滩羊EphA3基因克隆表达分析及生物信息学初步研究
2019-03-19徐芹芹刘玉芳康晓龙方美英
徐芹芹,刘玉芳,康晓龙,方美英
(1.中国农业大学动物科学技术学院,北京 100193;2.中国专利技术开发公司,北京 100193;3.河北工程大学生命科学与食品工程学院,河北邯郸 056021;4.宁夏大学农学院,宁夏银川 750021)
酪氨酸激酶受体3(EphA3)是Eph(Erythropoietin Producing Human Hepatocellular)受体家族中的一员,依据胞外结构域的序列同源性和优先与A类或B类Ephrin结合的能力可细分为EphA和EphB 2类[1]。EphA3作为EphA类受体的一员,属于单次跨膜分子,分子的胞外区含有配体结构域,胞内区含有多个磷酸化位点、激酶结构域和SH3结构域(Src同源结构域)等,这些结构决定了EphA3基因的功能[2]。EphA3基因的表达具有明显的时空性,在胚胎组织中高表达[3-4]。此外,EphA3基因参与机体的多种调控,在胚胎发育过程中发挥着重要的调控作用[5]。
EphA3基因在毛发生长过程中发挥着一定的作用。研究表明,EphA3可以加快生长初期小鼠毛发的发育、增加毛囊密度、加快毛囊的成熟过程等[6]。EphA3的表达贯穿于整个毛发生长周期:在毛发生长初期,EphA3在毛囊表皮和上皮细胞中表达;在过渡期,EphA3蛋白在新发育的次级毛芽中被发现,如毛囊膨大部区域;在第2个生长周期中,EphA3也在向下生长的毛囊中被检测到。上述研究表明,EphA3在动物毛发生长中具有重要的调控作用[7-8]。结合本课题组前期实验结果[9],推测EphA3基因可能参与滩羊被毛表型形成的生物学过程。本研究旨在通过对滩羊EphA3基因编码区序列进行结构特征分析,并结合其在不同时期的表达模式,为系统解析EphA3基因在滩羊被毛卷曲形成过程中的作用提供分子依据。
1 材料与方法
1.1 材料 选用1月龄及48月龄宁夏滩羊各15只,屠宰后分别取其心脏、肝脏、脾脏、胃、肺、肾脏、眼肌、皮肤组织,放入冻存管中并于液氮中迅速冷冻,然后迅速放入-80 冰箱中保存备用。
1.2 滩羊各组织RNA提取 取约40 mg 1月龄和48月龄滩羊的心脏、肝脏、脾脏、胃、肺、肾、眼肌及皮肤组织,采用TriZol与试剂盒(天根生化科技有限公司,北京)结合的方法分别进行总RNA提取,取出2 μL提取的总RNA在1.5%琼脂糖凝胶上电泳检测完整性,利用NanoDrop 1000进行RNA的浓度检测,其A260/A280值在 1.7~2.0。
1.3 反转录合成cDNA 利用反转录酶的转录活性,将mRNA转录成cDNA第一链。反应体系按照反转录试剂盒步骤进行反转录(天根生化科技有限公司,北京)。反转录产物置于-20 冰箱保存。用三磷酸甘油醛脱氢酶(GAPDH)基因进行普通PCR扩增,检测反转录合成cDNA的质量,扩增产物在1.5%琼脂糖凝胶上电泳检测。
1.4 CDS区克隆测序 利用Primer 5设计EphA3基因的CDS区引物,以NCBI中GenBank提供的预测绵羊EphA3基因mRNA序列为模板,登录号为XM_004002 846。克隆EphA3基因CDS区所用的引物由生工生物工程(上海)股份有限公司合成(表1)。PCR扩增反应体系:2×Taq PCR mix 10 μL,ddH2O 8 μL,上、下游引物(10 pmol/L)各 0.5 μL,模板 1 μL。58 反应条件:95 5 min 后进入循环;95 30 s,30 s,72 30 s,34个循环;72 10 min;4 保存。之后取PCR产物在1.5%琼脂糖凝胶中电泳,检验PCR产物的质量。
将PCR产物切胶回收后,连接至PMD-18T载体中进行单菌株克隆,最后送至测序公司(深圳华大基因科技有限公司,深圳)进行单菌株测序。具体实验操作步骤按照切胶回收试剂盒(普洛麦格生物技术有限公司,北京)和PMD-18T说明书进行操作。
表1 扩增目的基因CDS区的引物
1.5EphA3基因生物信息学分析 将克隆的滩羊EphA3基因CDS区的序列与 NCBI上提供的小鼠、大鼠、原鸡、牛、黑猩猩、野猪、家猫、非洲象、大熊猫、马和人的EphA3基因序列进行生物信息学分析。使用DNAMAN和ChromosPro软件进行序列拼接比对,采用Cluxtal X和MEGA5进行同源性比对及构建系统进化树;使用在线软件Epasy(http://web.expasy.org)服务器上的Protparam、ProtScale等程序对蛋白质的物理特性进行预测;采用NCBI的Conserved Domains在线工具进行蛋白质保守性预测;采用TMHMM Server v.2.0在线软件分析该蛋白的跨膜结构;采用在线软件SOPMA(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)进行蛋白质二级结构预测。
1.6 组织表达谱分析 采用半定量PCR方法研究滩羊EphA3基因在心脏、肝脏、脾脏、胃脏、肺脏、肾脏、眼肌及皮肤组织中的表达情况,以GAPDH作为内参基因。用于半定量的引物(生工生物工程股份有限公司,上海)见表2。

表2 扩增目的基因及持家基因的引物
为了确定半定量的最佳循环数,采用不同循环数进行实验,最终确定能够表明EphA3基因在不同组织中的表达情况。PCR体系参照1.4中反应体系,共34个循环;产物4 保存。之后取PCR产物在1.5%琼脂糖凝胶中电泳,用于半定量结果的分析。
1.7 荧光定量PCR及数据分析 使用Primer 5引物设计软件进行引物设计,参考序列来源同表1。其中EphA3-QF/R为该基因跨越内含子设计的定量引物(表2),以GAPDH基因作为内参基因。
将滩羊1月龄及48月龄滩羊各15个个体反转录得到的cDNA各取2 μL,混池。以混池cDNA为模板,以2倍稀释为梯度进行稀释,稀释5个梯度,制作q-PCR的标准曲线。按照荧光定量PCR反应体系在CFX96TM Real-Time System定量PCR仪上进行实验,反应条件:95 30 s;95 10 s,58 30 s,39 个循环;95 10 s,4 保存。根据PCR反应的动力曲线图、线性图谱和半对数图谱选择适当的ct 值和域值,以得到理想的标准曲线图。采用2-△△ct法计算相对表达量,并利用SAS 8.1(ANOVA)进行显著性差异分析。
2 结 果
2.1 组织总RNA提取结果 滩羊皮肤组织总RNA提取结果经过1.5%琼脂糖凝胶电泳检测显示,RNA主条带清晰,提取质量较好,无明显降解,可用于下游实验。
2.2 CDS区克隆测序 用EphA3-1、EphA3-2、EphA3-3 3对引物进行PCR扩增,将产物在1.5%的琼脂糖凝胶中进行电泳,分别可见目的片段大小的特异性条带。克隆测序后用DNAMAN对序列进行拼接,从而获得滩羊EphA3基因3 109 bp的核苷酸序列,序列分析发现包括2 955 bp的完整开放阅读框,共编码984个氨基酸。将该序列与绵羊的EphA3基因完整CDS序列进行同源性比较,发现其同源性为99.9%,推定该序列即为滩羊EphA3基因的完整CDS序列。
2.3EphA3基因生物信息学分析
2.3.1 氨基酸序列分析 利用ExPASy的Protparam软件对EphA3蛋白质序列理化参数进行分析,其蛋白分子式为C4877H7658N1328O1477S53,分子量为110 227.6 u,理论等电点为6.48,总平均亲水性(GRAVY)为-0.307,由于GRAVY值的范围为-2~2,正值表明此蛋白为疏水蛋白,负值表明为亲水蛋白,所以EphA3蛋白为亲水蛋白。
利用NCBI的Conserved Domains在线工具预测蛋白的保守区,结果表明该序列有6个保守区,分别位于 29~201、263~303、326~432、437~528、617~883、913~975位。其中包括1个配体结合域EphR_LBD_A3,1个酪氨酸蛋白激酶催化域PTKc_EphR_A,1个SAM结构域,2个Ⅲ型纤连蛋白(fibronectintypeⅢ,FN3)结构域。
利用ExPASy的ProtScale软件对蛋白质亲疏水性进行分析。氨基酸组成是蛋白质折叠的主要驱动力,蛋白质折叠时会形成疏水内核和亲水表面,同时在潜在跨膜区出现高疏水值区。结果表明,该蛋白质存在数个高分值峰(Scare>1.5),分别位于 6~15、188~195、545~561、723~727等氨基酸位点区域;而几个低分值峰(Scare<0)分别位于 80、134、237、312、478、576、662、778、868等氨基酸位点附近,这些区域属高亲水性(图1)。

图1 EphA3蛋白的亲疏水性分析
利用TMHMM Server v.2.0在线软件分析,发现EphA3蛋白含有1个跨膜结构域,其跨膜螺旋区预测位置在542~564(图2)。
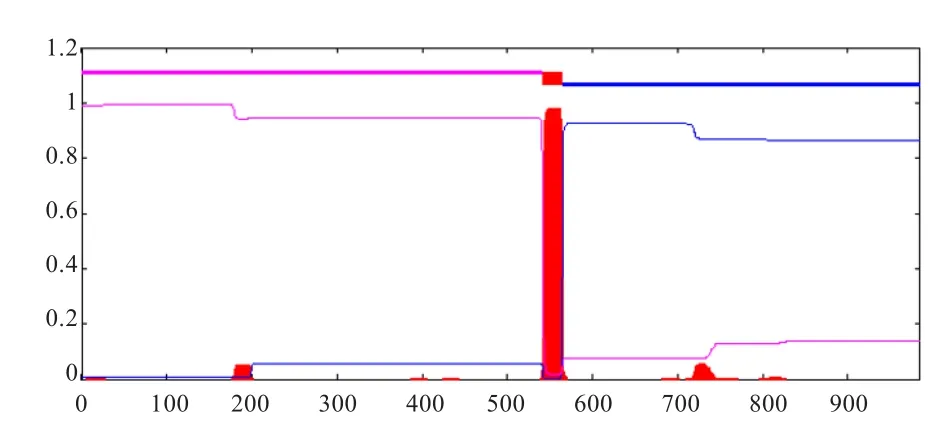
图2 EphA3蛋白的跨膜区预测结果
利用SOPMA软件进行蛋白质二级结构预测,结果表明,绵羊EphA3蛋白富含α-螺旋、延伸链及随机卷曲结构,且跨膜区所对应的序列位置正好存在α-螺旋,这也佐证了跨膜区域预测的准确性(图3)。
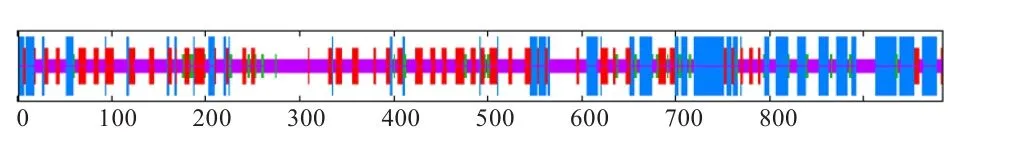
图3 EphA3蛋白质二级结构预测简图
2.3.2 同源性比较 利用Cluxtal X软件,将克隆获得的滩羊EphA3基因CDS区的序列与小鼠、大鼠、原鸡、牛、黑猩猩、野猪、家猫、非洲象、大熊猫、马和人的EphA3基因相应序列进行比对。由图4可见,滩羊EphA3基因核酸序列与牛的同源性最高,为98.5%,与原鸡的同源性最低,为81.2%。

图4 不同物种EphA3基因核苷酸序列差别(下三角)和相似性(上三角)
2.3.3 系统进化树分析 用MEGA5软件将各物种的EphA3基因的核苷酸序列进行比对构建N-J进化树(图5),其聚类结果与经典分类学结果基本一致,表现为滩羊与牛之间遗传距离最近,与野猪、马等哺乳动物遗传距离次之,与原鸡之间的遗传距离最远。这与羊、牛均属哺乳动物、偶蹄目、牛科有关,猪虽然也属偶蹄目,但科目不同,因此呈现出远近不同的亲缘关系。由聚类结果可知,EphA3基因在序列上的同源性很大程度上反映了各物种间的亲缘关系远近。滩羊EphA3基因CDS区编码984个氨基酸,通过蛋白质的BlastP比对发现,滩羊的EphA3蛋白与牛、野猪、大熊猫、人、非洲象的EphA3蛋白同源性较高,分别为99.4%、98.8%、98.5%、97.5%、97.4%。

图5 各物种EphA3蛋白的N-J进化树
2.4 组织表达谱 半定量PCR结果表明,在30~34个循环数时,基因表达量呈线性扩增,而且没有进入平台期,为达到理想的实验效果,本实验选择的循环数为34 。实验结果表明,EphA3基因在滩羊皮肤组织中表达量最高,在肺脏组织中表达量较高,在心脏、胃、肾脏、眼肌组织中有微量表达(图6)。

图6 滩羊EphA3基因(上)及GADPH基因(下)在各组织中的表达
2.5 荧光定量PCR结果 通过对30个滩羊皮肤组织cDNA混池进行2倍的梯度稀释,即2、2-1、2-2、2-3、2-45个梯度分析目的基因EphA3的标准曲线。由图7可见,所有的标准曲线回归分析均有1个高相关系数(R2=0.991>0.99),表明不同浓度的混池cDNA的模板扩增与ct值存在良好的线性关系。目的基因混池cDNA的扩增效率为100.6%,扩增曲线显示产物扩增重复性较好。目的基因熔解曲线为单峰,无杂峰,表明荧光定量PCR反应过程中无引物二聚体和非特异性扩增产物产生。故该反应条件和标准曲线可以用于EphA3基因的定量分析,所使用的目的基因的引物具有良好的特异性。
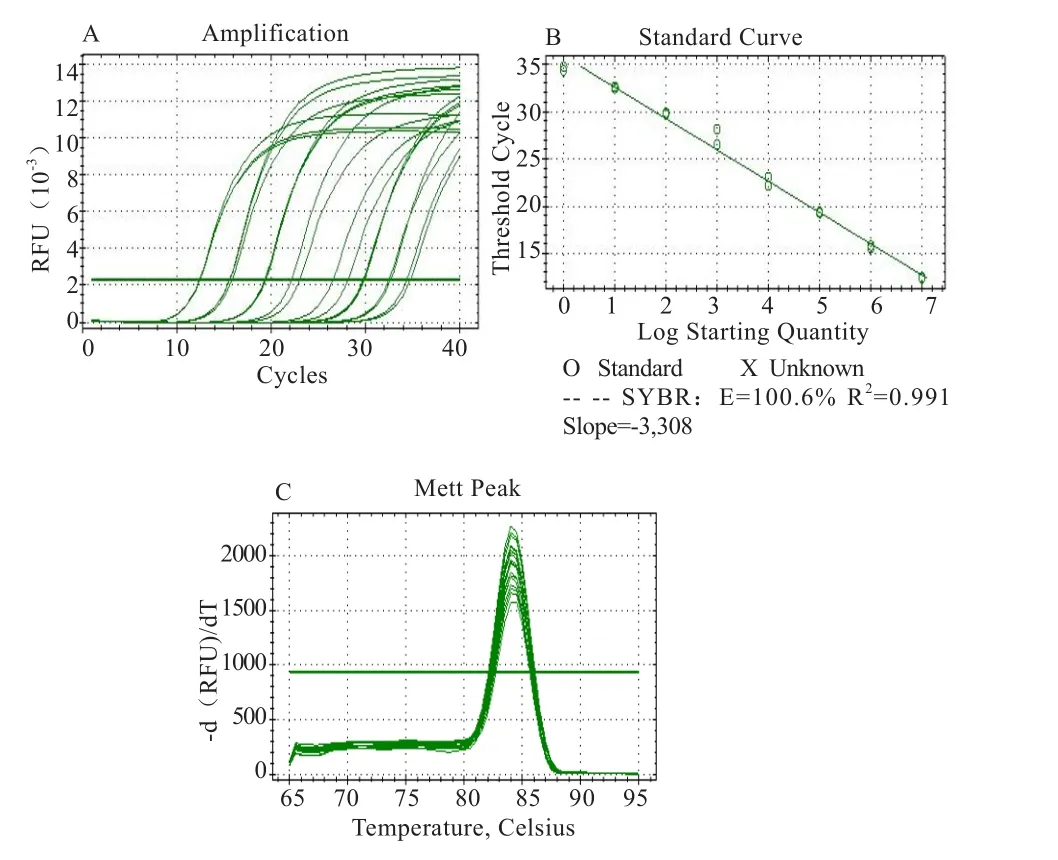
图7 10倍梯度稀释重组质粒DNA扩增形成的EphA3基因扩增曲线(A)、标准曲线(B)以及熔解曲线(C)
以GAPDH为内参基因,经过q-PCR检测并统计后发现,EphA3基因在48月龄滩羊皮肤组织中的相对表达量远高于1月龄(P<0.05)。
3 讨 论
滩羊是我国宁夏地区一个重要的地方品种,在滩羊被毛生长过程中被毛的形状发生着明显变化,1月龄滩羊被毛洁白并形成美丽的花穗,称为“二毛皮”, 特点为毛中短、花穗美观,具有波浪形花弯俗称“九道弯”,被当地人民誉为“宁夏五宝”之一的白宝,但是随着年龄的增长这种特性随之消失,在48月龄时已经观察不到这种美丽的花纹[10-11]。目前,EphA3对绵羊等经济动物毛发生长的调控作用几乎没有研究。因此,本研究以不同生长时期滩羊为研究对象,通过对滩羊被皮中EphA3基因编码区的克隆、结构特征生物学分析及表达模式分析,以期为揭示EphA3参与绵羊毛发生长的调控机理及生物学功能提供依据。
有文献报道,EphA3基因能够参与调节与毛发生长发育相关的过程。在人类、鼠等多个物种中都克隆得到了EphA3基因,并对该基因在毛囊生长中的调控作用进行了一系列研究。Yamada等[6]研究发现,注射了EphA3的老鼠毛囊中显示了弯曲的球状部位,这是毛发生长初期比较独有的特征,而对照组毛囊则保持垂直,这表明EphA3能够加速毛囊发育;同时,注射了EphA3的老鼠每单位长度皮肤中的毛囊数量明显增加。毛囊数量在胚胎期就已经决定而且不会在之后的生长过程中发生改变,一些没有发育成完整毛囊的毛芽会消失,然而如果EphA3过量,信号持续不断地作用于毛芽,它们就不会退化并可形成完整的毛囊。
本研究通过GenBank上提供的绵羊EphA3基因预测编码区序列设计了3对特异性引物,首次克隆获得了滩羊的EphA3基因编码区序列,全长2 955 bp,编码984个氨基酸(提交CDS区序列)。利用蛋白质的BlastP比对分析,发现滩羊的EphA3蛋白与牛的EphA3蛋白同源性最高(99.4%),与原鸡的同源性最低(81.2%)。N-J进化树分析发现,滩羊EphA3蛋白聚类结果与经典分类学结果基本一致,先与牛聚为一类,再与野猪、马等哺乳动物聚类,最后才与原鸡聚为一类,可见EphA3基因在不同物种间的进化关系与物种本身进化模式相似。因此,推断该基因在不同物种中具有相似的生物学功能,为后续研究该基因在毛发卷曲的生物学机制中的作用提供依据。通过对EphA3蛋白结构分析发现其富含α-螺旋及随机卷曲结构,而自由卷曲经常构成酶活性部位和其他蛋白质特异性功能部位;此外,跨膜区所对应的序列位置正好存在α-螺旋,这说明该蛋白的跨膜结构域可能是由1个或多个疏水的α-螺旋组成,EphA类受体是单次跨膜蛋白,跨膜结构域是其发挥生物功能的重要结构,对滩羊EphA3基因结构的探索有助于解析该基因的生物学功能。EphA3基因在滩羊的皮肤组织中表达量最高,揭示该基因可能是影响毛发生长的重要基因之一;而EphA3基因在48月龄滩羊皮肤组织中的相对表达量显著高于1月龄,则表明EphA3基因可能在不同月龄滩羊卷曲被毛变化进程中具有不可忽视的作用。综上所述,推测EphA3基因通过一种负调控机制参与不同时期滩羊被毛卷曲性状的分子遗传机制,但还需要进一步的实验数据验证。
4 结 论
本研究通过对EphA3基因编码区序列的克隆及生物学信息学分析,揭示了其主要结构特征;通过组织表达谱及荧光定量分析,明确了EphA3基因在滩羊各组织中的表达模式及差异。结合上述研究结果,推测EphA3基因在滩羊1月龄和48月龄皮肤组织中的差异表达可能是调控滩羊不同时期被毛表型差异的重要基因之一,该研究可为后续明确EphA3基因调控滩羊卷曲被毛生长的生物学机理提供分子依据。
