外切型琼胶酶AgaO的 高效表达及酶学性质表征
2019-04-12,,,,,,*
,,,,,,*
(1.山东农业工程学院食品科学与工程学院,山东省教育厅特色 农产品采后品控与综合利用重点实验室,山东济南 250100; 2.山东大学国家糖工程技术研究中心,糖化学与生物学山东省重点实验室,山东青岛 266237)
琼胶主要产自江蓠、紫菜等食用红藻,与褐藻胶、卡拉胶并称产量最大、用途最广的海洋三大多糖,广泛应用于食品、医药和化工行业。琼脂糖是琼胶的主要成分之一,属中性多糖,分子主链由二糖单元(3-6-内醚-L-吡喃半乳糖-α-1-3-吡喃半乳糖)通过β-1-4糖苷键重复交替链接而成,分子内存在α-1-3和β-1-4两种糖苷键类型[1]。
琼胶酶催化琼脂糖中糖苷键的水解,产生水溶性寡糖产物。因催化位点处糖苷键的类型不同,分为α-琼胶酶、β-琼胶酶[2]。这两种酶的寡糖产物分别以3,6-内醚-α-L-吡喃半乳糖(A)、β-D-吡喃半乳糖(G)为还原性末端,称之为琼寡糖(Agaro-oligosaccharides,AOs)、新琼寡糖(Neoagaro-oligosaccharides,NAOs)[3-4]。琼胶酶可用于DNA片段的琼脂糖凝胶回收以及海藻原生质体制备[5-6]。琼脂糖的降解产物-低聚糖具有抗炎症[7]、抗氧化[8]、预防糖尿病[9]、促进肠道有益菌群生长[10],以及通过抑制黑色素分泌促进皮肤增白[11-12]等重要活性,且未见毒副作用,表明其比琼脂糖本身具有更大应用潜力与更高经济价值。
目前,限制琼胶寡糖的商品化生产与广泛应用的主要因素之一是制备方法[13]。化学法虽然工艺简单、成本低,但反应条件剧烈、不易控制,且寡糖产物的分子量不均一,分布范围广。与化学法降解相比,琼脂糖的酶法降解具有条件温和可控、污染小且能耗低等绿色环保、高效的特征[14]。因此,深入挖掘产量大、活性高、稳定性强的工具型琼胶酶资源对于实现琼胶寡糖的商品化生产至关重要。课题组前期从海泥样品中分离到一株琼胶液化能力强的多糖降解菌-火色杆菌Flammeovirgasp. MY04,其基因组编码至少15个候选琼胶酶,可归类于糖基水解酶的GH16、GH50或GH86家族,无GH96或GH118家族成员[15]。候选琼胶酶AgaO在基因组编码产物中无同源蛋白,系统发育分析表明它与GH50家族的β-琼胶酶聚类,但序列相似性低于40%[16]。BLASTp在线分析表明,AgaO含有曾定义为GH50家族、现为GH42家族的催化模块,其催化位点残基包括Gln458和Gln606等高保守位点。然而,重组酶rAgaO与GH42家族成员不同,无糖苷酶活性;与GH50家族成员相似,是琼脂糖外切酶,在降解底物糖链时专一生产新琼二糖[16]。因此,琼胶酶AgaO是MY04菌株降解利用琼脂(多)糖的关键限速酶,且可用于新琼二糖的专一性制备。遗憾的是rAgaO的产量较低,易于形成包涵体。因此,为提高外切型琼胶酶AgaO的表达效率,本研究优化了AgaO的基因密码子、异源诱导表达条件,并分析了水溶性重组酶的酶学性质及变化,以期获得工具酶的高效生产工艺。
1 材料与方法
1.1 材料与仪器
大肠杆菌DH5α、大肠杆菌BL21(DE3)、大肠杆菌Trans-T1、大肠杆菌表达载体pET-30a(+) 实验室保存;LB液体培养基(pH7.0)、LB固体培养基(琼脂粉15 g/L) 实验室自配;DNA凝胶回收试剂盒、质粒提取试剂盒 天根生化科技(北京)有限公司;限制性内切酶、T4 DNA连接酶、Prime STAR Max DNA聚合酶 宝生物工程(大连)有限公司;载体pEasy Blunt Cloning Vector、DNA Marker 北京全式金生物科技有限公司;引物、卡那霉素、BCA蛋白测定试剂盒 上海生工生物工程股份有限公司;琼脂糖 美国Invitrogen公司;Ni-NTA琼脂糖凝胶 美国Novagen公司;硅胶板 德国Merck公司;其他试剂均为国产或进口分析纯。
DYCP-31DN型核酸电泳仪、DYCZ-24DN型蛋白电泳仪 北京六一仪器厂;Mastercycler nexus梯度PCR仪、5810R高速冷冻离心机 德国Eppendorf公司;GelDoc XR+凝胶成像仪 美国Bio-Rad公司;DU800分光光度计 美国Beckman公司;-80 ℃超低温冰箱 日本Sanyo公司;HYG-C恒温培养振荡器 江苏太仓市实验设备厂。
1.2 实验方法
1.2.1 EAgaO密码子的优化和人工合成 火色杆菌Flammeovirgasp. MY04(专利保藏号CGMCC 2777)是课题组分离自海泥样品的1株海洋细菌,可降解多种多糖,能直接液化琼脂糖并产生系列新琼寡糖[15]。通过新一代基因组测序技术测定了MY04全基因组序列(GenBank登录号:CP003560-CP003562),并进行了初步注释,发现存在1个单独聚类的候选琼胶酶AgaO。使用软件BioeEdit7.2.1和SignalP4.0对Flammeovirgasp. MY04琼胶酶基因agaO序列(GenBank登录号:KU524066)进行序列分析[17-18]。首先除去5′端75 bp信号肽编码序列,然后参照大肠杆菌菌株的密码子使用频率[19],将该基因的密码子替换为在大肠杆菌中高频使用的密码子,结合考虑(G+C)%含量、密码子适应指数等因素进行综合优化[19-20],并委托上海Invitrogen公司进行全基因合成,优化后的基因命名为eagaO。
1.2.2 重组表达载体pET30a-EAgaO的构建及鉴定 根据eagaO的基因序列设计引物并由生工生物工程(上海)股份有限公司合成,引物序列为:eagaO-F:gccgCATATGAATCCGCCTAAAAGCATCGATG(划线部分为添加的NdeI酶切位点),eagaO-R:gccgCTCGAGATTATTCACATTGCCCAGACG(划线部分为添加的XhoI酶切位点)。PCR扩增后得到基因片段eagaO,连接pEasy Blunt Cloning Vector,转化Trans-T1感受态细胞,菌液PCR筛选阳性克隆子。将阳性克隆子和pET-30a(+)分别用NdeI和XhoI进行双酶切,并经DNA凝胶回收试剂盒回收纯化后,T4 DNA 连接酶16 ℃过夜连接,转化DH5α感受态细胞,在含有50 μg/mL卡那霉素的LB平板上筛选转化子,并通过双酶切及测序对重组质粒进行检测及鉴定。构建的重组质粒命名为pET30a-EAgaO。
1.2.3 琼胶酶rEAgaO的诱导表达 将重组质粒pET30a-EAgaO转化E.coliBL21(DE3)宿主细胞。挑取转化后的单克隆至液体LB培养基(50 μg/mL卡那霉素)中,37 ℃、200 r/min培养6~8 h。按1%(V/V)的接种量转接至含100 mL新鲜LB培养基的500 mL三角瓶中,37 ℃,200 r/min 继续振荡培养,至OD600达到0.6~0.8。向培养基中添加一定量的诱导剂IPTG,在一定温度下诱导培养一段时间,10000×g、4 ℃离心10 min收集菌体,用Buffer A(50 mmol/L Tris、150 mmol/L NaCl,pH8.0)洗涤沉淀后,重悬于Buffer A后进行超声破碎,破碎液15000×g、4 ℃离心30 min,二次离心后所得上清即为粗酶液。用BradFord法[21]测定蛋白质的浓度。
1.2.4 重组酶表达条件的单因素优化
1.2.4.1 诱导时间对重组酶诱导表达的影响 采用1.2.3的诱导表达方法,首先使菌体培养至OD600达到0.6~0.8后,添加终浓度为0.1 mmol/L IPTG作为诱导剂,16 ℃分别诱导培养4、8、12、16、20、24 h(不包含加入IPTG前的培养时间)提取粗酶液。测定酶活力大小,以最高酶活力为100%,通过分析计算不同诱导时间下的相对酶活(%),比较分析确定最佳诱导表达时间。
1.2.4.2 诱导温度对重组酶诱导表达的影响 菌体首先在37 ℃培养OD600达到0.6~0.8后,添加终浓度为0.1 mmol/L IPTG作为诱导剂,然后分别在12、16、20、25、30、35 ℃下继续诱导培养20 h后提取粗酶液。测定酶活力大小,以最高酶活力为100%,通过分析计算不同诱导温度下的相对酶活(%),分析确定最佳诱导温度。
1.2.4.3 诱导剂IPTG终浓度对重组酶诱导表达的影响 菌体培养至OD600达到0.6~0.8后,在16 ℃下,分别添加终浓度为0、0.05、0.1、0.3、0.5、0.7、1.0 mmol/L IPTG,继续诱导培养20 h后提取粗酶液。测定酶活力大小,以最高酶活力为100%,通过分析计算不同诱导剂浓度下的相对酶活(%),分析确定最佳诱导剂IPTG终浓度。
1.2.5 琼胶酶rEAgaO的纯化 按照优化后的方案诱导表达重组蛋白,10000×g、4 ℃离心10 min收集菌体,用Buffer A洗涤后,重悬于Buffer A后进行超声破碎,破碎液15000×g、4 ℃离心10 min取上清获得粗酶液。
由于重组质粒pET30a-EAgaO诱导表达琼胶酶的N端融合有His标签,本实验利用Ni-NTA琼脂糖凝胶对该融合酶进行亲和纯化。具体步骤如下:取1 mL填料置于10 mL柱子中,待完全沉淀后,先用10 mL ddH2O洗涤,然后加入10 mL Buffer A进行平衡;将含有约50 mg总蛋白的酶液加入填料中,轻轻混匀,16 ℃结合4 h,在重力作用下小心流出液体;随后分别用1 mL含有咪唑浓度为10、50、100、250、500 mmol/L的缓冲液A进行梯度洗脱,优化洗脱液咪唑浓度条件,EP管分别收集洗脱液并标记。
用BradFord法[21]测定蛋白质的浓度。用分离胶浓度为13.2%的SDS-PAGE检测蛋白质的纯度。灰度扫描并进行面积积分,分析重组蛋白质的相对含量。
将上述经检测含有单一蛋白条带的样品液移于透析袋(截留分子量14 kDa),放置于Buffer B(25 mmol/L Tris,5%甘油,pH8.0)中,4 ℃透析连续透析4次,获得纯酶液以备后续研究。
1.2.6 酶活测定 配制0.1%(w/v)的琼脂糖溶液作为底物,向每500 μL底物中添加等体积的酶的稀释液,混合后水浴反应12 h。采用3,5-二硝基水杨酸(3,5-Dinitrosalicylic acid,DNS)法[22]测定体系中的还原糖量。酶活力单位(U)定义:每分钟生成1 μmol还原糖所需要的酶量为1 U。
1.2.7 琼胶酶rEAgaO的酶学性质分析
1.2.7.1 酶的最适温度及热稳定性测定 用100 mmol/L NaH2PO4-Na2HPO4(PBS,pH7.0)缓冲液配制0.1%的琼脂糖,吸取500 μL作为底物,加入500 μL适当稀释的纯酶液(对照组为沸水灭活酶液)分别在0、10、20、30、35、40、45、50、60、70 ℃不同温度下反应12 h后,沸水浴10 min终止反应,测定酶活力,以最高酶活力为100%,确定酶的最适反应温度。
将酶液分别在40、45、50、60、70 ℃不同温度下预先保温0、30、60、90、120 min后,以不经过预处理的酶液酶活定义为100%,测定酶的残余活力,分析酶的温度稳定性。
1.2.7.2 酶的最适pH及pH稳定性测定 分别用浓度为100 mmol/L的NaAc-HAc 缓冲液(pH5、6)、100 mmol/L的NaH2PO4-Na2HPO4(PBS)缓冲液(pH6.0、7.0、8.0)、100 mmol/L的Tris-HCl 缓冲液(pH8.0、9.0、10.0),与琼脂糖配制终浓度为0.10%的琼脂糖底物,吸取500 μL作为底物,加入500 μL适当稀释的纯酶液在45 ℃下反应12 h。在上述不同反应体系中测定酶的活力,以最高酶活力为100%,确定酶的最适反应pH。
将适当稀释的纯酶液分别置于50 mmol/L的NaAc-HAc 缓冲液(pH5、6)、50 mmol/L的NaH2PO4-Na2HPO4(PBS)缓冲液(pH6.0、7.0、8.0)、50 mmol/L的Tris-HCl缓冲液(pH8.0、9.0、10.0)不同pH缓冲体系中,4 ℃预先放置2 h后,然后在pH7.0、45 ℃条件下反应12 h,以未经预处理的酶液的酶活定义为100%,测定酶的残余活力,分析酶的pH稳定性。
1.2.7.3 金属离子和化学试剂对酶活力影响的测定 向纯化后的琼胶酶反应体系中添加AgNO3、KCl、LiCl、NaCl、CaCl2、CoCl2、CuSO4、FeSO4、HgCl2、MgSO4、MnSO4、NiSO4、PbAc2、ZnSO4、CrCl3、FeCl3、SDS、Na2EDTA、甘油、β-巯基乙醇、咪唑及DTT不同的金属离子和化学试剂,添加的离子终浓度分别为1、10 mmol/L,然后在pH7.0、45 ℃条件下反应12 h测定酶的活力,以不添加任何金属离子和化学试剂的对照组酶活力为100%,分析金属离子和化学试剂对酶活性的影响。
1.2.8 降解产物的分析 按照体积比1∶1混匀琼脂糖底物和1.2.5制备的纯酶液,45 ℃保温反应。隔时取样,沸水浴10 min使酶失活,离心收集产物,参考薄板层析法[23]使用硅胶板进行TLC分析。展开剂及各组分体积比为正丁醇∶乙醇∶水=2∶1∶1,显色剂为苯胺∶二苯胺∶磷酸∶丙酮=1∶1∶5∶50 (mL∶g∶mL∶mL)。染色后110 ℃加热10 min使其显色。所用含有新琼二糖(Neoagarobiose,NA2)、新琼四糖(Neoagarotetraose,NA4)和新琼六糖(Neoagarohexaose,NA6)的分子量标准物是用β-琼胶酶降解琼脂糖并纯化后的产物,由实验室自主制备[16,24]。
1.3 数据处理
所有实验进行3次重复,取其平均值,平均值和标准差的统计分析由Microsoft Excel 2007软件进行处理。
2 结果与分析
2.1 琼胶酶基因eagaO重组表达载体的构建与验证
首先对密码子进行了优化,优化后的eagaO基因(GenBank收录号:MG708345)GC含量由35.5%提升至42.5%。以人工合成的eagaO基因片段为模板,使用引物对eagaO-F/eagaO-R经PCR扩增后连接到载体pEasy Blunt Cloning Vector,命名为pEB-EAgaO(表1)。重组质粒pEB-EAgaO和表达质粒pET-30a(+)同时经NdeI/XhoI双酶切并连接后,获得重组琼胶酶rEAgaO的表达载体pET30a-EAgaO(表1)。该质粒的双酶切验证结果显示:插入的目的片段大小约2100 bp(图1),与理论分子量一致。

表1 本实验所使用的重组质粒性质和来源Table 1 Description and source of the recombinant plasmids used in the present study

图1 重组质粒pET30a-EAgaO双酶切验证Fig.1 Identification of the recombinant vector pET30a-EagaO by double restriction enzyme digestion注:M:Trans2K Plus II DNA Marker;1:重组质粒 pET30a-EAgaO经NdeI/XhoI双酶切的产物; 2:质粒pET-30a(+)经NdeI/XhoI双酶切的产物。
2.2 重组酶表达条件的单因素优化
2.2.1 诱导时间对诱导产酶酶活的影响 如图2所示,当添加诱导剂后继续表达的时间为4~20 h时,随时间延长,粗酶液的酶活增加。20、24 h时所获的酶活力达到最高值,且无明显变化。在20 h内,细胞的密度和细胞内的水溶性产物都是不断增加的,在20 h后,由于产物在宿主中已经大量积累,导致新产物主要形成包涵体而非水溶性蛋白[25]。因此,从节约成本和时间的角度考虑,选取20 h为最佳诱导时间。

图2 诱导时间对诱导产酶酶活的影响Fig.2 Effects of indution time on the production of enzyme activity
2.2.2 诱导温度对诱导产酶酶活的影响 如图3所示,温度在12~16 ℃时,粗酶液的酶活力逐渐升高;当温度为16 ℃时获得最大酶活力。当高于16 ℃时,发酵液产物酶活力逐渐降低,这是由于较高的温度可能会导致产物酶活力降低。则16 ℃为琼胶酶rEAgaO的最适诱导表达温度。

图3 诱导温度对诱导产酶酶活的影响Fig.3 Effects of indution temperatures on the production of enzyme activity
2.2.3 IPTG浓度对诱导产酶酶活的影响 如图4所示,在IPTG终浓度为0.1~0.7 mmol/L时所得粗酶液的活力没有明显差异;所添加IPTG终浓度继续加大后,粗酶液的活力开始下降。IPTG自身对大肠杆菌有一定的毒性,因此IPTG高于一定浓度后,表现出生长抑制作用而导致产酶量下降。从节约成本的角度考虑,选择IPTG终浓度为0.1 mmol/L时为最佳诱导剂添加浓度。
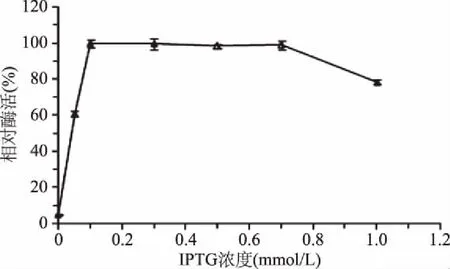
图4 IPTG浓度对诱导产酶酶活的影响Fig.4 Effects of IPTG concentrations on the production of enzyme activity
2.3 琼胶酶rEAgaO的分离纯化
SDS-PAGE检测分析初步表明,在最优条件下重组酶rEAgaO在大肠杆菌内实现了高效表达。灰度扫描和面积积分表明,在可比条件下,水溶性组分中重组蛋白的含量是水不溶组分中含量的19.2倍,表明95%以上重组蛋白为水溶性表达,这比原始基因表达产物中的含量(30%[16])提高了2.2倍。结果表明,当洗脱液中咪唑浓度为250 mmol/L时,可洗脱得到单一的蛋白条带。如图5所示,纯化后的目的蛋白大小约为81.3 kDa,分子量与目标蛋白理论分子量相符。根据菌体水溶性蛋白质的总浓度及目标蛋白含量推算,rEAgaO的发酵产量约为1432.7 mg/L,这比rAgaO的发酵产量(0.12 g/L[16])提高了10.9倍。纯化所得重组酶的纯度>95%,得率约为85.7%,活力为910.2 U/mg,比rAgaO的酶活(185 U/mg)[16]提高了近4倍。

图5 琼胶酶rEAgaO表达和纯化的SDS-PAGE检测Fig.5 SDS-PAGE analysis of expression and purification of agarase rEAgaO
2.4 琼胶酶rEAgaO的酶学性质分析
2.4.1 温度对rEAgaO活性及稳定性的影响 温度会影响酶蛋白分子的稳定性,温度过低,酶活性降低,温度过高会使酶蛋白变性失活。如图6所示,琼胶酶rEAgaO在45 ℃反应时具有最高的相对酶活,表明rEAgaO的最适反应温度是45 ℃,明显高于琼胶的凝固温度(40 ℃),有利于酶解反应的进行,降低了反应成本。如图7所示,rEAgaO酶活性在40、45、50、60 ℃具有较好的稳定性,保温2 h,酶活仍保持66%以上(分别为90.4%、83.5%、77.9%、66.3%)。70 ℃酶活急剧下降,保温1 h酶活仅为最高值的5.2%,至1.5 h已检测不到酶活。说明该酶具有一定的热稳定性。
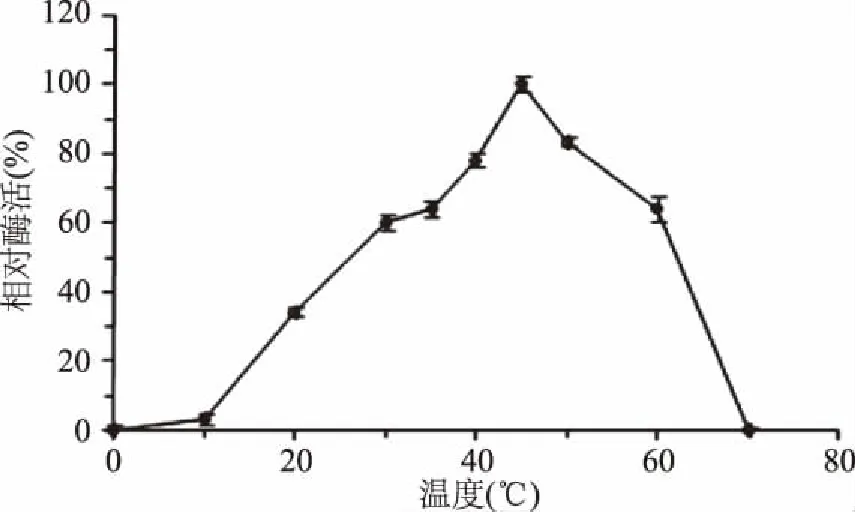
图6 温度对琼胶酶rEAgaO活性的影响Fig.6 Effects of temperature on agarase rEAgaO activity

图7 温度对琼胶酶rEAgaO热稳定性的影响Fig.7 Effects of temperature on thermal stability of agarase rEAgaO
2.4.2 pH对rEAgaO活性及稳定性的影响 pH会影响酶蛋白分子与底物的结合,过酸或者过碱都会使酶变性失活。如图8所示,以琼脂糖为底物测定酶活时,随着pH的升高,琼胶酶rEAgaO的酶活力呈现先升高后降低的趋势,pH7.0时,相对酶活力最高。在pH6.0~10.0范围内4 ℃处理2 h,rEAgaO的残余酶活在69.5%以上,显示该酶具有广泛的pH稳定性(图9)。
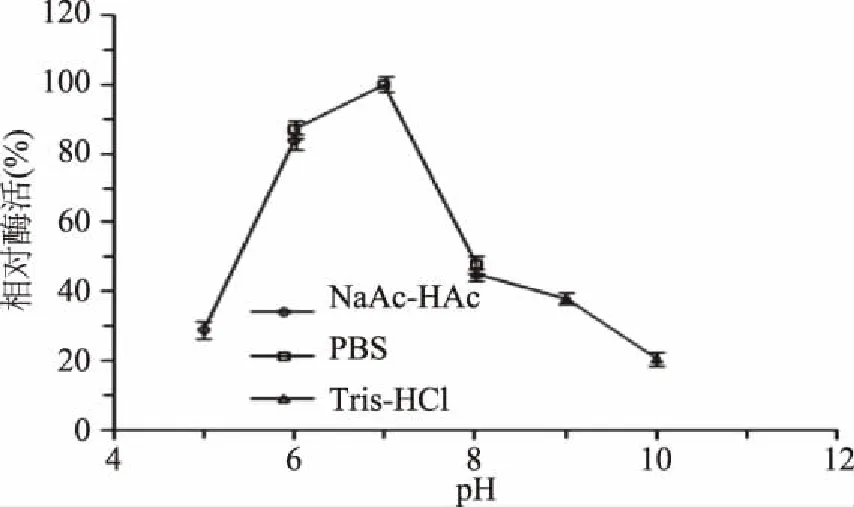
图8 pH对琼胶酶rEAgaO活性的影响Fig.8 Effects of pH on agarase rEAgaO activity

图9 琼胶酶rEAgaO pH稳定性Fig.9 pH stability of agarase rEAgaO
2.4.3 金属离子及化学试剂对rEAgaO活性的影响 不同的金属离子和化学试剂对琼胶酶rEAgaO的酶活力影响不同,如图10所示,实验的大多数金属离子对rEAgaO都有不同程度的抑制作用,例如Ag+、Co2+、Cr3+;化学试剂SDS、EDTA、咪唑、β-巯基乙醇及DTT也都对rEAgaO的酶活具有不同程度的抑制作用;其他金属离子,如Na+、K+、Ca2+等在海水中含量丰富的离子,对该酶的活性影响均不大,可能与该酶产生菌来源于海洋生境有关;相比之下,10 mmol/L的Mg2+对rEAgaO的活性有明显促进作用,可使酶活提高至118.8%。
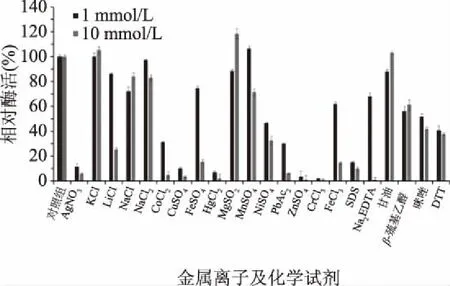
图10 金属离子和化学试剂对琼胶酶rEAgaO活性的影响Fig.10 Effects of metalic ions and chemicals on agarase rEAgaO activity
2.5 降解产物的分析
聚合度相同的新琼寡糖和琼寡糖在TLC分析中的展开系数Rf明显不同。如图11结果显示新琼寡糖的带型清晰,测得新琼寡糖标准物中新琼二糖(NA2)、新琼四糖(NA4)和新琼六糖(NA6)的Rf值分别为0.57、0.42和0.28。将琼胶酶rEAgaO和琼脂糖底物反应12、24 h,TLC分析降解产物中仅含有与新琼二糖NA2 Rf值一致的寡糖。这表明,与rAgaO相似,rEAgaO仍然是以NA2为主产物的外切型琼胶酶[16]。

图11 琼胶酶rEAgao降解琼脂糖的产物分析Fig.11 Analysis of the hydrolysis products of agarose by agarase rEAgaO
3 结论
以大肠杆菌为宿主的原核表达系统,因遗传背景清晰、操作简单、生产成本低等优点备受青睐。目前,已有多个不同物种来源的琼胶酶基因在大肠杆菌中成功表达[24,26-27],但却少见高效表达与商品化生产。本研究参照大肠杆菌的密码子使用频率,用高频密码子将火色杆菌Flammeovirgasp. MY04来源外切型琼胶酶基因agaO进行优化,并将GC含量由35.5%提升至42.5%,最终人工合成获得基因片段eagaO。所构建的表达载体,去除了野生型信号肽的编码序列及载体所固有的信号肽编码序列。经过蛋白质表达、纯化与性质鉴定、生化特征分析表明,与用原始基因agaO构建的载体pBAgaO所表达的重组酶rAgaO相比[16],rEAgaO的最适温度(45 ℃)和最适pH(7.0)没有变化。但是,可比条件下,rAgaO在0~40 ℃环境中预处理2 h,表现出热稳定性[16],而密码子改造后的rEAgaO在更宽的温度范围(0~60 ℃)表现出热稳定性(图7);rAgaO在pH6.0~8.0的环境中预处理2 h,表现出pH稳定性[16],而rEAgaO在更宽的pH范围(6.0~10.0)内表现出稳定性(图9)。而酶学性质分析表明:在最适条件下,rAgaO的酶活是185 U/mg[16],而rEAgaO的酶活是910.2 U/mg,比rAgaO提高了近4倍。与rAgaO相同,rEAgaO降解琼脂糖时的主产物始终是新琼二糖(图11)。研究者认为,尽管最适温度、最适pH、寡糖生成特性等特征没有因为基因密码子的改造而发生变化,但是水溶性重组酶的产量与酶活大幅提高,使得应用密码子优化后基因片段来高效表达、商品化生产外切型琼胶酶rEAgaO成为可能。这更有利于二糖外切型琼胶酶rEAgaO作为一种工具酶进行推广应用。
