施氮及根系分隔处理对间作尾巨桉和降香黄檀幼苗光合生理特性的影响
2019-04-16廖良宁汤文艳龙佳峰许峻模朱德意叶绍明
廖良宁 汤文艳 潘 婷 龙佳峰 许峻模 朱德意 叶绍明
( 1. 广西大学林学院,广西 南宁 530004;2. 湖南中彩生态环境科技有限公司,湖南 长沙 410000;3. 广西壮族自治区林业勘测设计院,广西 南宁 530004)
氮素作为植物体内重要的营养元素,是叶绿素的主要组成成分,直接参与植物细胞分裂、生理代谢、光合特性以及产量形成[1],在植物生长过程和光合作用中均起重要作用。植物缺氮会抑制植物生长发育,导致叶绿素含量、光合速率及生物量下降[2]。施氮可延长植物生长周期[3]和延缓叶片衰老[4],有利于提高植物叶面积指数、叶绿素含量、光能利用率与光合作用效率等,从而促进植物生长,增加植物生物量[5-7]。一定程度上,氮素直接决定了光合作用,亦间接决定植物干物质累积,尤其时对氮素需求量较大的速生树种生长具有显著提高作用[8]。但是速生树种对氮素的利用率极低,故提高速生树种对氮素的利用率是林业领域重要研究方向。
桉树(Eucalyptussp.)是中国南方的主要造林树种之一,广泛分布于广西、云南等地,传统的纯林栽培模式下,其氮肥利用效率低,且多代连栽可导致地力衰退等一系列问题[9]。桉树与豆科树种混交是解决这些问题的重要手段,如台湾相思与桉树混交改善了土壤结构、微生物生理类群,提高土壤养分的利用率[8]。桉树与台湾相思混交优势主要通过地上与地下部分的协同作用,促进植物对养分的吸收利用,但是该结果是否也同样存在于桉树与其他豆科树种混交模式中值得进一步研究。本研究以尾巨桉(Eucalyptus urophylla×Eucalyptus grandis)和降香黄檀(Dallergia odorifera)幼苗为研究对象,探讨了不同氮素水平及根系分隔方式对尾巨桉与降香黄檀幼苗叶绿素含量、保护酶活性、光能利用率及光合特性的影响,分析间作体系地上部分资源互补的相关性,探讨氮素与根系分隔对尾巨桉和降香黄檀间作体系地上部分产生影响的相关机制,为营造桉树混交林提供参考依据。
1 材料与方法
1.1 试验设计
试验在广西大学林学院试验基地大棚内(108°17′09″E, 22°50′28″N)进行,于 2016 年 2 月15日选取3月生的尾巨桉和1年生的降香黄檀进行盆栽(口径40 cm,高45 cm),设3种根系分隔模式:1) 不分隔 (T1),根系互作与种间竞争关系同时存在;2) 30 µm孔径尼龙网分隔(T2),存在种间竞争关系,但消除根系互作关系;3) 2 mm厚度塑料分隔模式(T3),消除种间竞争与根系互作关系,相当于单植。其中尼龙网及塑料膜将盆栽从中间平均分隔,保证生长空间一致,种植比例为1∶1。土壤取自良凤江国家公园马尾松林中龄林林地 (108°17′24″E, 22°55′48″N),去除石粒及杂质,将其风干混匀珍珠岩保持透水性 (m(泥土)∶m(珍珠岩)=34∶1),装满盆钵后共35 kg,其中土壤全氮含量为0.42 mg/g,全磷含量为0.3 mg/g,全钾含量为11.85 mg/g,硝态氮含量为0.11 mg/g。设3个氮肥水平(氮肥为普通尿素,氮含量46.7%):分别为不施肥 (N0)、3.0 g/盘 (N1)、6.0 g/盘 (N2)。种植前将1/2的尿素溶于水均匀的施入土壤,苗木定根后将另一半溶于水,用注射法施入土壤。试验过程中严格控水,采用刻度壶进行浇水,每盆浇水量一致,且保证其他生长条件一致。每个处理4次重复,随机排列。
1.2 测定方法
于2016年8月13日—8月14日的晴朗天气,使用Li-3000C叶面积仪(Li-Cor公司,美国)测定尾巨桉与降香黄檀幼苗叶面积指数,采用Li-6400便携式仪光合测定系统分析仪(Li-Cor公司,美国) 对苗木生长良好的叶片进行定时(每日9:00—11:30)、定位测定净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)。每个重复测定8片叶。2016年8月15日采用IMAGING-PAM(上海泽泉科技股份有限公司,中国)对尾巨桉与降香黄檀幼苗进行叶绿素荧光参数测定,测定前将幼苗进行暗处理30 min,Fv/Fm模式下测定光系统II (PS II) 的实际光化学量子效率(Y(II)) 、PS II处调节性能量耗散的量子产量(Y(NPQ))、光化学淬灭系数(qP)、非光化学淬灭系数(qN)和相对电子传递速率(ETR),每个处理测定重复5次。选取新鲜叶片测定相关生理指标,采用丙酮—乙醇混合提取法测定叶片叶绿素含量,采用蒽酮比色法测定可溶性糖 (SS) 含量,分别采用NBT光化还原法、紫外吸收法以及俞创木酚法测定植物超氧化物歧化酶 (SOD)活性、过氧化氢酶 (CAT) 活性和过氧化物酶 (POD) 活性[10]。光合及生理指标测定完毕后,将苗木按根、茎、叶收集,置于70 ℃烘箱中烘干至恒重并称量各部位干物质积累量。
1.3 分析方法
对测定指标进行描述性统计分析,描述性统计包括平均值、标准差等,分别采用Shapiro-Wilk检验法和levene检验法对各指标进行正态分布假设检验和方差齐性检验,若指标符合正态分布且方差齐性的假设成立,则进行方差分析和多重比较(LSD)。数据分析和图形制作分别通过SPSS 19.0和Sigmaplot 10.0完成。
2 结果与分析
2.1 氮素水平与根系分隔处理对幼苗叶面积指数的影响
氮素水平与根系分隔处理尾巨桉与降香黄檀幼苗叶面积指数差异见图1。由图1可知,施氮显著提高了尾巨桉与降香黄檀叶面积(P<0.05)。在T1模式下,施氮显著增加尾巨桉与降香黄檀幼苗叶片比叶重 (P<0.05)。N2显著提高了T2、T3的叶片比叶重(P<0.05)。不同根系分隔处理,尾巨桉叶面积变化规律表现为T1>T3>T2,降香黄檀则表现为T3>T1>T2;尾巨桉比叶重以T1比叶重最大,降香黄檀则是T3最高。尾巨桉幼苗叶片叶面积与比叶重均以T1N2处理组合最佳,降香黄檀则是T3N2组合最佳。说明施氮促进尾巨桉与降香黄檀叶幼苗叶片面积指数的增长,但不同根系分隔对2个树种叶面积指数产生的影响不同。
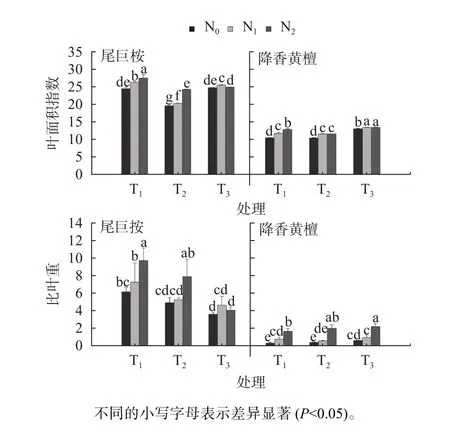
图1 氮素水平与根系分隔处理尾巨桉与降香黄檀幼苗叶面积指数差异Fig. 1 Differences in leaf area of E. urophylla × E.grandis and D. odorifera under different nitrogen levels and root partition treatments
2.2 氮素水平与根系分隔处理对幼苗叶绿素含量的影响
氮素水平与根系分隔处理尾巨桉与降香黄檀叶绿素含量差异见表1。由表1可知,随着氮素水平增加,尾巨桉和降香黄檀的叶绿素a、叶绿素b与叶绿素总含量呈递增趋势变化。尾巨桉T1的叶绿素含量均高于T2与T3,同一氮素尾巨桉叶片叶绿素a含量无显著差异,但N2水平下,T1显著高于T2和T3(P<0.05)。叶绿素b含量在不同施氮水平处理中,T1显著高于T2和T3(P<0.05)。尾巨桉叶绿素含量以T1N2最高,比N0提高39.13%。降香黄檀则以T3N2的数值最大,比N0提高19.63%,分别比 N2水平的 T2与 T1高 21.88%、18.90%。
2.3 氮素水平与根系分隔对叶片光能利用的影响
2.3.1 不同处理对幼苗叶片Y(II)和Y(NPQ)的影响
施氮与根系分隔对尾巨桉与降香黄檀叶片叶绿素荧光参数的影响见表2。由表2可知,施氮处理提高尾巨桉与降香黄檀幼苗Y(II),但降香黄檀N1与N0差异不显著,整体表现为N2>N1>N0。N0的根系分隔对尾巨桉及降香黄檀的Y(II)和Y(NPQ)影响均不显著,尾巨桉N1、N2水平T1Y(II)显著高于T2和T3(P<0.05);降香黄檀N1水平下T3与T1的Y(II)值显著高于N0和N2(P<0.05)。尾巨桉与降香黄檀幼苗Y(NPQ)在同一氮素水平不同模式处理差异不显著。说明施氮对Y(II)和Y(NPQ)的影响较大,但不同分隔模式对此影响较小。

表1 氮素水平与根系分隔处理尾巨桉与降香黄檀叶绿素含量差异Table 1 Differences in chlorophyll content of E. urophylla × E. grandis and D. odorifera under nitrogen levels and root partition treatments mg/g

表2 施氮与根系分隔对尾巨桉与降香黄檀叶片叶绿素荧光参数的影响Table 2 Effects of nitrogen levels and root partition treatments on chlorophyll fluorescence parameters of E. urophylla × E. grandis and D. odorifera leaves
2.3.2 不同处理对幼苗叶片qP、qN和ETR的影响
由表2可知,施氮显著提高尾巨桉幼苗的qP、qN,N2显著提高降香黄檀qN(P<0.05)。大部分施氮处理显著提高尾巨桉与降香黄檀ETR (P<0.05),但N1、N2差异较小。根系分隔对ETR影响也较小,对尾巨桉qP和qN的影响均不显著;但降香黄檀T3优于T1与T2。两者交互作用对尾巨桉与降香黄檀光能利用影响不显著。
2.4 氮素水平与根系分隔对幼苗光合特性的影响
2.4.1 不同处理对幼苗叶片Pn的影响
氮素水平与根系分隔尾巨桉和降香黄檀Pn差异见图2。由图2可知,尾巨桉和降香黄檀的净光合速率随氮素水平的增加而增加。氮肥显著提高3种栽培模式的尾巨桉Pn(P<0.05),T1表现最好。N0水平下,3种间作模式Pn差异不显著,N1、N2水平下T1显著高于T2与T3(P<0.05)。T1N2的Pn最高,比N0提高20.93%。降香黄檀T3的Pn在不同氮素水平均显著高于T1与T2(P<0.05),其中N2的Pn最高,比N0提高71.06%。T1与施氮均能促进桉树幼苗叶片Pn,但降香黄檀在T3条件下Pn最大。

图2 氮素水平与根系分隔对尾巨桉和降香黄檀Pn差异Fig. 2 Differences in Pn of E. urophylla × E.grandis and D. odorifera under nitrogen levels and root partition treatments
2.4.2 不同处理对幼苗叶片Gs的影响
氮素水平与根系分隔处理尾巨桉和降香黄檀Gs差异见图3。

图3 氮素水平与根系分隔处理尾巨桉和降香黄檀Gs差异Fig. 3 Differences in Gs of E. urophylla × E. grandis and D. odorifera under nitrogen levels and root partition treatments
由图3可知,施肥能显著提高尾巨桉与降香黄檀的气孔导度。T1模式下,尾巨桉与降香黄檀的Gs在不同氮素水平处理均显著高于T3与T2(P<0.05),尤其在N0条件下差异最大,尾巨桉T1分别比T2和T3高37.04%和60.87%,降香黄檀T1均比T2与T3高出20.00%。2种不同树种均以T1模式N1水平的Gs值最高。说明尾巨桉与降香黄檀间作互惠互利,起到生态位互补作用。
2.4.3 不同分隔模式和施肥处理下幼苗Ci的变化
氮素水平与根系分隔处理尾巨桉和降香黄檀Ci差异见图4。由图4可知,幼苗Ci与Pn、GS的变化趋势相反,随着施氮水平的增加,尾巨桉与降香黄檀Ci显著降低 (P<0.05)。在不同氮素处理条件下,尾巨桉均以T3最大,分别高于T2与T1的N05.98%和15.07%,差异达到显著水平(P<0.05)。降香黄檀N0与N1水平均以T2最高,N2水平以T1间作模式最高,与T2模式差异显著 (P<0.05),但与T3差异不显著。降香黄檀幼苗叶片GS最大的为T2模式N0,分别比T1与T3提高7.25%和7.60%。

图4 氮素水平与根系分隔处理尾巨桉和降香黄檀Ci差异Fig. 4 Differences in Ci of E. urophylla × E. grandis and D. odorifera under nitrogen levels and root partition treatments
2.4.4 不同分隔模式和施肥处理下幼苗Tr的变化
氮素水平与根系分隔处理尾巨桉和降香黄檀Tr差异见图5。由图5可知,在不同间作模式处理中,施氮能显著提高尾巨桉和降香黄檀的Tr(P<0.05)。同一氮素水平处理的T1模式下,尾巨桉的Tr均显著高于T3与T2(P<0.05),尾巨桉以T1N2处理的Tr最大,比N0高47.34%,且分别比T2与T3的同一氮素水平提高3.05%和35.12%。降香黄檀在N0表现为T1显著高于T2与T3(P<0.05)。N1、N2水平则以T3最高。表明施氮可以显著提高两不同树种蒸腾速率。而在施肥情况下,不隔间作提高了桉树的Tr,但降低了降香黄檀Tr。

图5 氮素水平与根系分隔处理尾巨桉和降香黄檀Tr差异Fig. 5 Differences in Tr of E. urophylla × E. grandis and D. odorifera under nitrogen levels and root partition treatments
2.4.5 光合作用指标方差分析
根系分隔与施氮的条件下幼苗光合作用指标方差分析结果见表3。由表3可知,施氮对尾巨桉和降香黄檀光合作用的影响是显著的 (P<0.05)。而根系分隔对尾巨桉光合作用各指标的影响也达到了显著水平 (P<0.05),但是对降香黄檀的Gs和Ci的影响不显著。同时根系分隔与施氮交互作用对光合作用的影响也不显著。
2.5 不同处理下幼苗可溶性糖含量的变化
施氮与根系分隔处理尾巨桉与降香黄檀SS含量差异见图6。由图6可知,施氮显著提高桉树与降香黄檀幼苗SS含量,但T3下N1、N0水平无显著差异。对比不同根系分隔处理,尾巨桉幼苗均以T1含量最高,施氮处理时显著高于其他2种种植模式(P<0.05),T1的N2水平SS含量最高,比N0提高8.17%,比N2水平T2、T3分别提高了5.15%和6.74%;但降香黄檀则以T3种植模式显著优于T1与T2(P<0.05)。说明T1间作与施氮对尾巨桉SS含量具有显著提升作用,但T1抑制降香黄檀SS的产生。

表3 根系分隔与施氮的条件下幼苗光合作用指标方差分析Table 3 Variance analysis of the photosynthetic characteristics index of seedling under nitrogen levels and root partition treatments
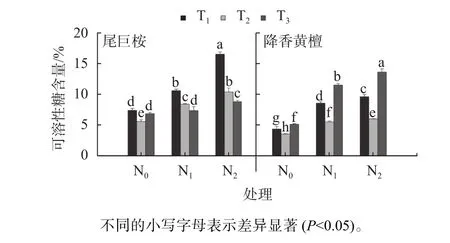
图6 施氮与根系分隔处理尾巨桉与降香黄檀SS含量差异Fig. 6 Difference in SS content of E.urophylla × E.grandis and D.odorifera under nitrogen levels and root partition treatments
2.6 尾巨桉与降香黄檀叶片保护酶活性变化
不同氮素水平与根系分隔模式对尾巨桉与降香黄檀叶片保护酶活性的影响见表4。由表4可知,尾巨桉与降香黄檀叶片SOD、POD以及CAT活性因氮素水平与根系分隔不同而差异显著(P<0.05),均表现为N2>N1>N0。不同根系分隔处理,尾巨桉和降香黄檀幼苗叶片保护酶活性变化与叶绿素含量、碳水化合物等指标变化基本一致,尾巨桉以T1优于T2和T3,降香黄檀则以T3最优,T1次之,T2最差。但根系分隔与施氮交互作用对保护酶活性的影响不显著,见表5。
2.7 氮素水平与根系分隔对幼苗干物质积累的影响
氮素是植物生长发育所必需的营养元素,施氮与根系分隔处理尾巨桉与降香黄檀干物质积累量差异见图7。
由图7可知,施氮显著提高尾巨桉与降香黄檀幼苗的干物质积累量 (P<0.05)。对比不同根系分隔处理,尾巨桉幼苗均为T1含量高于其余2种间作模式,其中N2水平的干物质积累量最高,比N0显著提高55.0% (P<0.05),比相同氮素水平的T2、T3间作模式分别提高了5.2%和13.3%;而同一施氮水平下,降香黄檀T3间作模式的干物质积累量显著高于T1和T2(P<0.05)。

表4 不同氮素水平与根系分隔模式对尾巨桉与降香黄檀叶片保护酶活性的影响Table 4 Effects of nitrogen levels and root partition treatments on activities of protective enzyme in E. urophylla × E.s grandis and D. odorifera leaves U/(min·g)
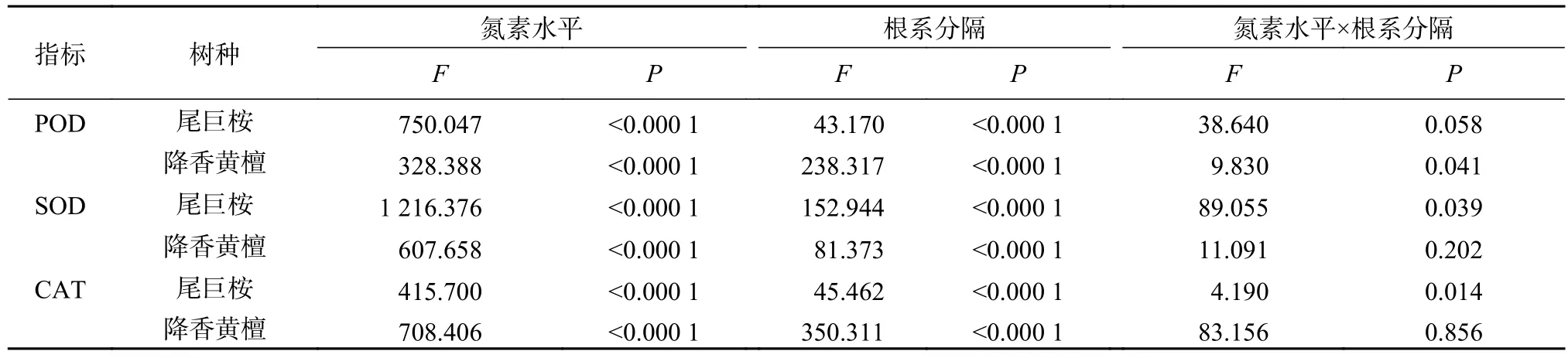
表5 根系分隔与施氮的条件下幼苗保护酶活性方差分析Table 5 Variance analysis of protective enzyme activity of seedlings under nitrogen levels and root partition treatments

图7 施氮与根系分隔处理尾巨桉与降香黄檀干物质积累量差异Fig. 7 Differences in dry matter accumulation of E. urophylla × E. grandis and D. odorifera under nitrogen levels and root partition treatments
3 结论与讨论
1) 氮素与根系分隔模式对叶面积与叶绿素含量的影响。叶片是植物进行生理代谢的重要器官,具有较强的可塑性和变异性,光合色素含量是植物光合作用的基础,叶绿素的减少会导致光合速率的降低[11],影响植物光合作用。氮素是叶绿素的主要组成成分,且大部分存在于叶绿体中[12],氮素含量的增加可促进叶绿素含量相应提高[13],如施氮可显著提高小麦 (Triticum aestirum)[14]、甘蓝 (Brassica oleracea)[15]、杨树 (Populussp.)[8]等不同植物叶绿素含量。本试验中,施氮显著提高了尾巨桉与降香黄檀叶面积指数和叶绿素含量,且随着氮素水平增加而增加,说明氮素可提高叶片质量和叶绿素含量。在不同根系分隔模式处理中,尾巨桉叶面积指数和叶绿素含量以T1最优,T2次之,T3最差,但降香黄檀以T3最好,T1次之,T2最差。出现这一现象可能是由于T1间作系统中,尾巨桉在竞争体系中处于优势地位,能更有效地吸收土壤养分,故而T1与T2体系叶面积指数均优于T3;其次是根系互作体系对尾巨桉具有促进作用,增加尾巨桉养分空间生态位,提高其对氮素的吸收,从而提高其叶面积、比叶重和叶绿素含量。降香黄檀则以T3最优,这可能是塑料膜隔消除尾巨桉与降香黄檀种间竞争,降香黄檀幼苗能更有效吸收土壤营养成分。但相比较T2,T1降香黄檀叶面积指数及其比重较大,推断可能是T1间作体系中根系互作对降香黄檀也存在一定促进作用,尾巨桉吸收大量养分,进一步刺激降香黄檀生物固氮,更大程度促进降香黄檀对养分的吸收与利用。根系互作中既相互竞争又相互协调的关系,尾巨桉在竞争作用中占优势地位,促进尾巨桉对养分的吸收。
2) 施氮与根系分隔模式对叶片保护酶活性的影响。植物生长过程中体内释放有害物质活性氧,破坏植物细胞结构与功能,造成植物细胞死亡,其代谢失衡引起植物衰老[16]。植物受胁迫时会形成相应的抗氧化酶保护系统,包括POD、CAT和SOD是植物保护系统的主要成分[17]。抗氧化酶POD、CAT的活性均具有清除植物体内歧化反应生成的H2O2的功能,与SOD协同作用维持体内活性氧代谢平衡,从而降低植物衰老进程[18]。根系分隔对尾巨桉与降香黄檀幼苗保护酶活性的影响有所差异,不同根系分隔模式,尾巨桉保护结构功能以 T1>T2>T3,降香黄檀则以 T3>T1>T2,出现不同变化规律。这可能是由于T1模式促进养分的吸收,增强其抗性。这与前人研究间作提高植物吸收养分,改变植物碳氮代谢,提高植物对环境胁迫的抗性结果一致[19]。
3) 施氮与根系分隔模式对叶片光能利用效率的影响。叶绿素荧光参数是植物利用光能主要参照指标,决定光合机构容量与性能[20],氮素缺乏明显降低了植物Y(II)、qP、qN和ETR,添加氮素将会刺激植物该指标的增加,提高尾巨桉与降香黄檀幼苗PS II反应中心的实际量子产量和提高光合相关电子传递能力,但降低了Y(NPQ)。与施氮对旱地小麦灌浆期叶绿素荧光参数影响时得出结论一致,施氮提高光能转换效率、qP和qN,提高光合电子传递能力[21]。有研究指出外源NO可以稳定干旱水稻PSII反应中心的结构和功能,增加其光能利用[22],CK对不同种植模式尾巨桉幼苗叶片叶绿素荧光参数表现无显著差异,N1、N2水平T1处理的Y(II)均显著高于T2与T3,但其他各处理间大部分无显著差异。这可能是由于施氮与T1间作协同作用显著增加了尾巨桉幼苗叶片叶绿素含量,从而促进叶片对电子光能传递与捕捉,提高植株光合作用。有研究指出豆科与非豆科作物间作时,存在豆科向非豆科氮素转移现象,促进非豆科作物生长生理特征[23]。本试验不同间作根系分隔模式中,尾巨桉以T1模式为最优,但降香黄檀则以T3最好,这可能是由于尾巨桉与降香黄檀根系互作时的竞争作用中尾巨桉处于优势地位,氮素由豆科树种向尾巨桉转移,提高尾巨桉氮素吸收量,通过增加叶片面积指数及叶绿素a含量等提高其对光能吸收与传递。
4) 施氮与根系分隔模式对叶片光合特性的影响。光合作用是植物生长的必需过程,是植物一系列代谢反应的总和。植物通过Pn、Tr、Gs、Ci和水分利用效率(WUE)等指标反映其光合特性[24]。光合作用受CO2浓度、干旱、光照强度、光质、温度、叶绿素含量、生长期盐碱度和营养元素等不同因子直接影响[25-30],尤其是氮素的缺乏对其具有较为直接性的影响。前人研究认为,施氮增加降香黄檀容器苗[31]、雷竹(Phyllostachys preacox)幼苗[32]以及巨桉(Eucalyptus grandis)[33]等Pn、Tr和Gs,但降低Ci。本试验研究得出结论与其一致,施氮可提高尾巨桉与降香黄檀Pn、Gs、Tr,降低叶片Ci。但也有研究指出,米槁(Cinnamomum migao)Ci随着氮素水平增加而增大[34],本实验结果与其相反,可能是由于本实验施氮增加Gs,促进CO2进入细胞内供给光合作用,降低了Ci。豆科与非豆科植物间作处理时2个物种变化趋势一致,如玉米 (zea mays) 与花生 (Arachis hypogaea) 间作光合特性影响的研究表明间作光合指标高于单植[35],研究玉米与苜蓿 (Medicago sativa) 间作则认为间作降低玉米及苜蓿净光合速率[36],本研究与前人结果有所不一致。尾巨桉与降香黄檀间作处理中各光合特性各指标与其相关生理指标变化一致,互作促进了尾巨桉的光合作用,但对降香黄檀起抑制作用。这也说明植物叶片生理代谢与光合能力有较强相关性[37]。出现2种植物光合作用变化趋势不一样的现象可能存在2种机制,机制一:豆科植物向非豆科植物转移氮素,促进非豆科植物的生长,同时刺激豆科植物提高生物固氮效率;机制二:尾巨桉与降香黄檀根系互作导致尾巨桉分泌一些未知物质,抑制降香黄檀的生长。尾巨桉在间植系统中可能响应机制一,但降香黄檀可能同时响应机制一与机制二。T1与T2间作氮素发生氮素转移促进尾巨桉的生理代谢与光合作用,T3消减氮素转移现象,利于降香黄檀吸收土壤氮素营养,提高生理代谢,增强光合作用,同时尾巨桉分泌的某些物质透过尼龙网抑制降香黄檀生长,导致T2处理生长较差,抑制其光合作用。
5) 施氮与根系分隔模式对叶片SS含量的影响。SS是植物的主要营养物质,是植物体内大分子物质的基础,是光合作用的产物,也是碳水化合物代谢和贮藏的场所,在碳代谢过程发挥重要的作用[16]。叶片中的SS含量主要用于满足自身的生长消耗,过剩的则运输到根茎中以淀粉形式储存[8]。本试验得出施氮显著提尾巨桉与降香黄檀幼苗叶片SS含量,与前人研究结果基本一致。表明施氮能够刺激植株的细胞生理功能,达到一定活跃值从而促进植株的增长,增加植物生理代谢。不同根系分隔模式处理,尾巨桉与降香黄檀SS含量与叶绿素含量及光合作用表现基本一致。原因可能是SS是光合作用产物,其含量与光合作用强弱具有正向关系,尾巨桉不隔间作光合能力强于其他2种根系分隔模式,故SS含量较多,而降香黄檀则是塑料膜隔无竞争作用处理含量最高。这可能是根系互作对地下部分养分生态位的扩展具有一定的作用,促进竞争能力较强的尾巨桉生理代谢与光合作用,抑制了降香黄檀。
通过盆栽试验发现,施氮有益于尾巨桉和降香黄檀生理代谢和光合作用的增强,但由于尾巨桉在间作体系中占优势地位,所以根系分隔处理对尾巨桉和降香黄檀生理生长的影响不一致。在根系互作的情况下,尾巨桉的干物质积累显著高于单植处理。由试验结果得出高氮水平不隔间作模式最佳,即合理施氮与根系互作同时存在对尾巨桉的生理生长方面具有较强的促进作用,引入具有生物固氮的树种进行混交是解决桉树多代连栽引起环境问题的有效途径。
