HtrA对乳牙变异链球菌产酸、黏附的体外研究
2019-04-06韩林秀刘兴容
王 玲,刘 瑶,韩林秀,刘兴容
(西南医科大学:1口颌面修复重建与再生实验室,2附属口腔医院儿童口腔科,四川泸州 646000;3四川省交通运输厅公路局医院口腔科)
乳牙龋是儿童最常见的口腔疾病,第四次全国口腔健康流行病学调查结果显示5岁儿童乳牙龋患率为70.9%,其中经过充填治疗的牙齿比例仅为4.1%[1],可见我国儿童乳牙存在患龋率高而就诊率低的状况。多项研究[2-4]证实,儿童唾液中变异链球菌浓度与乳牙龋坏严重程度呈正相关性。其致龋性主要毒力因子包括:产酸耐酸能力、在牙面定植形成生物膜的能力、合成胞内及胞外多糖的能力等[5-6]。丝氨酸蛋白酶是变异链球菌的致龋关键酶,由HtrA基因编码的HtrA蛋白作为一种热休克蛋白,属于丝氨酸蛋白酶家族重要成员。前期实验已筛选出的乳牙变异链球菌HtrA 高毒力株的致龋性高于其他临床分离株[7],同时利用分子生物学技术构建的HtrA 缺陷株的转化力减低,进而使其毒力出现了下降[8]。本次研究目的拟在前期实验基础上,比较HtrA高毒力株与HtrA 缺陷株产酸能力、体外黏附能力的差异,为HtrA 基因对乳牙变异链球菌致龋性调控研究提供一定实验依据。
1 材料与方法
1.1 材料
1.1.1 实验菌株
乳牙变异链球菌HtrA 高毒力株(前期课题组筛选完成);乳牙变异链球菌HtrA 缺陷株(前期课题组构建完成);变异链球菌国际标准株UA159(简称UA159标准株)(四川大学华西口腔医学院国家级重点实验室提供)。
1.1.2 主要试剂
BHI液体培养基、BHI固体培养基(北京Solarbio科技有限公司)、羟基磷灰石粉末(上海迈坤化工有限公司)、荧光染料2',7'-bis-(2-carboxyethyl)-5-(6)-carboxyfluorescein,acetoxymethyl Ester(BCECF/AM,上海碧云天生物技术有限公司)、NaOH粉末(四川西陇化工有限公司)。
1.1.3 主要仪器
超净工作台(苏州净化设备有限公司)、荧光酶标仪、Thermo Multiskan Go全波长酶标仪(美国Thermo 赛默飞公司)、雷磁PHSH-3F 型精密酸度计(上海仪电科学仪器股份有限公司)、电热恒温鼓风干燥箱(上海齐欣科学仪器有限公司)。
1.2 方法
1.2.1 细菌的复苏、培养及鉴定
将冻存的乳牙变异链球菌HtrA 高毒力株和HtrA缺陷株复苏,经革兰染色形态学及生化鉴定为纯培养后,分别接种BHI琼脂培养基中,37 ℃恒温厌氧培养48 h,挑取单菌落接种于BHI液体培养基中,37 ℃厌氧培养24 h 增菌,酶标仪调节菌液吸光度OD600=1.0,备用。
1.2.2 生长曲线测定
将HtrA 高毒力株和HtrA 缺陷株的菌液(OD600=1.0)与BHI 液体培养基分别按总体积比为5%:95%的比例加入试管内,每组18管,37 ℃厌氧培养16 h,期间每隔2 h从两组中各取2管菌液放置于4 ℃冰箱保存,直至16 h。各样本震荡混匀后各取200 uL 加入96 孔板,每管样本取2 次,酶标仪测OD600值并做好记录。
1.2.3 产酸能力测定
将HtrA高毒力株和HtrA缺陷株的菌液(OD600=1.0)与pH4.5~7.0(0.5 为间隔)的BHI 液体培养基分别以体积比为1:10的比例接种,每组菌株相同pH设置三份,37 ℃厌氧培养48 h,3 000 r/min 离心15 min,移取上清液,酸度计测定上清液终末pH 值,取三组均值,计算ΔpH(初始pH-终末pH)。实验重复两次。
1.2.4 黏附能力测定
1.2.4.1 细菌悬液的标记
分别取500 μL 菌液(OD600=1.0)HtrA 高毒力株和HtrA缺陷株的菌液于试管中,再分别加入1.5 mL PBS 缓冲液和3 μL 荧光染料BCECF/AM,放入37 ℃恒温厌氧培养2 h,进行荧光标记。标记完成后,3000 r/min 离心5 min,PBS 洗涤。将洗涤好的两组细菌分别重悬于37 ℃的PBS缓冲液中,酶标仪调节菌液OD600=1.0,分装于96孔板中,备用。
1.2.4.2 唾液包被的羟基磷灰石的制备
量取50 mg 羟基磷灰石粉末,悬浮于10 mL 0.2 mol/L的NaOH溶液中,处理10 min,超纯水洗涤直到上清液pH接近7.0,静置使粉末沉淀,小心吸弃上清液,沉淀放入烘箱60 ℃过夜干燥。收集同一健康志愿者进食4 h后的无刺激性唾液30 mL,12 000 r/min离心20 min,收集上清液,无菌微孔滤膜过滤除菌。按照羟基磷灰石粉末:唾液=1 mg:200 μL的比例混匀,37 ℃恒温放置1 h,2000 r/min离心5 min,移除上清液,沉淀即包被完成的羟基磷灰石。
1.2.4.3 黏附实验
在96 孔板中加入包被完成的羟基磷灰石粉末1 mg,标记后的菌株200 μL,混合均匀。其中HtrA高毒力株和HtrA 缺陷株各3 个孔。37 ℃恒温厌氧培养30 min。小心吸弃上清液,PBS 缓冲液轻柔洗涤沉淀1 次,PBS 缓冲液使沉淀重新悬浮,荧光酶标仪分别测定两组细菌荧光值,即为黏附菌的荧光值。将只加入羟基磷灰石粉末和PBS缓冲液的相应对照孔作为阴性对照,将未加入羟基磷灰石粉末,只含标记菌液的相应对照组荧光值作为总的荧光值。在荧光酶标仪上设置激发波长为485 nm、发射波长为535 nm,测定各孔的荧光值,以减去阴性对照后的值作为测定的荧光值。
黏附率公式如下:
细菌黏附率=(黏附细菌的荧光值-阴性对照荧光值)/(总的荧光值-阴性对照荧光值)×100%
1.3 统计分析
2 结果
2.1 生长曲线测定
两组菌株革兰氏染色油镜下(×1 000)形态分别如图1、2 所示。在非应激环境下,两组菌株在0~4 h 的生长趋势和生长速度基本一致(P>0.05),在4 h 及以后,HtrA 高毒力株的菌液浓度均高于HtrA缺陷株,其差异有统计学意义(P<0.05),见图3。

图1 HtrA高毒力株(×1 000)

图2 HtrA缺陷株(×1 000)
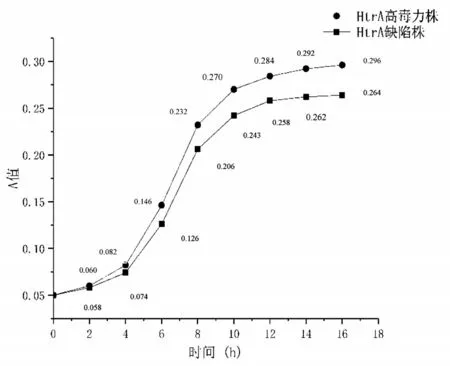
图3 两菌株生长曲线的比较
2.2 产酸能力测定
在pH 为6.5 和7.0 时,两菌株均能产酸,产酸能力没有明显不同,其差异无统计学意义(P>0.05);在pH 值6.0、5.5、5.0 时,HtrA 缺陷株产酸能力弱于HtrA 高毒力株,其差异有统计学意义(P<0.05);在pH值4.5时,HtrA缺陷株和HtrA高毒力株的产酸能力均受到明显抑制,几乎不产酸,其差异不具统计学意义(P>0.05),见表1。
2.3 黏附实验测定结果
变异链球菌HtrA 缺陷株在唾液包被的羟基磷灰石表面的黏附率显著低于HtrA 高毒力株(P<0.05),见图4。
表1 不同pH值下HtrA缺陷株和HtrA高毒力株的产酸能力的比较(±s,n=6)

表1 不同pH值下HtrA缺陷株和HtrA高毒力株的产酸能力的比较(±s,n=6)
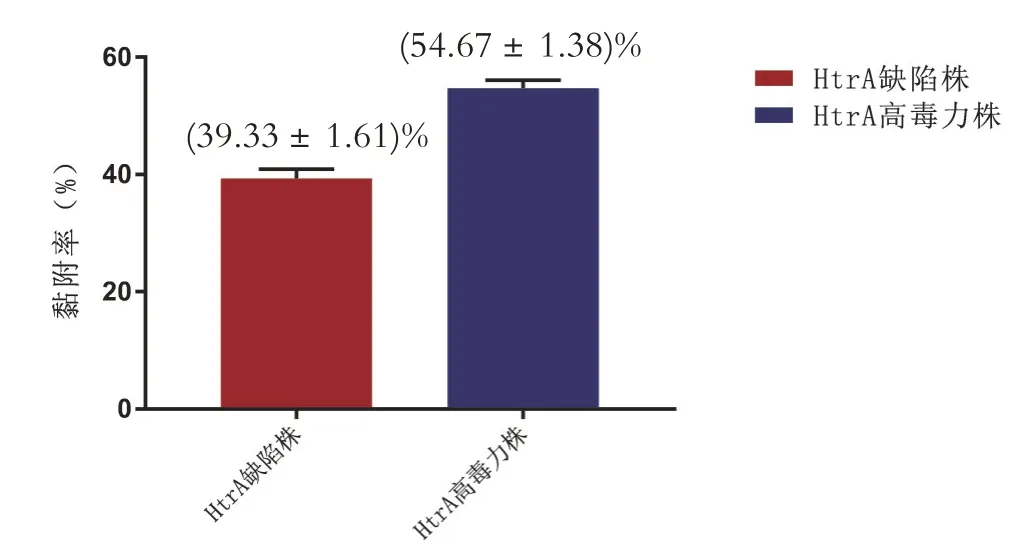
图4 两菌株在唾液包被的羟基磷灰石表面的黏附率的比较(P<0.05)
2.4 荧光显微镜下两菌株黏附观察
如图5、6 所示,A、D 分别为荧光显微镜下缺陷株和高毒力株在羟基磷灰石表面的黏附情况,B、E分别为普通光镜下同一视野观察,C、F 为荧光显微镜和普通光镜观察图片合并的情况,可见HtrA缺陷株在羟基磷灰石粉末表面的黏附比例少于HtrA 高毒力株。
3 讨论
在龋病病因学的研究中,大量证据表明细菌是龋病发生发展先决条件,而变异链球菌是口腔龋病中最重要的致龋菌,一直以来都是各国学者龋病防治研究的一个热点。有研究[9-10]表明包括变异链球菌在内的多数链球菌中,HtrA基因位于染色体复制起始点附近,包含有与染色体复制和细菌增殖有关的DnaA 蛋白的结合位点。本实验结果表明在非应激条件下,HtrA基因缺失对乳牙变异链球菌高毒力株增殖速度产生了一定影响。因此推测HtrA 基因缺失影响了乳牙高毒力变异链球菌染色体的复制过程从而影响了它的生长增殖速度。变异链球菌在长期进化过程中形成了多种途径代谢碳水化合物产生乳酸等有机酸[11],有机酸持续作用于牙釉质表面,导致牙釉质软化脱矿。本次实验结果显示:在酸性环境下,HtrA基因缺失降低了乳牙高毒力株变异链球菌产酸能力。Biswas 等[9]的研究显示HtrA 基因缺陷可能影响烯醇酶和甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase)的表达,这两种酶都是与糖酵解有关的细胞外酶。由此可以推测,HtrA是通过影响糖酵解过程中相关的酶,从而影响了细菌产酸的能力。
变异链球菌对牙面的黏附是形成牙菌斑的前提和致龋的重要条件,主要包括蔗糖依耐性黏附和非蔗糖依耐性黏附两种途径[12]。致龋菌对牙面的蔗糖非依赖性黏附,实际上是细菌表面的黏结素与获得性膜中受体成分的特异性结合[13]。致龋菌对牙面的蔗糖依耐性黏附与葡聚糖介导的细菌黏附和聚集相关,并且葡糖基转移酶(glucosyltransferase,GTF)和葡聚糖结合蛋白(glucan binding protein,GBPs)是与该过程相关的主要蛋白。Ahn 等[14]的研究表明在变异链球菌中只存在一个HtrA基因,且其对变异链球菌生物膜形成起重要调控作用。GTF利用蔗糖合成具有黏性的非水溶性葡聚糖来介导变异链球菌蔗糖依耐性黏附,GTF 还可进入早期获得性膜并合成葡聚糖,给GBPs 提供结合位点,直接影响变异链球菌的初始黏附过程[15]。李政等[16]研究表明变异链球菌HtrA缺陷株GTF的表达高于HtrA高毒力菌株,但是其生物学活性弱于HtrA高毒力菌株,推测HtrA基因可能不是简单影响GTF 的产量,而是影响了GTF 向细菌表面的转运,同时,HtrA 基因可能对GTF 等蛋白的折叠发挥调节作用。本研究中HtrA 缺陷株在唾液包被的羟基磷灰石表面的黏附率低于HtrA 高毒力株,原因可能是HtrA蛋白作为一种蛋白酶和分子伴侣,对蔗糖依耐性黏附途径中GTFs 和GBPs 等重要蛋白的折叠和胞内向胞外转运过程中起重要调节作用,HtrA 基因缺陷导致HtrA 蛋白表达异常,从而影响了变异链球菌的黏附能力。
4 结论
本研究表明HtrA 基因缺陷会影响乳牙变异链球菌的生长、产酸及黏附能力,但其具体影响机制还需进一步研究。
