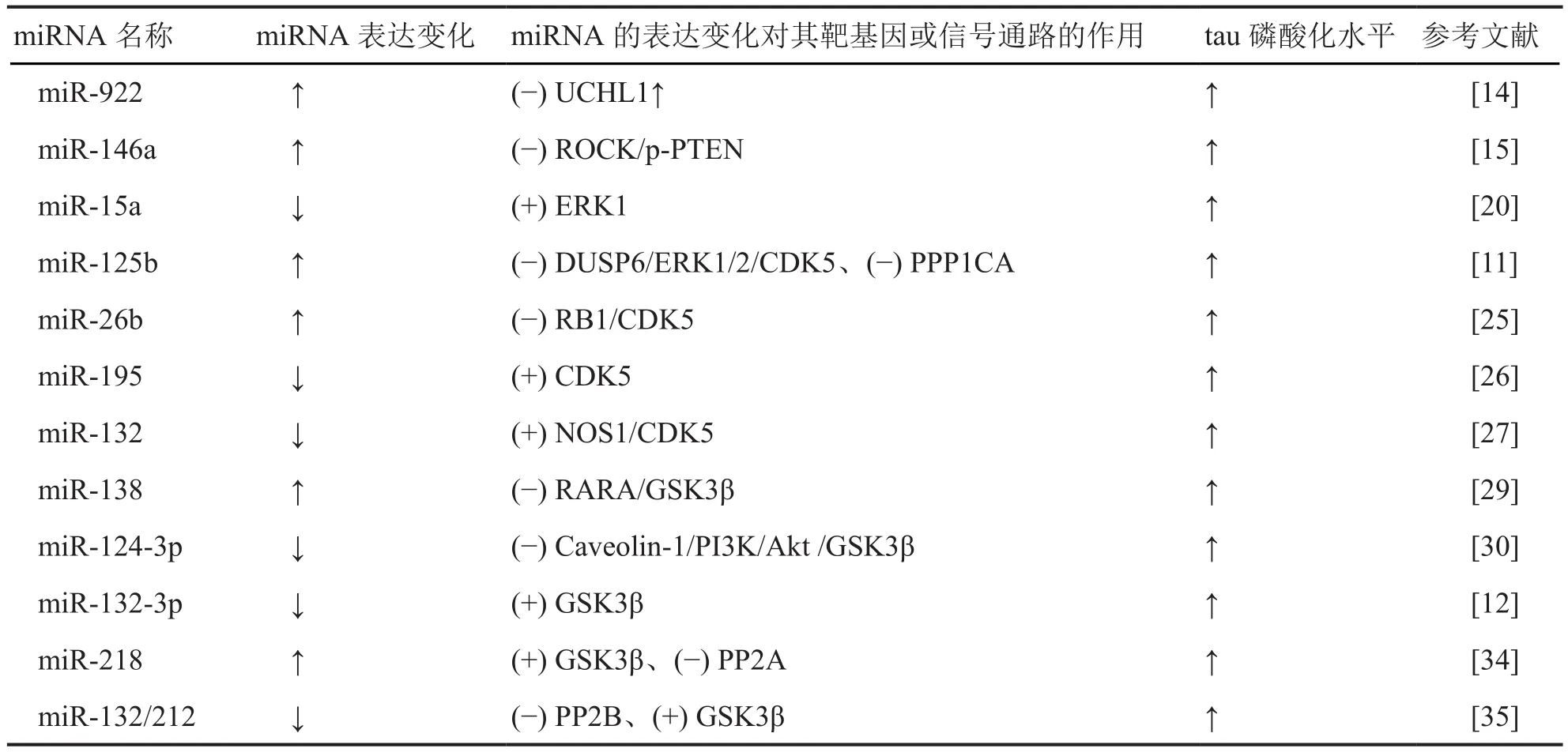
表1 AD患者或模型中异常表达的miRNAs对tau蛋白磷酸化的作用及机制Tab.1 The role and mechanism of abnormally expressed miRNAs in the hyperphosphorylation of tau protein among AD patients or models
近年来关于miRNA在AD样tau蛋白过度磷酸化中的作用越来越受到重视,已成为学者们研究的热点。然而,对于如何通过靶向调控miRNA抑制tau蛋白过度磷酸化,防止AD的发生发展,是一个亟待解决的问题。
