不同供氮形态下水稻苗期磷吸收累积与根系形态的关系
2019-03-22陈晨龚海青金梦灿郜红建
陈晨 龚海青 金梦灿 郜红建
不同供氮形态下水稻苗期磷吸收累积与根系形态的关系
陈晨 龚海青 金梦灿 郜红建*
(安徽农业大学 资源与环境学院/农田生态保育与污染防控安徽省重点实验室, 合肥 230036;*通讯联系人, E-mail: hjgao@ahau.edu.cn)
【目的】植物根系形态对于适应低磷胁迫具有一定的可塑性,对提高磷的吸收利用具有重要意义。因此,本研究以长江中下游地区主推的102个水稻品种为供试材料,研究根系形态与水稻幼苗磷吸收利用的相关性。【方法】采用国际水稻所营养液培养方法,研究在NH4+-N和NO3--N供应条件下苗期植株生物量、磷含量和磷素累积量及其与根系形态指标的相关性。【结果】研究结果表明,在相同供氮水平(40 mg/L)下,供应NH4+-N时,水稻苗期平均生物量为67.87 mg/株,比供应NO3--N时高4.27 mg/株;水稻苗期平均磷含量为0.49%,比供应NO3--N时高0.10%;水稻苗期平均磷累积量为0.37 mg/株,比供应NO3--N时高0.10 mg/株。在NH4+-N条件下,水稻根系形态指标变异系数呈现根尖数>总根长>分支数>总根面积>交叉数>总根体积>平均根系直径的规律;在NO3--N条件下,水稻根系形态指标变异系数呈现根尖数>分枝数>总根长>总根面积>交叉数>总根体积>平均根系直径的趋势。在NH4+-N条件下,总根长、总根面积、分枝数、交叉数四个形态指标与植株生物量、磷含量、磷累积相关最为显著(<0.01),而在NO3--N培养下,总根长、总根面积、根尖数、交叉数与植株生物量及磷素吸收累积指标相关性最为显著(<0.01)。【结论】供应氨态氮,水稻营养指标与根系形态指标的相关性更高。水稻苗期根系总根长、总根面积、交叉数可作为水稻磷高效评价的重要指标。
水稻;氮素形态;根系形态;磷吸收累积
根系是作物从外界吸收营养物质及水分的主要器官,对于植物的生长发育有着至关重要的作用[1]。磷是影响水稻产量形成的重要营养元素,土壤中的磷易被土壤颗粒固持或固定,有效性低,已成为限制作物产量的重要因素之一。在植物生长过程中,根系会启动有利于植物高效吸收、利用土壤中磷素的适应性机制,包括根的形态特征演变(如增加根毛长度和密度)[2-4]、根构型变化[5,6]、特异根系分泌物的分泌[7]等。水稻利用氮的形态主要有NH4+-N和NO3--N,两种氮素在吸收、运输、储藏和同化等方面存在较大差异,很大程度上影响水稻的生长和对磷素的吸收与利用[8]。究其原由,不同形态氮肥引起根际土壤pH变化,导致磷素的吸收差异。作物供给NH4+-N时,等量分泌质子(H+)引起根际pH降低,促进了根际土壤中磷的溶解和植物对磷的吸收;供给NO3−-N时,根系非等量分泌HCO3−或OH−引起根际pH升高,抑制难溶性磷向土壤溶液中转移及植物对磷的吸收[9]。利用作物本身的生物学特性,通过调节根系的生长发育和形态结构,对于挖掘作物自身对磷素高效吸收利用的潜力,对磷素的吸收利用具有重要意义。
根系形态及其可塑性对植物磷效率具有重要作用[11-13]。根长与吸磷量正相关,而根半径与磷的吸收效率成反比[14]。植株具有合理的三维空间分布和较长的总根长,其磷效率和产量高[15]。在水稻生产中,根系形态参数是决定其是否能高效吸收和利用磷的关键性因素[16]。已有的研究多关注根生长分布特征的定性描述,对根系表面积、根系直径、根尖数等形态参数缺少有效的分析手段,有关水稻根系形态与磷吸收利用之间的关系尚不清楚。研究水稻的根系形态在不同供氮形态下的适应性差异及其在水稻磷吸收、累积的影响,明确根系形态与磷吸收累积的相关性,对于遴选磷高效水稻品种,提高磷利用效率、减少磷肥施用降低环境污染具有重要意义。
本研究采用营养液培养方法,利用根系专用扫描仪(Epson perfection V700 photo,日本)和根系形态专用分析软件(Win RHIZO_Pro V2007d, Regent Instrument Inc. 加拿大)研究了102个水稻品种在相同浓度的NH4+-N和NO3--N供应条件下苗期植株生物量、磷含量和磷累积量及其与总根长、总根面积、总根体积、平均根系直径、根尖数、分支数、交叉数等根系形态指标的相关性。通过对根系形态指标进行变异特征分析,拟合植株生物量、磷含量、磷累积量与根系形态指标之间的最优回归模型,为磷高效种质的筛选提供理论依据。
1 材料与方法
1.1 试验材料与设计
试验材料包括长江中下游地区主推的102个水稻品种/系(编号1~102),分别由湖南省水稻研究所、安徽农业大学、安徽省农业科学院和南京农业大学提供(表1)。
选择饱满一致水稻种子,用10%的双氧水消毒30 min后,自来水冲洗2~3遍,再用蒸馏水清洗干净,放入底部垫有浸润滤纸的发芽盒。水稻种子置于28 ℃恒温培养箱中避光催芽,待出现胚芽鞘后,移至安科发芽纸上培养。将发芽种子沿宽边均匀放置在发芽纸(规格38#: 25.4 cm×38.1 cm)上,卷成圆筒状后用亚克力支撑架固定,垂直放入蒸馏水中,光照培养2 d后,依次在国际水稻研究所1/4和1/2标准浓度的水稻专用营养液中分别培养4 d,1倍标准浓度的水稻专用营养液中培养7 d后,用蒸馏水冲洗水稻根系后,移入相同氮浓度(40 mg/L)的NH4+-N和NO3--N营养液中培养10 d。国际水稻研究所水稻专用营养液的组成如下: NH4NO3408 mg/L,NaH2PO4·2H2O 46.8 mg/L,K2SO4174 mg/L,MgSO4·7H2O 393.6 mg/L,CaCl2111 mg/L,MnCl2·4H2O 1.98 mg/L,(NH4)6Mo7O24·4H2O 0.124 mg/L,H3BO30.62 mg/L,ZnSO4·7H2O 0.0574 mg/L,CuSO4· 5H2O 0.1 mg/L,FeSO4·7H2O 19.5 mg/L,柠檬酸 30 mg/L。含NH4+-N和NO3−-N的营养液分别用(NH4)2SO4和KNO3替换NH4NO3,含NH4+-N营养液中加入5.89 mg/L的双氰胺(C2H4N4)以抑制可能发生的硝化作用,同时设置只加入5.89 mg/L的双氰胺(C2H4N4)不含NH4+-N的营养液为对照。培养液每2 d更换一次,pH用1 mol/L 盐酸或1 mol/L NaOH 调节至5.5±0.1。试验分为NH4+-N和NO3--N两个处理,每个处理3个重复,每个重复6株水稻幼苗,水稻培养至6叶期时收获[17]。

表1 供试水稻品种(系)及编号
1.2 测定项目及方法
水稻培养至6叶期时,分别收获植株的茎叶和根部样品。部分茎叶和根部样品于105℃下杀青30 min后,置75℃下烘干至恒重,称量茎叶干质量与根部干质量,计算整株生物量、磷含量、磷累积量。水稻茎叶和根部样品经H2SO4-H2O2消煮,钒钼黄比色法测定植株全磷含量。茎叶磷累积量、根部磷累积量和整株磷累积量分别为茎叶、根部、整株生物量与其磷含量之积。
收获时,每个水稻品种从3个重复试验中各选择1株水稻用于根系形态分析。新鲜水稻根系先用蒸馏水冲洗干净表面附着物后,用根系形态专用扫描仪(Epson perfection V700 photo, 日本)数字化扫描后,根系形态分析专用软件(WinRHIZO _Pro V2007d, Regent Instrument Inc. 加拿大)分析总根长、总根面积、总根体积、平均根系直径、根尖数、分支数、交叉数等根系形态参数,根系形态参数为3次重复试验的平均值。
1.3 数据分析方法
数据采用Excel 2007和SPSS 19.0软件进行统计分析,采用检验分析不同参数之间的显著(<0.05)和极显著水平(<0.01)。根系形态指标采用根系分析系统软件WinRHIZO_Pro V2007d (Regent Instrument Inc.,加拿大)进行定量分析。采用逐步回归分析建立NH4+-N和NO3−-N供应条件下水稻干质量、磷含量、磷累积量与根系形态参数之间的最优回归方程。
2 结果与分析
2.1 不同氮素培养下水稻苗期各性状分析
水稻在NH4+-N和NO3−-N培养条件下的苗期植株生物量差异显著。在NH4+-N条件下,水稻生物量为11.31~150.77 mg/株,平均为67.87 mg/株,变异系数为52.99%,其中以两优766的生物量最高,为150.77 mg/株,以丰两优品种的生物量最低,为11.31 mg/株。在NO3−-N条件下,水稻生物量为11.73 ~141.27 mg/株,均值为63.60 mg/株,变异系数为53.96%,其中以甬优9号的生物量最高,为141.27 mg/株,以隆香优130的生物量最低,为11.73 mg/株。在相同供氮水平下,不同水稻品种在NH4+-N条件下的平均生物量高于其在NO3−-N条件下平均生物量(图1-A)。
水稻植株在NH4+-N条件下的磷含量为0.27%~0.76%,均值为0.49%,变异系数为25.48%。其中,以盐粳11的含磷量最高,为0.76%,以深两优3117的含磷量最低,为0.27%。水稻植株在NO3−-N条件下的磷含量为0.21%~ 0.70%,均值为0.39%,变异系数为28.00%。其中,以丰两优80的含磷量最高,为0.70%,以湘早籼24的含磷量最低,为0.21%。在相同供氮水平下,不同水稻品种在NH4+-N培养条件下的平均含磷量比其在NO3−-N条件下高0.10%(图1-B)。
水稻植株在NH4+-N培养下的磷累积量为0.03~1.12 mg/株,均值为0.37 mg/株,变异系数为66.82%。其中,以盐粳11号的磷累积量最高,为1.12 mg/株,以丰两优品种的磷累积量最低,为0.03 mg/株。在NO3−-N培养下,水稻磷累积量为0.03~0.91 mg/株,均值为0.27 mg/株,变异系数为70.63%。其中,以丰两优80的磷累积量最高为0.91 mg/株,以湘早籼24号品种的磷累积量最低,为0.03 mg/株。在相同供氮水平下,不同水稻品种在NH4+-N培养条件下的平均磷累积量高于其在NO3−-N条件下平均磷累积量(图1-C)。

1,2……102分别表示供试水稻品种编号;A图中单位为mg/株;B图中单位为%;C图中单位为mg/株
Fig. 1. Biomass(A), phosphorus content(B), phosphorus accumulation(C) of rice plants at the seedling stage cultured in NH4+-N and NO3−-N solutions.
不同大小写字母分别代表在0.01和0.05水平上差异显著。
Different uppercase and lowercase letters indicate significant differences at 0.01 and 0.05 levels, respectively.

图2 不同形态氮素营养对水稻根系形态的影响
Fig. 2. Effects of N forms on root morphology of rice plants.
2.2 不同氮素供应下水稻苗期根系形态性状变异特征
根系形态是品种间差异的重要特征,变异系数越大,水稻品种间的差异越大。NH4+-N和NO3−-N条件下,水稻根系形态指标均表现出一定的变异性,变异系数变幅相差较大。在NH4+-N条件下,水稻根系形态指标变异系数范围为19.28%~65.96%,后者为前者的3.47倍,变异系数呈根尖数>总根长>分枝数>总根面积>交叉数>总根体积>平均根系直径的趋势,其中平均根系直径变异系数最小,为19.28%;而根尖数变异系数最大,为65.96%。在NO3−-N条件下,水稻根系形态指标变异系数呈根尖数>分枝数>总根长>总根面积>交叉数>总根体积>平均根系直径的趋势,其变异范围为21.25%~63.17%;其中平均根系直径变异系数最小,为21.25%,根尖数的变异系数最大,为63.17%(表2)。
由根系扫描图(图2)可以看出,NH4+-N条件下水稻的总根长、总根面积、根尖数、交叉数明显较NO3−-N条件下增加,而不同氮形态供应下水稻的平均根系直径、总根体积、交叉数并无明显差异。NH4+-N营养对水稻根系发育的促进作用较NO3−-N明显。
2.3 不同氮素供应下水稻苗期磷营养指标与根系形态相关性分析
供应NH4+-N,水稻整株生物量与平均根系直径呈显著负相关(相关性系数为−0.22),而分别与总根长、总根面积、总根体积、根尖数、分枝数、交叉数呈极显著正相关,相关性系数为0.81~0.93(表3)。总根长、总根面积、根尖数、交叉数与植株生物量相关性最为显著。水稻整株磷含量与总根长、总根面积、总根体积、根尖数、分枝数、交叉数呈现显著极显著负相关,相关性系数为0.54~0.65;而与平均根系直径呈现负相关,但相关性系数分别仅为−0.15。水稻整株磷累积量除与平均根系直径呈负相关外(相关性系数为−0.21),与其他根系形态指标皆呈极显著正相关,相关性系数为0.77~0.88。总根长、总根面积、分枝数、交叉数与植株磷吸收累积相关性最为显著。
在NO3−-N条件下,水稻整株生物量与平均根系直径呈显著负相关,而分别与总根长、总根面积、总根体积、根尖数、分枝数、交叉数呈极显著正相关,相关系数为0.77~0.97。这与在NH4+-N供应下的相关性规律类似,而总根长、总根面积、根尖数、交叉数与植株生物量相关性最为显著。水稻整株磷含量与总根长、总根面积、总根体积、根尖数、分枝数、交叉数呈极显著正相关,相关系数为 0.33~0.56;而与平均根系直径呈负相关,但相关性系数分别仅为−0.17。水稻整株磷累积量除与平均根系直径呈负相关外(相关系数为−0.22),与其他根系形态指标皆呈现极显著正相关,相关性系数为0.63~0.89。总根长、总根长、总根面积、根尖数、交叉数与植株磷吸收累积相关性最为显著。

表3 不同供氮形态下水稻生物量、磷养分指标与根系形态指标相关性分析
**表示在<0.01水平上呈极显著相关,*表示在<0.05水平上呈显著相关。
**means highly significant correlation at the 0.01 level (bilateral). *means significant correlation at the 0.05 level (bilateral).
2.4 不同氮素供应下水稻营养指标与形态指标的多元线性回归分析
不同氮素供应条件下水稻植株生物量、磷含量和磷累积量与根系形态指标多元线性逐步回归方程中,X对Y的决定系数皆呈显著相关(表5),其在NH4+-N供应条件下的决定系数(2)范围为0.27~0.92,低于其在NO3−-N条件下的决定系数(2)范围(0.40~0.94)。
在NH4+-N供应条件下,水稻植株生物量(1)、磷含量(2)、磷累积量(3)与根系形态指标之间的最优回归模型分别如下:
1=−2.37+2×10-31+0.192+10-36+10-371)
2=0.01−4.09×10-772)
3=−22.75+8×10-36+ 4×10-373)
其中,在NH4+-N供应条件下,水稻不同品种根系形态指标中以总根长、总根面积、分枝数、交叉数对植株生物量的影响最大;以交叉数对植株磷含量的影响最大;以分枝数、交叉数对植株磷累积量的影响最大,且均达到了显著差异水平。
在NO3−-N供应条件下,水稻植株生物量(4)、磷含量(5)、磷累积量(6)与根系形态指标之间的最优回归模型分别如下:
4=−0.30+5.60×10−21+0.1724)
5=0.27−1.03×10−56+2.03×10−575)
6=−4.49+4×10−21+1×10−366)
在NO3−-N条件下,水稻不同品种根系形态指标中以总根长、总根面积对植株生物量的影响最大;以分枝数、交叉数对植株磷含量的影响最大;以总根长、分枝数对植株磷累积量的影响最大,且均达到了显著差异水平。
3 讨论
植物吸收养分的能力主要取决于根系,其中,根系形态性状起着决定性的作用[18]。本研究发现,在不同供氮形态下,水稻植株的全磷含量和单株磷累积量上具有较大差异,植株根系形态学的差异也较显著。这表明水稻不同供试品种植株磷累积量的差异主要与根系形态学上的差异有关。水稻磷高效吸收与其根系形态关系极为密切,良好的根系形态是水稻磷高效的重要基础[19,35]。本研究结果表明,水稻根系形态中的总根长、总根面积、交叉数三个形态指标与水稻生物量及磷吸收累积量相关性最为显著,即水稻可通过发育良好的根系形态来提高根系对磷的吸收和利用能力,从而提高地上部磷含量和磷积累量[20]。具有较好根形态构型的植株基因型,有利于从耕层土壤中吸收有效磷和其他养分,其产量和磷效率均较高[15]。水稻作为须根系植物,根系分布密度大,根长延展深,有利于形成不定根及次生侧根适当分散、均匀分布,多数根留在表层吸磷、少数根扎到深处吸水的植株磷高效吸收理想根系特征,这与廖红等的观点基本一致[21]。前人研究发现,植物对磷素的吸收能力与位于根系根毛和根表皮细胞质膜上磷转运蛋白(phosphate transporter)的亲和性及其转运生长介质中Pi的能力和效率密切相关[22]。因此,根系表皮和根毛细胞质膜上的高亲和磷转运蛋白的分布数量多、亲和介质中P的K值低和磷转运蛋白在膜内外的拓扑变构效率高,是磷高效水稻品种具有较多磷素累积量的分子生物学原理[37]。
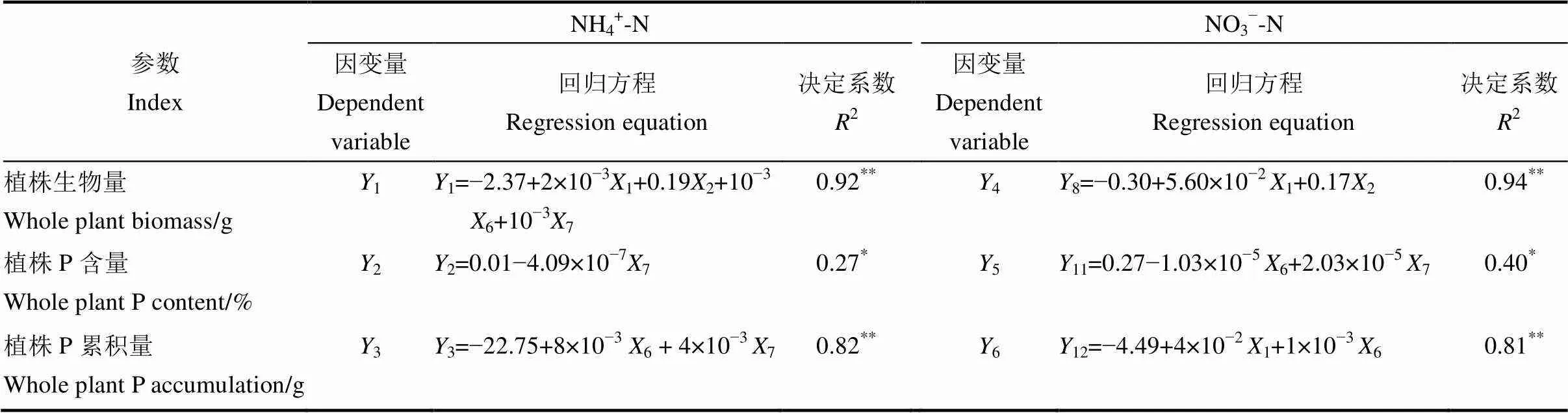
表4 不同氮形态供应下水稻营养指标与形态指标多元逐步回归分析
X(=1,2……7)分别表示水稻根系总根长、总根面积、平均根系直径、总根体积、根尖数、分枝数、交叉数。
X(=1,2……7) represent total root length, root area, average diameter, root tip number, root branching number and root crossing number.
根系吸磷效率不仅与根系形态及品种的遗传特性有关,同时也取决于氮素形态[23,24]。作物根系吸收NH4+-N和NO3−-N后,由于在吸收、储存、运输、同化过程上存在较大差异,导致根系形态发生变化,进而影响着作物对磷素的吸收利用[25-26]。已有的研究表明:NH4+-N进入植物细胞后与有机酸结合,形成氨基酸或酰胺,而NO3−-N在进入植物体后一部分还原成NH4+-N,并在细胞质中进行代谢,再参与水稻吸收代谢过程[27-29]。NH4+-N的吸收和还原比NO3−-N的吸收还原减少了能量的消耗,因此,NH4+-N条件下具有优越的根形态特征,根系长度、体积、分布密度和有效吸收面积均较大,这均有利于水稻根系与养分介质的接触,提高磷的吸收能力[30-31]。本研究结果也证实,在单供NH4+-N的体系中,水稻的根系形态体系要高于单供NO3--N的体系,这是由于植物根系形态学的不同导致了其吸磷特性的差异[32]。植物根系在营养介质中分布面积越广、延展越深越有利于其磷素的吸收利用[33]。在NH4+-N供应下,水稻根系具有发根力强、吸收面积大、分布密度大等特点,加快了地上部生长和主根伸长的速度,从而提高了磷的吸收利用效率。此外,NH4+-N和NO3−-N所带电荷的不同,也导致了植物对磷素吸收、利用上的差异。NH4+-N供应有利于阴离子的吸收,而NO3−-N供应可以促进阳离子的吸收。这是因为供应NH4+-N时,植株会吸收大量NH4+-N进入体内,为了维持电荷平衡,植株会减少阳离子的吸收;同时,NH4+-N在根表的集聚对其他阳离子具有排斥作用,在一定程度上降低了其他阳离子的吸收,促进了阴离子吸收[34]。NO3−-N的作用正好相反,会减少阴离子的吸收,增加阳离子的吸收[35]。因此,硝态氮供应条件下水稻磷素累积降低的原因可能与硝态氮和磷酸根同为阴离子,水稻根系吸收存在竞争关系,特别是竞争根系细胞质膜上的转运蛋白/载体有关[28,36]。
[1] Liedgens M, Soldati A, Stamp P. Root development of maize (L.) as observed with minirhizotrons in lysimeters., 2000, 40(6): 1665-1672.
[2] Yan X, Liao H, Beebe S E, Blair M W, Lynch J P. QTL mapping of root hair and acid exudation traits and their relationship to phosphorus uptake in common bean., 2004, 265(1/2): 17-29.
[3] Bates T R, Lynch J P. Stimulation of root hair elongation in Arabidopsis thaliana by low phosphorus availability., 1996, 19(5): 529-538.
[4] Jungk A. Root hairs and the acquisition of plant nutrients from soil., 2015, 164(2):121-129
[5] Bonser A M, Lynch J, Snapp S. Effect of phosphorus deficiency on growth angle of basal roots in phaseolus Vulgaris., 1996, 132(2): 281-288.
[6] Liao H, Rubio G, Yan X, Cao A Q, Brown K M, Lynch J P. Effect of phosphorus availability on basal root shallowness in common bean., 2001, 232(1/2): 69-79
[7] 沈宏, 严小龙. 根系分泌作用及其诱导机制. 生态环境学报, 2001, 10(4): 339-342.
Sheng H, Yan X L. Root exudation and their induced mechanism., 2001, 10(4): 339-342(in Chinese with English abstract)
[8] 张平. 氮形态对低磷胁迫下苗期玉米磷素吸收的影响及其机理. 南京: 南京农业大学, 2012.
Zhang P. Mechanisms and effects of nitrogen forms on phosphorus absorption of maize (L.) under phosphorus stress at seedling stage.Nanjing: Nanjing Agricultural University, 2012. (in Chinese with English abstract)
[9] 邹春琴, 杨志福. 氮素形态对春小麦根际pH与磷素营养状况的影响. 土壤通报, 1994(4): 175-177.
Zhou C Q, Yang Z F. Effects of different form nitrogen on pH and phosphorus content of spring wheat rhizosphere., 1994(4): 175-177. (in Chinese with English abstract)
[10] 李朝霞. 磷匮乏影响玉米根系发育机制的研究. 济南: 山东大学, 2011.
Li C X. Study on the mechanism of root developmental changes to phosphate deficiency in maize. Jinan: Shandong University, 2011. (in Chinese with English abstract)
[11] Chen X P, Cui Z L, Vitousek P M, Cassman K G, Matson P A, Bai J S. Integrated soil-crop system management for food security., 2011, 108(16): 6399-6404.
[12] 严小龙, 廖红, 戈振扬. 植物根构型特性与磷吸收效率. 植物学报, 2000, 17(6): 511-519.
Yan X L, Liao H, Ge Z Y. root architectural characteristics and phosphorus acquisition efficiency in plants., 2000, 17(6): 511-519. (in Chinese)
[13] 赵静, 付家兵, 廖红,罗锡文. 大豆磷效率应用核心种质的根构型性状评价. 科学通报, 2004, 49(13): 1249-1257.
Zhao J, Fu J B, Liao H, Luo X W. Properties evaluation of phosphorus efficiency on roots structure of soybean., 2004, 49(13): 1249-1257. (in Chinese)
[14] Forde B, Lorenzo H. The nutritional control of root development. Interactions in the root environment: An Integrated Approach. 2002: 51-68.
[15] 刘灵, 廖红, 王秀荣, 严小龙. 不同根构型大豆对低磷的适应性变化及其与磷效率的关系. 中国农业科学, 2008, 41(4): 1089-1099.
Liu L, Liao H, Wang X R, Yan X L. Adaptive changes of soybean genotypes with different root architectures to low phosphorus availability as related to phosphorus efficiency., 2008, 41(4): 1089-1099. (in Chinese with English abstract)
[16] 李佳. 水稻根系应答根际水分与磷素环境变化的研究. 扬州: 扬州大学, 2015.
Li J. Study on rhizosphere moisture and phosphorus environment change of rice root response. Yangzhou: Yangzhou University, 2015. (in Chinese with English abstract)
[17] 陈晨, 龚海青, 张敬智, 郜红建. 水稻根系形态与氮素吸收累积的相关性分析. 植物营养与肥料学报, 2017, 23(2): 333-341.
Chen C, Gong H Q, Zhang J Z, Gao H J. Correlation between root morphology and nitrogen uptake of rice., 2017, 23(2): 333-341. (in Chinese with English abstract)
[18] Lynch J. Root Architecture and Plant Productivity.1995, 109(1): 7-13.
[19] 李宝珍, 王松伟, 冯慧敏, 徐国华. 氮素供应形态对水稻根系形态和磷吸收的影响. 中国水稻科学, 2008, 22(6): 665-668.
Li B Z, Wang S W, Feng H M, Xu G H. Effects of nitrogen forms on root morphology and phosphate uptake in rice., 2008, 22(6): 665-668 (in Chinese with English abstract)
[20] Ladha J K, Kirk G J, Bennett J, Peng S, Reddy C K, Reddy P M, Singh U. Opportunities for increased nitrogen-use efficiency from improved lowland rice germplasm., 1998, 56(1/2): 41-71.
[21] 廖红, 戈振扬, 戈振场, 严小龙. 水磷耦合胁迫下植物磷吸收的理想根构型:模拟与应用. 科学通报, 2001, 46(8): 641-646.
Liao H, Ge Z Y, Yan X L, Yan X L. Optimized root architecture for phosphorus acquisition under water and phosphorus stress: Simulation and application., 2001, 46(8): 641-646. (in Chinese with English abstract)
[22] Muchhal U S, Pardo J M, Raghothama K G. Phosphate transporters from the higher plantthaliana., 1996, 93(19): 10519.
[23] 李宝珍, 辛伟杰, 徐国华. 氮饥饿水稻利用不同形态氮素的差异及其生理机制. 土壤学报, 2007, 44(2): 273-279.
Li B Z, Xin W J, Xu G H. Physiological mechanisms in uptake and use of different forms of nitrogen by nitrogen starved rice crop., 2007, 44(2):273-279. (in Chinese with English abstract)
[24] 范志强. 氮磷营养及氮形态对水曲柳幼苗生长和生理的影响机制. 沈阳: 东北林业大学, 2004.
Fan Z Q. Effects of nitrogen, phosphorus and nitrogen forms on growth and physiological traits of fraxinus mandshurica seedlings. Shenyang: Northeast Forestry University, 2004 (in Chinese with English abstract)
[25] 张彦东, 范志强, 王庆成, 王政权. 不同形态N素对水曲柳幼苗生长的影响. 应用生态学报, 2000, 11(5): 665-667.
Zhou Y D, Fan Z Q, Wang Q C, Wang Z Q. Effects of different nitrogen forms on growth ofseedlings., 2000, 11(5): 665-667.(in Chinese with English abstract)
[26] Miller M H. Effects of nitrogen on phosphorus absorption by plants., 1974.
[27] Thien S J, Mcfee W W. Effect of Nitrogen on phosphorus transport systems inL.1972, 36(4): 617-620.
[28] Smith F W, Jackson W A. Nitrogen enhancement of phosphate transport in roots ofL.: I. Effects of ammonium and nitrate pretreatment., 1987, 84(4): 1314-1318.
[29] Hirel B, Limami A M. Prospects for improving nitrogen use efficiency: Insights given by 15 N-labelling experiments., 2002, 2(1/2): 133-144.
[30] Radin J W. Differential regulation of nitrate reductase induction in roots and shoots of cotton plants., 1975, 55(2): 178-182.
[31] Zhang H, Xue YG, Wang ZQ, Yang J, Zhang J. Morphological and physiological traits of roots and their relationships with shoot growth in “super” rice., 2009, 113(1): 31-40.
[32] Mackown C T, Jackson W A, Volk R J. Restricted nitrate influx and reduction in corn seedlings exposed to nitrate and ammonium nutrition., 1982, 62(9): 353-359.
[33] Bonser A M, Lynch J, Snapp S. Effect of phosphorus deficiency on growth angle of basal roots in Phaseolus vulgaris., 1996, 132(2): 281-288.
[34] Ingestad T, Agren G I. The Influence of plant nutrition on biomass allocation., 1991, 1(2): 168.
[35] Serna M D, Borras R, Legaz F, Primo-Millo E. The influence of nitrogen concentration and ammonium /nitrate ratio on N-uptake, mineral composition and yield of citrus., 1992, 147(1): 13-23.
[36] Muchhal U S, Pardo J M, Raghothama K G. Phosphate transporters from the higher plant., 1996, 93(19): 10519.
Correlation Between Root Morphology and Accumulation of Phosphorus in Rice Seedlings under Different N Forms
CHEN Chen, GONG Haiqing, JIN Mengcan, GAO Hongjian*
(,,,,;,:)
【Objective】Plant root morphology has certain plasticity for adapting to low phosphorus stress, and is of great significance for improving phosphorus absorption and utilization. Therefore, we used 102 major rice varieties in the middle and lower reaches of the Yangtze River as the tested materials to study the correlation between root morphology and phosphorus uptake of rice seedlings. 【Method】Hydroponic experiments were conducted to investigate the influences of NH4+-N and NO3−-N at the same concentration (40 mg∙L-1) on dry weight, P concentrations and contents as well as the root morphology.【Result】At the six-leaf stage of rice incubated under the same concentration of nitrogen, the average biomass of the rice seedlings amounted to 67.87 mg per plant in the NH4+-N solution, 4.27 mg higher than that in the NO3--N solution. The average P content was 0.49% when grown in the NH4+-N solution, 0.10% higher than that of rice seedlings grown in the NO3--N solution. In addition, the average accumulation of P in the seedlings was 0.37 mg per plant when supplied with NH4+-N, 0.10 mg higher than that supplied with NO3--N. The coefficient of variation of root morphological indexes were in the order of the number of tips > total root length > the number of branches > total root area > the number of crossing > the total root volume > average diameter in the NH4+-N solution; the coefficient of variation of root morphological indexes were in the order of the number of tips > the number of branches > total root length > total root area > the number of crossing > the total root volume > average diameter in the NO3--N solution. The total root length, total root area, branching number and crossing number were significantly correlated with biomass, P content, and P accumulation in rice plants in the NH4+-N solution; whereas the total root length, total root area, tip number and crossing number were significantly correlated with biomass, P content, and P accumulation in rice plants in the NO3--N solution.【Conclusion】The correlations between P uptake and the indices of root morphology when plants were supplied with NH4+-N were higher than those supplied only with NO3--N. The total root length, total root area and crossing number might be important indicators for evaluating the P use efficiency of rice at seedling stage.
rice; nitrogen forms; root morphology; P uptake and accumulation
10.16819/j.1001-7216.2019.8059
S143.1; S143.2; S511.01
A
1001-7216(2019)02-0167-09
2018-05-07;
2018-11-26。
国家科技支撑计划资助项目(2013BAD07B08);国家重点研发计划资助项目(2016YFD0200107, 2016YFD0300801, 2016YFD0300900)。
