甘蓝型油菜NRT1.5和NRT1.8家族基因的生物信息学分析及其对氮-镉胁迫的响应
2019-03-18梁桂红华营鹏宋海星张振华
梁桂红 华营鹏 周 婷 廖 琼 宋海星 张振华,*
甘蓝型油菜和家族基因的生物信息学分析及其对氮-镉胁迫的响应
梁桂红1,2华营鹏1,2周 婷1,2廖 琼1,2宋海星1,2张振华1,2,*
1湖南农业大学资源环境学院, 湖南长沙 410128;2南方粮油作物协同创新中心, 湖南长沙 410128
植物对硝酸盐的吸收和转运需要硝酸盐转运体(nitrate transporters, NRTs)的协助。在拟南芥中, 硝酸盐的长途转运及其在根部和地上部的分配, 主要受家族的两个成员和的协同调控, 且两者的表达均受到硝酸盐的强烈诱导。本文以和基因序列为基础序列, 采用生物信息学方法鉴定了白菜、甘蓝和甘蓝型油菜中和同源基因, 并对基因结构和分子特性、基因拷贝数变异、基因染色体分布、系统进化树、蛋白保守序列比对和跨膜结构域、基因响应低氮和镉胁迫的转录组测序以及基因共表达网络进行了分析。结果表明, 白菜、甘蓝及甘蓝型油菜中NRT1.5和NRT1.8蛋白均含有保守的跨膜结构域和保守基序(F-Y-L-A-L-N-L- G-S-L), 属于MFS (major facilitator superfamily)超家族的小肽转运体PTR (peptide transporter)家族。转录组测序结果表明, 甘蓝型油菜低氮处理72 h, 根部基因的表达丰度上调而抑制的表达; 镉处理条件下, 乙烯/茉莉酸-硝酸盐转运体介导的信号途径能够促进表达上调而抑制的表达, 从而使更多的硝酸盐从地上部运输到根部, 提高植物抗镉胁迫的能力。本研究为进一步了解甘蓝型油菜和家族基因的生物学功能及其对氮-镉胁迫的响应奠定基础, 同时为和家族基因在其他物种中的生物信息学研究提供参考。
甘蓝型油菜; 生物信息学;;; 硝酸盐; 镉
氮(nitrogen, N)是植物生长发育所必需的大量营养元素, 广泛参与生物体蛋白质和核酸等大分子化合物的合成, 在生物体的物质和能量代谢过程中起重要作用[1]。铵态氮(NH4+-N)和硝态氮(NO3–-N)是作物氮素吸收利用的主要形态, 其中硝态氮是旱地作物吸收利用的主要氮源[2]。植物从土壤中吸收硝酸盐是一个主动运输的过程, 其能量主要来源于质子电化学势梯度, 协助硝酸盐转运的载体称为硝酸盐转运体(nitrate transporters, NRTs)[3], 包括低亲和硝酸盐转运体(nitrate transporter 1)和高亲和硝酸盐转运体(nitrate transporter 2) 2个家族[4-5]。
硝酸盐被植物根系细胞吸收后, 在植物体内的分配主要有硝酸盐短途分配(液泡硝酸盐分配)和长途转运(硝酸盐在根部和地上部的转运)两种途径, 它们共同调控着硝酸盐的利用效率, 进而影响作物的氮素利用率(Nitrogen Use Efficiency, NUE)[6]。大部分的硝酸盐在蒸腾拉力的作用下经木质部长途转运, 将硝酸盐分配到地上部的不同组织和器官中被同化和利用。该过程主要由向上运输的木质部和向下运输的韧皮部介导[7]。有研究证明, 拟南芥木质部介导的硝酸盐长途转运及其在根和地上部的分配, 主要受家族的2个成员和的协同调控, 且二者的表达都受到硝酸盐的强烈诱导[8-12]。主要在根部原生木质部的中柱鞘细胞中表达, 负责将根部细胞质中的硝酸盐装载进入木质部以运往地上部分。当其功能缺失后, 突变体木质部伤流液中的硝酸盐含量降低, 植物向上转运硝酸盐的能力减弱[8]。在木质部薄壁细胞表达, 通过调控硝酸盐在木质部的卸载进而影响硝酸盐从根向地上部分的运输[10]。由此可见,和基因的表达模式相反, 但共同影响硝酸盐通过木质部向地上部分的长途转运, 进而影响作物的产量和氮素利用率。此外,和通过调控硝酸盐在地上部和根部的分配, 从而影响植物对镉胁迫的耐受能力。研究发现镉处理条件下,基因的表达显著上调, 抑制的表达, 使更多的硝酸盐从地上部运输到根部, 根部较高浓度的硝酸盐有利于增强植株对镉胁迫的耐受能力[10,13]。
在我国, 油菜作为一种重要的油料作物, 种植面积和总产量约占世界的1/3[14], 在有效供给油脂和改善生活结构等方面起着重要作用。甘蓝型油菜(L, AnAnCnCn, 约1345 Mb, 2= 4= 38)属于十字花科芸薹属[15], 由白菜(, ArAr, ~485 Mb, 2= 2= 20)[16]和甘蓝(CoCo, ~630 Mb, 2= 2= 18)[17]这2个二倍体基本种在7500年前通过天然远缘杂交形成, 其全基因组包含约十万多个蛋白编码基因[18-19]。甘蓝型油菜进化、遗传的复杂性形成了其复杂的基因组, 在一定程度上影响了各种功能基因在甘蓝型油菜生长和发育中的作用。
硝酸盐转运蛋白负责植物根系对硝酸盐的吸收和转运, 一直是植物营养学领域研究的热点之一。家族有53个家族成员, 目前, 其他作物中对于与硝酸盐长途转运的关系研究主要集中在小麦和水稻中, 但由于非模式植物研究的突变体材料和克隆基因技术的限制, 只在小麦中研究了和基因对氮胁迫的响应[20], 以及水稻中过表达材料的氮素生理特性[21], 对和基因的结构功能及发育进化分析较少[22], 尤其在经济作物甘蓝型油菜中的研究更少。
本研究利用和的基因序列和蛋白序列, 对甘蓝型油菜中和基因进行鉴定和分析, 通过对基因生物信息数据的挖掘和整理, 分析该基因在白菜、甘蓝和甘蓝型油菜中的系统进化关系和保守基序特征, 预测其蛋白质的理化性质等, 为进一步系统研究和基因在其他物种中的功能提供参考依据。
1 材料与方法
1.1 试验材料
水培试验所用的甘蓝型油菜(L)品种为“湘油15号”, 油菜种子来自国家油料作物改良中心湖南分中心官春云院士团队。
1.2 白菜、甘蓝及甘蓝型油菜NRT1.5和NRT1.8基因序列的数据来源及检索
以拟南芥(AT1G32450)和(AT4G21680)的基因序列为种子序列, 利用BLAST搜索白菜、甘蓝和甘蓝型油菜中和的目的基因序列, 同时下载筛选到的基因序列对应编码的氨基酸序列, 将其保存为FASTA格式。本研究所用的数据库包括拟南芥信息资源(TAIR)(https:// www.arabidopsis.org/)。白菜()、甘蓝()及甘蓝型油菜()基因组全长DNA序列、CDS序列、氨基酸序列及其注释信息均从BRAD (http://brassicadb.org/brad/)数据库下载[23]。胡杨的氨基酸序列从NCBI (https://www.ncbi.nlm.nih.gov/)在线数据库下载。高粱、水稻、粟、玉米等单子叶植物的氨基酸序列从JGI (https://phytozome.jgi.doe. gov/pz/portal.html#)在线数据库下载。
1.3 白菜、甘蓝及甘蓝型油菜的基因命名
对三种芸薹属作物基因的命名以物种名称+染色体(后缀.)+拟南芥中的同源基因。例如,代表甘蓝型油菜中定位在A5染色体上的基因。
1.4 白菜、甘蓝及甘蓝型油菜NRT1.5和NRT1.8蛋白的分子特性分析
采用ExPASy ProtoParam (https://web.expasy. org/protparam/)[24]在线软件预测白菜、甘蓝和甘蓝型油菜NRT1.5和NRT1.8蛋白的氨基酸数目和组成、相对分子量(MW)、理论等电点(pI)、蛋白质的亲水性(GRAVY)及稳定性等理化性质。
1.5 白菜、甘蓝及甘蓝型油菜NRT1.5和NRT1.8基因的内含子-外显子结构特征
从拟南芥信息资源(TAIR)和BRAD数据库中下载拟南芥、白菜、甘蓝和甘蓝型油菜的全长基因组DNA序列和编码序列(CDS), 利用在线软件GSDS 2.0 (http://gsds.cbi.pku.edu.cn/)描绘内含子和外显子结构[25]。
1.6 白菜、甘蓝及甘蓝型油菜NRT1.5和NRT1.8基因的染色体定位分析
利用BLAST分别确定和在白菜、甘蓝和甘蓝型油菜中每条染色体上的起始位置, 并在GBrowse (http://brassicadb.org/cgi-bin/gbrowse/ Brassica/)中查找4个物种的各条染色体全长。利用软件MapInspect v. 2010 (http://www.softsea.com/ review/MapInspect.html)绘制和在白菜、甘蓝及甘蓝型油菜中染色体的分布情况。一般认为, 在100-kb基因组范围内排列有2个或以上基因, 我们定义其为串联重复基因[26]。
1.7 白菜、甘蓝及甘蓝型油菜NRT1.5和NRT1.8的系统发育关系
提取3种芸薹属作物NRT1.5和NRT1.8蛋白的氨基酸序列, 利用软件MEGA 7.0 (http://www. megasoftware.net/)[27]分别进行多重比对分析, 去除冗余序列[28]。采用邻接法(Neighbor Joining, NJ)[29]对白菜、甘蓝和甘蓝型油菜NRT1.5和NRT1.8的比对蛋白序列构建系统发育树, 并进行自检, 设定Bootstrap重复为1000, 去除bootstrap支持率低于50%的节点。
1.8 白菜、甘蓝及甘蓝型油菜NRT1.5和NRT1.8蛋白的进化选择压力分析
为了研究NRT1.5和NRT1.8蛋白在物种进化过程中是否涉及到达尔文的选择, 分别对拟南芥和白菜、甘蓝和甘蓝型油菜中NRT1.5和NRT1.8的CDS序列和蛋白序列进行比对, 利用在线软件PAL2NAL (http://www.bork.embl.de/pal2nal/)计算核苷酸的同义突变频率(synonymous, Ks)和非同义突变频率(non-synonymous, Ka), 及其比值Ka/Ks。
1.9 白菜、甘蓝及甘蓝型油菜NRT1.5和NRT1.8蛋白的保守基序和跨膜结构域预测分析
为了研究拟南芥和3种芸薹属作物中NRT1.5和NRT1.8蛋白的结构差异, 利用软件MEME v. 4.12.0 (http://meme-suite.org/tools/meme)[30]对蛋白质的保守基序进行预测和分析。所用默认参数, 最佳基序宽度为6~50 bp, 基序最大数为15。利用在线软件NovoPro (http://www.novopro.cn/tools/color_align_ prop.html)进行蛋白保守序列分析。同时利用在线软件TMpred Server (https://embnet.vital-it.ch/software/ TMPRED_form.html)对甘蓝型油菜的NRT1.5和NRT1.8蛋白序列进行跨膜结构域的预测和分析[31]。
1.10 甘蓝型油菜NRT1.5和NRT1.8基因在氮-镉胁迫条件下的表达分析
1.10.1 甘蓝型油菜的培养条件 油菜苗期培养试验在光照培养室中, 其温度22℃, 光照周期14 h (光照)/10 h (黑暗), 光照强度300~320 μmol m–2s–1, 湿度60%~75%。选取大小一致的油菜种子, 使用75%的酒精杀菌处理10 min, 用超纯水将种子表面冲洗干净后, 4℃下用无菌水浸泡24 h, 期间换水5~6次。将浸泡后的种子均匀播种在塑料育苗盘表面固定的纱布上, 育苗盘中加适量超纯水。育苗7 d后, 将长势一致的幼苗移栽到盛有4 L营养液的黑色塑料盒中。营养液浓度为Hoagland完全营养液[32]的1/4。
1.10.2 低氮处理高通量转录组测序试验 油菜幼苗以硝态氮为唯一氮源, 在6.0 mmol L–1NO3–中培养10 d后, 转移至0.3 mmol L–1NO3–条件下处理0、3和72 h, 对地上部(S)和根(R)取样, 将每个样品单独分装到干净的离心管中, 并作好标记, 在液氮中速冻, –80℃冰箱中保存待测, 每个处理作3个独立的生物学重复。测序平台为Illumina Hiseq 4000[33], 每个样品产生6.0 Gb测序数据, 两端配对末端(PE)读数, 由RNA-seq数据确定[34]转录组丰度(FPKM值)。
1.10.3 镉处理高通量转录组测序试验 油菜幼苗在无镉的正常营养液中培养10 d后, 将油菜样品分成两组, 一组继续在正常营养液中培养, 另一组转移至含10 μmol L–1镉的营养液中处理6 h, 分别对两组样品的地上部(S)和根(R)取样, 将每个样品单独分装到干净的离心管中, 作好标记。样品的高通量转录组测序试验参照步骤1.10.2。
2 结果与分析
2.1 白菜、甘蓝及甘蓝型油菜NRT1.5和NRT1.8基因/蛋白的分子特性
利用和数据在甘蓝型油菜全基因组数据库中分别检索到8条和8条同源基因序列(表1和表2)。白菜、甘蓝和甘蓝型油菜的NRT1.5蛋白均由22种氨基酸组成, 其中Leu和Ser的含量最高。基因编码区长度在1236 bp ()~1863 bp ()之间, 编码411~620个氨基酸, 预测蛋白质的理论分子量为45.80~69.41 kD。8个NRT1.5蛋白中酸性氨基酸(Asp+Glu)个数均多于碱性氨基酸(Arg+Lys), 等电点(pI)范围在5.63~6.35, 为酸性蛋白。Guruprasad等[35]认为, 蛋白质的不稳定系数大于40时为不稳定蛋白。NRT1.5蛋白的不稳定性系数在27.54~34.68间, 均小于40, 属于稳定性蛋白。氨基酸的亲水性指数(GRAVY)是氨基酸的化学特性, 是描述其支链的亲水性或疏水性程度的值, 根据亲水性指数介于–0.5~0.5为两性蛋白(GRAVY为负值表示亲水性, 正值表示疏水性)的原则[36], 氨基酸的亲水性指数在–0.006 (BolC5.NRT1.5b)~0.096 (BraA5.NRT1.5)之间, 表明NRT1.5主要为两性蛋白。脂肪指数在86.13 (BolC5.NRT1.5b)~89.77 (BnaC5.NRT1.5b)之间。
NRT1.8蛋白在白菜、甘蓝和甘蓝型油菜中均由22种氨基酸组成。其中除了BraA3.NRT1.8、BolC6.NRT1.8和BnaA3.NRT1.8三种蛋白的Leu和Ala含量较高外, 其余五种蛋白均是Leu和Ser的含量较高。除(897 bp)和(1167 bp)外,的基因编码区均为1752 bp, 编码583个氨基酸, 分子量分布在64.55 kD (BraA3. NRT1.8)~64.72 kD (BraA1.NRT1.8)之间。3个NRT1.8蛋白(BolC6.NRT1.8、BnaA3.NRT1.8和BnaC7.NRT1.8)中Arg+Lys>Asp+Glu, pI分别为8.18、7.52和8.18, 为碱性蛋白; BraA3.NRT1.8中Arg+Lys和Asp+Glu均为51, pI为7.06, 为中性蛋白; 其余4个NRT1.8蛋白中Asp+Glu>Arg+Lys, 为酸性蛋白。蛋白的不稳定性系数在18.64 (BolC6.NRT1.8)~ 32.07 (BnaCn.NRT1.8)之间, 为稳定性蛋白。GRAVY介于0.088 (BolC6.NRT1.8)~0.218 (BraA3.NRT1.8)之间, 为双亲和蛋白。脂肪指数在87.96 (BnaCn. NRT1.8)~94.85 (BnaC7.NRT1.8)之间。本研究中3种芸薹属作物NRT1.5和NRT1.8蛋白中Leu的含量均较高, 均为稳定的双亲和蛋白。

表1 白菜、甘蓝和甘蓝型油菜NRT1.5和NRT1.8基因的分子特性
(续表1)

基因名Gene name基因编号Gene ID分区Block亚类Subgenome物理位置Physical position编码区长度CDS (bp)外显子/内含子Exon/intron BnaC5.NRT1.5aBnaC05g28620DBMF226898087–2690321318456/5 BnaC5.NRT1.5bBnaC05g24580DBLF19037761–1904190018606/5 NRT1.8 BraA1.NRT1.8Bra013547ULF6243271–624529817524/3 BraA3.NRT1.8Bra038763UMF124230588–2423259417524/3 BolC1.NRT1.8Bol028440ULF8086050–808807517524/3 BolC6.NRT1.8Bol024295UMF142081659–420827298973/2 BnaA1.NRT1.8BnaA01g11510DULF5715790–571812617524/3 BnaA3.NRT1.8BnaA03g44820DUMF122774854–2277686217524/3 BnaC7.NRT1.8BnaC07g36810DUMF138737319–3873932517524/3 BnaCn.NRT1.8BnaCnng78690DULF80520183–8052181611673/2
表2 白菜、甘蓝和甘蓝型油菜NRT1.5和NRT1.8蛋白的分子特性
2.2 白菜、甘蓝、甘蓝型油菜及拟南芥NRT1.5和NRT1.8的基因拷贝数变异、染色体定位及基因结构分析
以拟南芥和的基因序列为基础序列, 对在油菜全基因组数据库中检索到的8个和8个同源基因的拷贝数变异、染色体定位及基因结构特征进行如下分析。
拷贝数变异(copy number variations, CNVs)是由基因组发生重排所致。为了比较白菜、甘蓝及甘蓝型油菜和基因的进化多样性, 分别以和为查询对象, 在NCBI数据库中进行BLAST搜索。图1显示, 在异源四倍体甘蓝型油菜中鉴定了4个和4个同源基因, 明显多于拟南芥、甘蓝和白菜所具有的和基因数量。甘蓝型油菜中基因的数量与甘蓝和白菜中的总数相同,基因的数量等同于甘蓝和白菜中的总数, 这意味着甘蓝型油菜和基因在异源多倍体自然加倍过程中没有损失。
利用MapInspect软件绘制出和基因在拟南芥、白菜、甘蓝及甘蓝型油菜染色体上的定位图(附图1)。表明和基因分布在不同的染色体上。基因在染色体上的分布是不均匀的, 每条染色体上有1~2个基因。Ar5、Ar9、An5和An9各分布1个; Co5和Cn5各分布2个串联重复基因。异源四倍体甘蓝型油菜中的A染色体全部来源于亲本二倍体白菜, C染色体全部来源于亲本二倍体甘蓝, 说明NRT1.5蛋白在自然加倍的进化过程中相对保守。与基因在染色体上的分布相比,基因在芸薹属作物染色体上的分布相对均匀, Ar1和Ar3、Co1和Co6以及An1、An3和Cn7每条染色体上各有一个基因。
外显子–内含子结构的数目是某些基因家族中典型的进化印记[37]。因此, 利用GSDS 2.0软件对芸薹属作物和拟南芥中和的基因结构进行分析(图2)。白菜和甘蓝的基因均有4个内含子, 甘蓝型油菜有5个内含子, 与拟南芥的基因结构类似。除和有2个内含子外, 3种芸薹属作物基因均含3个内含子。同源基因中内含子个数的不同在一定程度上表明了即使是直系同源基因也有可能经历基因结构的多样化。

图1 白菜、甘蓝、甘蓝型油菜及拟南芥NRT1.5和NRT1.8基因的拷贝数变异
柱状图顶部数值为该物种拷贝的基因数目。
The number at the top of the histogram is the number of genes copied for that species.

图2 白菜、甘蓝、甘蓝型油菜及拟南芥NRT1.5和NRT1.8的基因结构特征
图a表示拟南芥、白菜、甘蓝及甘蓝型油菜的基因结构特征; 图b表示拟南芥、白菜、甘蓝及甘蓝型油菜的基因结构特征。其中, 黄色部分代表CDS序列, 蓝色部分代表上游基因和下游基因, 黑色细线代表基因的内含子。
Fig. a and Fig. b show the gene structure characteristics ofandrespectively in,,,andThe yellow parts represent the CDS sequence and the blue parts represent the upstream and downstream genes. The black lines represent the intron of genes.
2.3 不同物种NRT1.5和NRT1.8蛋白的系统进化分析
为了比较NRT1.5和NRT1.8蛋白在不同物种发育进化过程中的分子进化规律和系统发生关联, 分别对拟南芥、甘蓝型油菜、胡杨等5个双子叶物种和水稻、玉米、高粱等5个单子叶物种的32个NRT1.5和33个NRT1.8蛋白序列进行同源性比对, 利用邻接法构建系统进化树(图3)。图3所示, NRT1.5和NRT1.8家族成员均分为双子叶植物和单子叶植物两个分支, 表明NRT1.5和NRT1.8这两种蛋白均在生物形态发生后开始出现差异[36]。白菜、甘蓝和油菜中NRT1.5和NRT1.8两种蛋白均位于双子叶的分支, 且非常紧密地聚集在一起, 同源性较高。
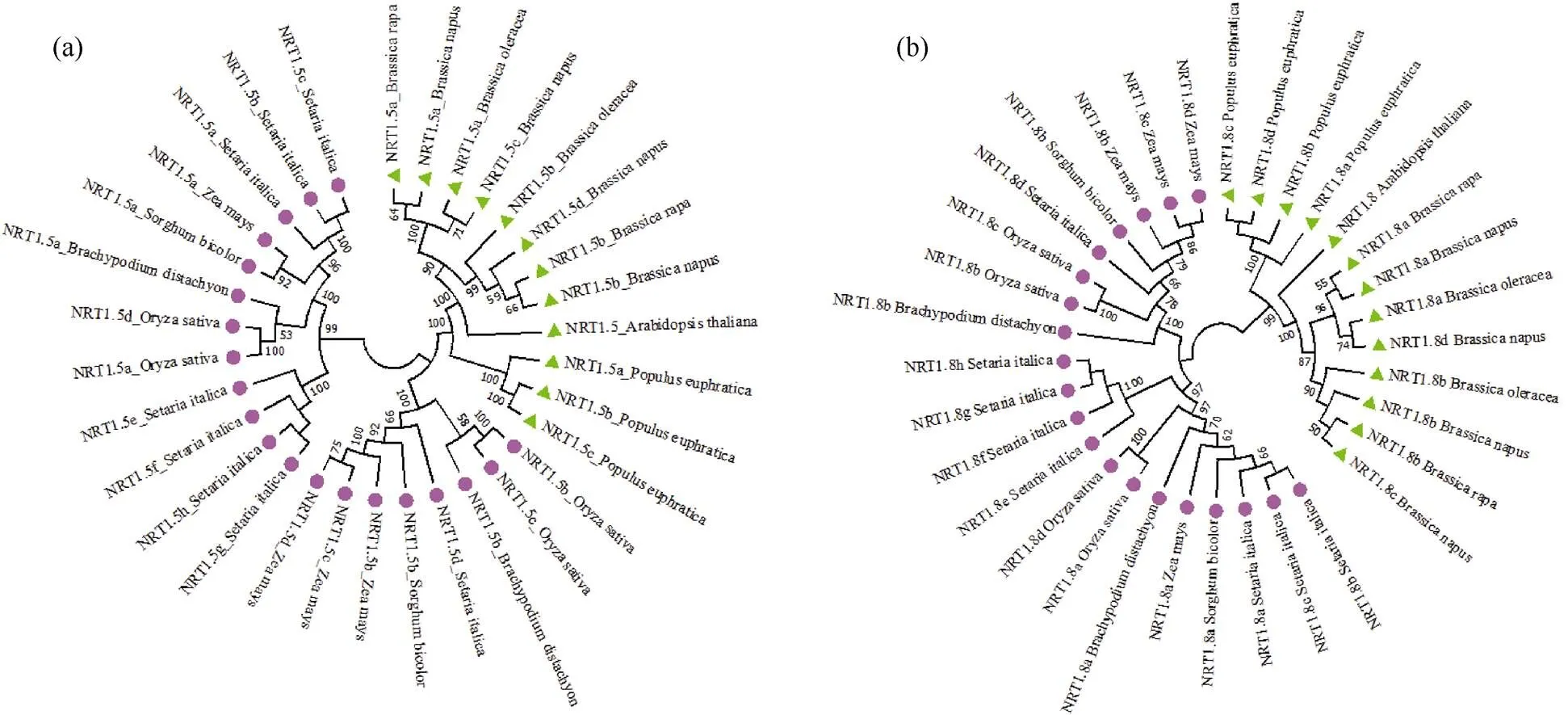
图3 不同物种NRT1.5和NRT1.8蛋白的系统发育关系
图a表示在双子叶和单子叶植物中NRT1.5蛋白的系统发育关系; 图b表示在双子叶和单子叶植物中NRT1.8蛋白的系统发育关系。其中, 绿色部分代表双子叶植物, 包括拟南芥、白菜、甘蓝、甘蓝型油菜和胡杨, 红色部分代表单子叶植物, 包括高粱、水稻、小米、玉米和二穗短柄草。
Fig.a and Fig.b show the phylogenetic relationships of NRT1.5 and NRT1.8 proteins respectively in dicots and monocotsThe green parts represent the dicots, including,andThe red parts represent the monocots, including,,,, and.
2.4 白菜、甘蓝及甘蓝型油菜NRT1.5和NRT1.8蛋白的进化选择压力分析
十字花科芸薹属作物全基因组可分为LF, MF1和MF2[38]3个亚基因组。表1显示, NRT1.5蛋白定位在B分区, 其中50%为LF亚基因组, 50%为MF2亚基因组。NRT1.8蛋白定位在U分区, 分为LF和MF1两个亚基因组。
为了研究NRT1.5和NRT1.8蛋白在物种进化过程中是否受到选择压力, 分别对拟南芥和3种芸薹属作物NRT1.5和NRT1.8的CDS序列及蛋白序列进行比对, 分别计算Ka、Ks和Ka/Ks。在遗传学中认为, 用Ka/Ks可以判断这个基因在进化中是否受到选择压力。当Ka/Ks>1.0时, 认为有正选择效应; 当Ka/Ks=1.0时, 认为该蛋白存在中性选择; 当Ka/Ks<1.0时, 则认为存在纯化选择作用[39]。图4所示, NRT1.5和NRT1.8在3种芸薹属作物中的Ka/Ks值均小于1.0, 表明这两种蛋白在物种进化过程中保留其重要功能而存在纯化选择作用。
2.5 白菜、甘蓝、甘蓝型油菜及拟南芥NRT1.5和NRT1.8蛋白的保守基序分析
不同物种硝酸盐转运体的蛋白序列高度同源, 具有相似的保守序列。在拟南芥中,家族属于MFS (major facilitator superfamily)的小肽转运体PTR (peptide transporter)家族。PTR转运蛋白一般含有450~600个氨基酸和12个跨膜区。在第5跨膜区有一个PTR家族的保守序列(F-Y-x-x-x-N-x-G-S- L)[40]。对3种芸薹属作物及拟南芥NRT1.5和NRT1.8的氨基酸序列比对发现, 4个物种NRT1.5和NRT1.8蛋白的氨基酸序列相似性很高, 蛋白结构保守, 均含有PTR家族的保守基序(F-Y-L-A-L-N-L-G-S- L)(图5-a和图5-b蓝色虚线区域)。利用在线软件MEME对芸薹属作物及拟南芥NRT1.5和NRT1.8蛋白的保守基序进行分析, 不同颜色的盒子表示不同的保守基序, 灰色的线条表示没有检测到基序的蛋白质区域。图6-a显示, NRT1.5蛋白在4个物种中相对保守, 保守基序为10~15个。其中, 甘蓝型油菜和拟南芥中NRT1.5蛋白非常保守, 保守基序均为15个; BraA9.NRT1.5、BraA5.NRT1.5和BolC5.NRT1.5a缺失Motif 1, 含有14个保守基序; BolC5.NRT1.5b有10个保守基序, 缺失Motif 1、Motif 9、Motif 10、Motif 13及Motif 14。图6-b所示, 4个物种NRT1.8蛋白的保守基序为7~15个, 均含有保守基序Motif 6和Motif 7。其中BraA1.NRT1.8、BraA3.NRT1.8、BnaA1.NRT1.8、BnaA3.NRT1.8、BnaC7.NRT1.8及BolC1.NRT1.8的保守基序为15个; AtNRT1.8、BnaCn.NRT1.8和BolC6.NRT1.8保守基序分别为14、10和7个。从分子进化的角度看, 氨基酸残基在进化过程中的保守性, 可以认为其在功能或结构上具有重要性[41]。

图4 白菜、甘蓝及甘蓝型油菜中NRT1.5和NRT1.8蛋白的同义突变频率(Ks)和非同义突变频率(Ka)
图a, b, c分别表示白菜、甘蓝及甘蓝型油菜NRT1.5蛋白的同义突变频率和非同义突变频率; 图d, e, f分别表示白菜、甘蓝及甘蓝型油菜NRT1.8蛋白的同义突变频率和非同义突变频率;
Fig. a, Fig. b, and Fig. c show synonymous nucleotide substitution rates and non-synonymous nucleotide substitution rates of NRT1.5 proteins in,, and,respectively. Fig. d, Fig. e, and Fig. f show synonymous nucleotide substitution rates and non-synonymous nucleotide substitution rates of the NRT1.8 proteins in,, and, respectively.
图a表示拟南芥、白菜、甘蓝及甘蓝型油菜NRT1.5蛋白的保守序列比对; 图b表示拟南芥、白菜、甘蓝及甘蓝型油菜NRT1.8蛋白的保守序列比对。其中, 蓝色虚线部分表示PTR家族的保守序列(F-Y-L-A-L-N-L-G-S-L)。
Fig. a and Fig. b show the conservative sequence alignment of NRT1.5 and NRT1.8 proteins respectively in,,, and. The blue parts show the conservative sequence (F-Y-L-A-L-N-L-G-S-L) of peptide transporter.
2.6 甘蓝型油菜NRT1.5和NRT1.8蛋白跨膜结构域分析
拟南芥家族的蛋白含12个跨膜区, 且在第6、7跨膜区之间有一个由约100个氨基酸组成的大亲水环相连, 将12个跨膜区分成两部分[42]。BnaNRT1.5和BnaNRT1.8家族蛋白的跨膜结构域如图7所示, 除了BnaCn.NRT1.8含有7个跨膜结构域外(图7-h), NRT1.5和NRT1.8均含有12个跨膜结构域, 在第6~7跨膜区间由一个大的亲水环相连, 与拟南芥中家族的结构特点相似。NRT1.5蛋白跨膜区主要分布在第41~580氨基酸残基间, 12个跨膜结构域长度为17~29个氨基酸。BnaA1.NRT1.8、BnaA3.NRT1.8和BnaC7.NRT1.8蛋白的跨膜区位于第35~565个氨基酸残基间, 跨膜结构域长度为17~27个氨基酸。BnaCn.NRT1.8蛋白的跨膜区分布在260~370氨基酸之间, 跨膜结构域长度为17~22个氨基酸。
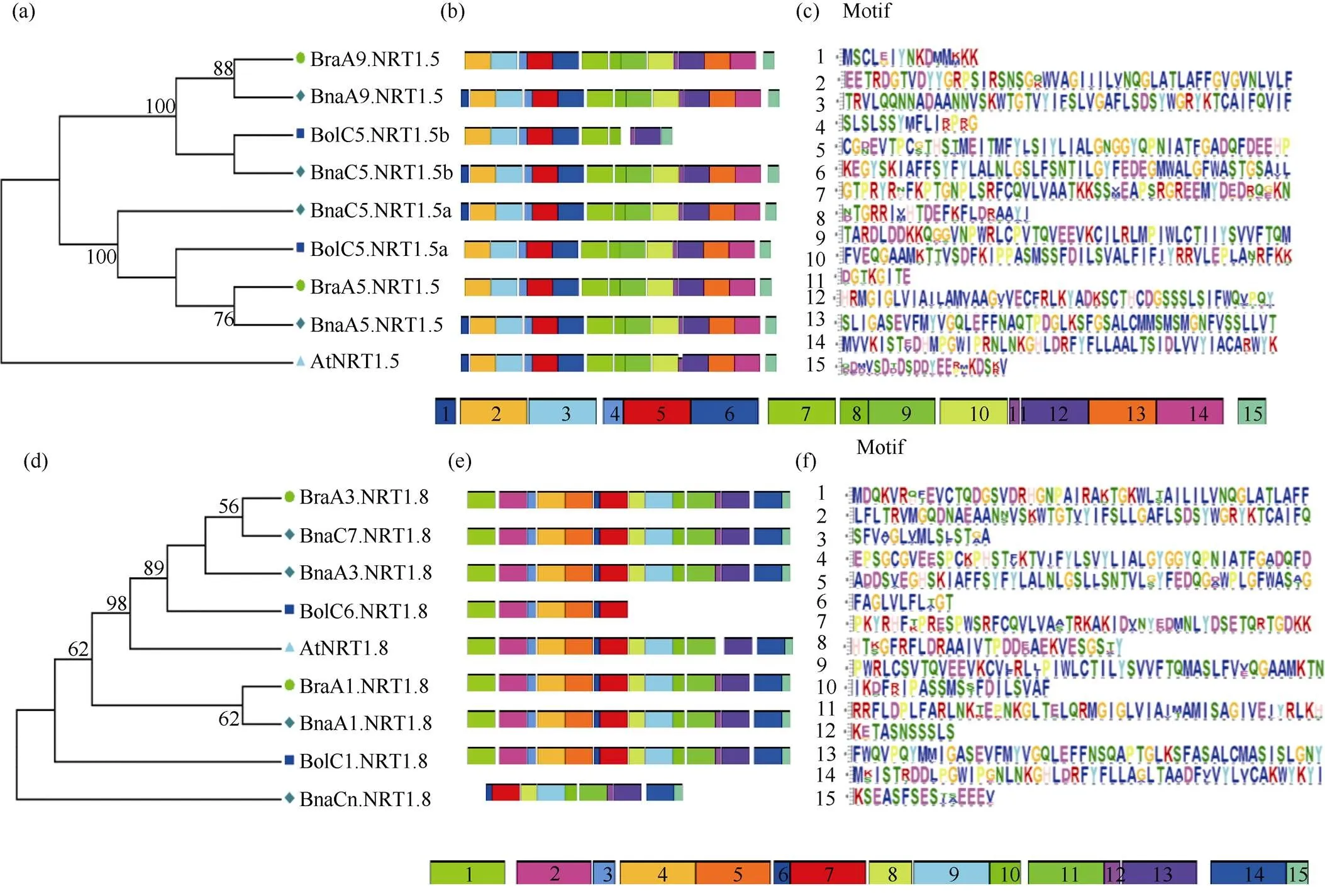
图6 白菜、甘蓝、甘蓝型油菜及拟南芥NRT1.5和NRT1.8蛋白的保守基序特征
图a, d分别表示NRT1.5和NRT1.8蛋白在拟南芥、白菜、甘蓝及甘蓝型油菜中的系统进化关系; 图b, e分别表示NRT1.5和NRT1.8蛋白在拟南芥、白菜、甘蓝及甘蓝型油菜的保守基序; 图c, f分别表示NRT1.5和NRT1.8蛋白中每个保守基序的序列。
Fig. a and Fig. d show the phylogenetic relationships of NRT1.5 and NRT1.8 proteins respectively in,,, and. Fig. b and Fig. e show the motives of NRT1.5 and NRT1.8 proteins respectively in,,, and. Fig. c and Fig. f show the sequence of conserved motif of NRT1.5 and NRT1.8 proteins.
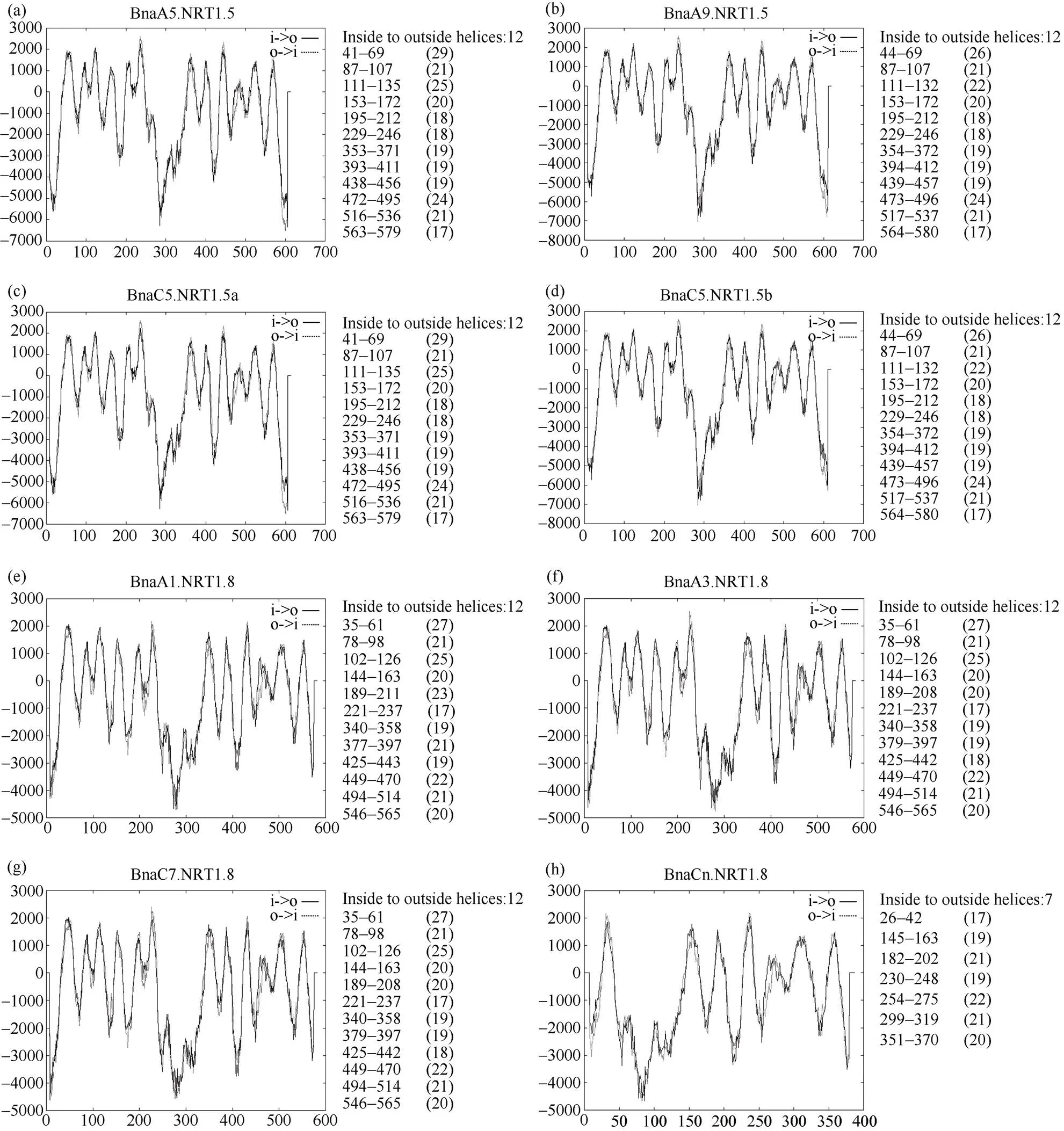
图7 甘蓝型油菜NRT1.5和NRT1.8蛋白的跨膜结构域
图a~d表示甘蓝型油菜中4个NRT1.5蛋白的跨膜结构域, 其中a表示Bna.A5.NRT1.5, b表示Bna.A9.NRT1.5, c表示Bna.C5.NRT1.5a, d表示Bna.C5.NRT1.5b。图e~h表示甘蓝型油菜中4个NRT1.8蛋白的跨膜结构域, 其中a表示Bna.A1.NRT1.8, f表示Bna.A3.NRT1.8, g表示Bna.C7.NRT1.8, h表示Bna.Cn.NRT1.8。
Fig. a to Fig. d show four transmembrane domains of NRT1.5 protein in. Fig. a, Fig. b, Fig. c, and Fig. d show the protein of Bna.A5.NRT1.5, Bna.A9.NRT1.5, Bna.C5.NRT1.5a, and Bna.C5.NRT1.5b, respectively. Fig. e to Fig. h show four transmembrane domains of NRT1.8 protein in. Fig. e, Fig.f, Fig. g, and Fig. h show the protein of Bna.A1.NRT1.8, Bna.A3.NRT1.8, Bna.C7.NRT1.8, and Bna.Cn.NRT1.8, respectively.
2.7 油菜NRT1.5和NRT1.8基因在氮-镉胁迫条件下基因表达丰度和基因共表达网络分析
硝酸盐不仅是重要的营养物质, 同时也是一种重要的信号分子, 对硝酸盐转运体发挥功能起重要的调控作用[43]。为了确定和对低氮胁迫是否存在响应, 对短期(3 h)和长期(72 h)低氮处理的甘蓝型油菜地上部和根进行了高通量转录组分析(图8)。表明低氮处理0~72 h后, 甘蓝型油菜中和基因在根部的表达均受到显著影响, 地上部差异不大。低氮处理后基因在根部的表达丰度上调, 且在72 h时达到最大值。其中,和表达量在0~3 h内上调, 3~72 h内趋于平缓。和表达量高, 且在0~72 h内持续上调。从甘蓝型油菜基因的共表达分析网络中可以看出,在响应低氮胁迫中均起到调控作用, 其中基因的调控占主导作用, 且与的功能互作较强。低氮处理同样会影响的表达。根部低氮处理0~72 h, 其基因表达量呈下调的趋势, 在0~3 h内急剧下调, 3~72 h内下调较缓慢, 在72 h表达量最低。其中, 低氮诱导相同的时间,和的表达量均高于。基因共表达网络分析也表明,和在响应低氮胁迫中均起调控作用, 其中前者起主导作用。
甘蓝型油菜湘油15号在10 μmol L–1的镉胁迫条件下, 根部和地上部基因的表达均受到抑制(图9-a, b), 基因共表达网络分析表明(图9-c),和在镉胁迫条件下均起到调控作用, 其中起主导调控作用。根部在镉胁迫下受到显著诱导,和在此过程中均起到调控作用, 其中起主要作用,和之间的功能互作较强(图9-d, e, f)。
甘蓝型油菜根部和基因在低氮和镉胁迫条件下的表达量均受到不同程度的诱导。低氮胁迫条件下,基因的表达丰度上调抑制基因的表达, 使更多的硝酸盐从根部运输到地上部被作物同化和利用, 从而提高植物的氮素利用率。植物受到镉胁迫时,基因的表达上调抑制基因的表达, 从而使更多的硝酸盐运输到根部, 根部高浓度的硝酸盐有利于提高植物的胁迫防御能力。

图8 甘蓝型油菜NRT1.5和NRT1.8基因对低氮的响应及基因共表达网络分析
图a表示甘蓝型油菜地上部和根部家族基因在低氮处理0 h、3 h、72 h 时的基因表达丰度; 图b表示家族基因在低氮处理时的基因共表达网络分析。图c表示甘蓝型油菜地上部和根部家族基因在低氮处理0 h、3 h、72 h 时的基因表达丰度; 图d表示家族基因在低氮处理时的基因共表达网络分析; FPKM表示每千个碱基转录每百万映射读取的fragments。
Fig. a shows the gene expression abundance offamily genes in shoots and roots ofwith low nitrate treatment for 0 h, 3 h, 72 h. Fig. b shows the co-expression network analysis offamily genes with low nitrate treatment. Fig. c shows the gene expression abundance offamily genes in shoots and roots ofwith low nitrate treatment for 0 h, 3 h, 72 h. Fig. d shows the co-expression network analysis offamily genes with low nitrate treatment. FPKM means the fragments per kilobase of exon model per million mapped reads.

图9 甘蓝型油菜NRT1.5和NRT1.8基因对镉胁迫的响应及基因共表达网络分析
图a和图b分别表示甘蓝型油菜地上部和根部家族基因在镉处理时的基因表达丰度; 图c表示家族基因在镉处理时的基因共表达网络分析。图d和图e分别表示甘蓝型油菜地上部和根部家族基因在镉处理时的基因表达丰度; 图f表示家族基因在镉处理时的基因共表达网络分析。FPKM表示每千个碱基转录每百万映射读取的fragments。图中所示显著性差异是单个基因对照和镉处理两两对比, 未达到显著性差异的图中未标记。误差线代表3个独立生物学重复的标准误, a和b表示差异显著(< 0.05)。
Fig. a and Fig. b show the gene expression abundance offamily genes in shoots and roots ofwith cadmium stress. Fig. c shows the co-expression network analysis offamily genes with cadmium stress. Fig. d and Fig. e show the gene expression abundance offamily genes in shoots and roots ofwith cadmium stress. Fig. f shows the co-expression network analysis offamily genes with cadmium stress. FPKM means the fragments per kilobase of exon model per million mapped reads. The significant difference shown in the figure is the pairwise comparison between control and cadmium treatment, and the figure without significant difference is not marked. Data are shown as means±SE (= 3) and the different lowercases above text are used to show statistical significance (< 0.05).
3 讨论
植物吸收和转运硝酸盐需要硝酸盐转运体(NRTs)的协助完成。在拟南芥中,家族有53个成员, 目前鉴定有8个成员()在硝酸盐吸收和转运中各自发挥着独特的作用[3,8,42,44-45], 其中和在调控硝酸盐长途转运及其根系和地上部的分配中起着重要的作用。白菜、甘蓝和甘蓝型油菜作为十字花科芸薹属中3种重要的经济作物, 对其和基因的结构功能及进化分析研究具有重要的实际意义。
拟南芥家族属于MFS (major facilitator superfamily)超家族的小肽转运体PTR (peptide transporter)家族。PTR转运蛋白是一类可以转运氨基酸、寡肽以及硝酸盐等含氮化合物的膜转运蛋白, 广泛存在于动物、植物和微生物中[46], 一般含450~ 600个氨基酸, 12个跨膜区, 在第6、第7跨膜区之间由一个约100个氨基酸组成的亲水环相连, 在第5跨膜区有一个PTR家族的保守基序(F-Y-x-x-x-N- x-G-S-L)。3种芸薹属作物中大部分NRT1.5和NRT1.8蛋白含411~620个氨基酸, 跨膜结构域分析显示有12个跨膜区, 且在第6~7跨膜区间有一个亲水环。蛋白保守序列比对和保守基序分析发现, 2个蛋白均含有PTR家族的保守基序(F-Y-L-A-L-N-L- G-S-L), 与拟南芥家族基因相似。一般认为, 含有多个内含子的基因比较保守, 而不含有内含子的基因保守性较差[47-48]。3种芸薹属作物和基因结构含有2~5个内含子, 分别属于B分区不同亚类(LF、MF2)和U分区不同亚类(LF、MF1)。系统进化树和Ka/Ks表明蛋白在不同物种发育进化过程中的同源性和进化过程中是否受到选择压力[39]。芸薹属作物NRT1.5蛋白同源性较高, Ka/Ks值均小于1.0, 表明NRT1.5在3个物种进化过程中存在纯化选择作用且发挥相似的功能。NRT1.8蛋白Ka/Ks值均小于1.0, 但同源性较低, 表明NRT1.8在物种系统进化中受到纯化选择压力, 但在不同物种进化过程中其功能可能存在一定差异。
硝酸盐是氮素储存和运输的重要形式, 同时作为一种重要的信号物质对硝酸盐转运体起到重要的调控作用。植物通过硝酸盐转运体感知土壤中硝酸盐的浓度, 诱导硝酸盐转运体家族的不同成员发挥作用[43]。Wang等[49]指出, 在不同氮水平下小麦根系硝酸盐转运蛋白的不同成员基因表达存在差异, 从而使植株更有效地利用养分。此外, 硝酸盐转运体在植物胁迫防御方面发挥着重要作用。研究表明, 在逆境胁迫条件下,和通过调控硝酸盐在根部和地上部的分配从而影响植物对逆境胁迫的耐受能力。乙烯/茉莉酸-硝酸盐转运体介导的信号途径通过促进表达上调而抑制的表达, 从而使更多的硝酸盐从地上部运输到根部, 进而提高植物抗逆境胁迫的能力[10-11]。本研究中, 油菜湘油15号根部在低氮处理0~72 h时,基因的表达丰度显著上调, 而基因的表达丰度呈现下调趋势, 由此可见,和两个基因在响应低氮条件下诱导产生的基因表达模式相反,但共同影响硝酸盐通过木质部向地上部的长途转运, 进而影响作物的产量和氮素利用率。在10 μmol L–1镉处理条件下, 甘蓝型油菜中表达上调抑制基因的表达, 这与Li等[10]研究的拟南芥基因对镉胁迫的响应一致。显著上调使更多的硝酸盐从地上部分运输到根部, 根部较高浓度的硝酸盐有利于根系适应镉胁迫。Li等[10]的研究也表明, 随着培养液中镉浓度的适当增加,中的硝酸盐含量较野生型显著降低, 这说明在根部运输硝酸盐的过程中,基因的表达受镉的正向调控。基因共表达网络分析发现, 油菜在适应低氮胁迫时是和起主要调控作用, 镉胁迫时发挥主导作用的是和, 这可能是因为和基因对氮和镉胁迫的响应程度不同, 由此可以推断这2个基因家族在抵抗非生物胁迫过程中可能扮演着不同的角色, 但不可否认的是和基因在植物适应氮-镉胁迫方面发挥着重要的调控作用。因此, 深入全面地了解和的基因结构和进化关系对更好地发挥其功能具有重要的意义, 同时也为这2个家族基因在其他物种中的功能研究提供依据。
[1] 李建勇, 龚继明. 植物硝酸根信号感受与传导途径. 植物生理学报, 2011, 47: 111–118. Li J Y, Gong J M. Nitrate signal sensing and transduction in higher plants., 2011, 47: 111–118 (in Chinese with English abstract).
[2] Tang Y, Sun X C, Hu C X, Tan Q L, Zhao X H. Genotypic differences in nitrate uptake, translocation and assimilation of two Chinese cabbage cultivars [L. ssp.(L.)]., 2013, 70: 14–20.
[3] Fan S C, Lin C S, Hsu P K, Lin S H, Tsay Y F. Thenitrate transporter, expressed in phloem, is responsible for source-to-sink remobilization of nitrate., 2009, 21: 2750–2761.
[4] Orsel M, Chopin F, Leleu O, Smith S J, Krapp A, Daniel-Vedele F, Miller A J. Characterization of a two-component high-affinity nitrate uptake system in. Physiology and protein- protein interaction., 2006, 142: 1304–1317.
[5] Møller A L, Pedas P, Andersen B, Svensson B, Schjoerring J K, Finnie C. Responses of barley root and shoot proteomes to long-term nitrogen deficiency, short-term nitrogen starvation and ammonium., 2011, 34: 2024–2037.
[6] 张振华. 作物硝态氮转运利用与氮素利用效率的关系. 植物营养与肥料学报, 2017, 23: 217–223. Zhang Z H. The relationship between nitrate transport and utilization in crop and nitrogen utilization efficiency., 2017, 23: 217–223 (in Chinese with English abstract).
[7] Dechorgnat J, Nguyen C T, Armengaud P, Jossier M, Diatloff E, Filleur S, Daniel-Vedele F. From the soil to the seeds: the long journey of nitrate in plants., 2011, 62: 1349–1359.
[8] Lin S H, Kuo H F, Canivenc G, Lin C S, Lepetit M, Hsu P K, Tillard P, Lin H L, Wang Y Y, Tsai C B, Gojon A, Tsay Y F. Mutation of thenitrate transporter causes defective root-to-shoot nitrate transport., 2008, 20: 2514–2528.
[9] Chen C Z, Lv X F, Li J Y, Yi H Y, Gong J M.is another essential component in the regulation of nitrate reallocation and stress tolerance., 2012, 159: 1582–1590.
[10] Li J Y, Fu Y L, Pike S M, Bao J, Tian W, Zhang Y, Chen C Z, Zhang Y, Li H M, Huang J, Li L G, Schroeder J I, Gassmann W, Gong J M. Thenitrate transporterfunctions in nitrate removal from the xylem sap and mediates cadmium tolerance., 2010, 22: 1633–1646.
[11] Zhang G B, Yi H Y, Gong J M. Theethylene/ jasmonic acid-NRT signaling module coordinates nitrate reallocation and the trade-off between growth and environmental adaption., 2014, 26: 3984–3998.
[12] Léran S, Varala K, Boyer J C, Chiurazzi M, Crawpord N, Daniel-Vedele F, David L, Dickstein R, Fernandez E, Forde B, Gassmann W, Geiger D, Gojon A, Gong J M, Halkier B A, Harris J M, Hedrich R, Limami A M, Rentsch D, Seo M, Tsay Y F, Zhang M, Coruzzi G, Lacombe B. A unified nomenclature of nitrate transporter 1/peptide transporter family members in plants., 2014, 19: 5–9.
[13] 宋田丽, 周建建, 徐晨曦, 蔡晓锋, 戴绍军, 王全华, 王小丽. 植物硝酸盐转运蛋白功能及表达调控研究进展. 上海师范大学学报(自然科学版), 2017, 46: 740–750. Song T L, Zhou J J, Xu C X, Cai X F, Dai S J, Wang Q H, Wang X L. Progress in function and regulation of nitrate transporters in plants.(Nat Sci), 2017, 46: 740–750 (in Chinese with English abstract).
[14] 殷艳, 陈兆波, 余健, 王汉中, 冯中朝. 我国油菜生产潜力分析. 中国农业科技导报, 2010, 12(3): 16–21. Yin Y, Chen Z B, Yu J, Wang H Z, Feng Z C. Analysis of potential for rapeseed production in China., 2010, 12(3): 16–21 (in Chinese with English abstract).
[15] Blackshaw R E, Johnson E N, Gan Y T, May W E, McAndrew D W, Barthet V, McDonald T, Wispinski D. Alternative oilseed crops for biodiesel feedstock on the Canadian prairies., 2011, 91: 889–896.
[16] Wang X, Wang H, Wang J, Sun R, Wu J, Liu S, Bai Y, Mun J H, Bancroft I, Cheng F, Huang S, Li X, Hua W, Wang J, Wang X, Freeling M, Pires J C, Paterson A H, Chalhoub B, Wang B, Hayward A, Sharpe A G, Park B S, Weisshaar B, Liu B, Li B, Liu B, Tong C, Song C, Duran C, Peng C, Geng C, Koh C, Lin C, Edwards D, Mu D, Shen D, Soumpourou E, Li F, Fraser F, Conant G, Lassalle G, King G J, Bonnema G, Tang H, Wang H, Belcram H, Zhou H, Hirakawa H, Abe H, Guo H, Wang H, Jin H, Parkin I A, Batley J, Kim J S, Just J, Li J, Xu J, Deng J, Kim J A, Li J, Yu J, Meng J, Wang J, Min J, Poulain J, Wang J, Hatakeyama K, Wu K, Wang L, Fang L, Trick M, Links M G, Zhao M, Jin M, Ramchiary N, Drou N, Berkman P J, Cai Q, Huang Q, Li R, Tabata S, Cheng S, Zhang S, Zhang S, Huang S, Sato S, Sun S, Kwon S J, Choi S R, Lee T H, Fan W, Zhao X, Tan X, Xu X, Wang Y, Qiu Y, Yin Y, Li Y, Du Y, Liao Y, Lim Y, Narusaka Y, Wang Y, Wang Z, Li Z, Wang Z, Xiong Z, Zhang Z. The genome of the mesopolyploid crop species., 2011, 43: 1035–1039.
[17] Liu S, Liu Y, Yang X, Tong C, Edwards D, Parkin I A, Zhao M, Ma J, Yu J, Huang S, Wang X, Wang J, Lu K, Fang Z, Bancroft I, Yang T J, Hu Q, Wang X, Yue Z, Li H, Yang L, Wu J, Zhou Q, Wang W, King G J, Pires J C, Lu C, Wu Z, Sampath P, Wang Z, Guo H, Pan S, Yang L, Min J, Zhang D, Jin D, Li W, Belcram H, Tu J, Guan M, Qi C, Du D, Li J, Jiang L, Batley J, Sharpe A G, Park B S, Ruperao P, Cheng F, Waminal N E, Huang Y, Dong C, Wang L, Li J, Hu Z, Zhuang M, Huang Y, Huang J, Shi J, Mei D, Liu J, Lee T H, Wang J, Jin H, Li Z, Li X, Zhang J, Xiao L, Zhou Y, Liu Z, Liu X, Qin R, Tang X, Liu W, Wang Y, Zhang Y, Lee J, Kim H H, Denoeud F, Xu X, Liang X, Hua W, Wang X, Wang J, Chalhoub B, Paterson A H. Thegenome reveals the asymmetrical evolution of polyploid genomes., 2014, 5: 3930.
[18] Bayer P E, Hurgobin B, Golicz A A, Chen C K, Yuan Y, Lee H, Renton M, Meng J, Li R, Long Y, Zou J, Bancroft I, Chalhoub B, King G J, Batley J, Edwards D. Assembly and comparison of two closely relatedgenomes., 2017, 15: 1602–1610
[19] Chalhoub B, Denoeud F, Liu S, Parkin I A, Tang H, Wang X, Chiquet J, Belcram H, Tong C, Samans B, Corréa M, Da Silva C, Just J, Falentin C, Koh C S, Le Clainche I, Bernard M, Bento P, Noel B, Labadie K, Alberti A, Charles M, Arnaud D, Guo H, Daviaud C, Alamery S, Jabbari K, Zhao M, Edger P P, Chelaifa H, Tack D, Lassalle G, Mestiri I, Schnel N, Le Paslier M C, Fan G, Renault V, Bayer P E, Golicz A A, Manoli S, Lee T H, Thi V H, Chalabi S, Hu Q, Fan C, Tollenaere R, Lu Y, Battail C, Shen J, Sidebottom C H, Wang X, Canaguier A, Chauveau A, Bérard A, Deniot G, Guan M, Liu Z, Sun F, Lim Y P, Lyons E, Town C D, Bancroft I, Wang X, Meng J, Ma J, Pires J C, King G J, Brunel D, Delourme R, Renard M, Aury J M, Adams K L, Batley J, Snowdon R J, Tost J, Edwards D, Zhou Y, Hua W, Sharpe A G, Paterson A H, Guan C, Wincker P. Early allopolyploid evolution in the post-Neolithicoilseed genome., 2014, 345: 950–953.
[20] 轩红梅, 王永华, 魏利婷, 杨莹莹, 王利娜, 康国章, 郭天财. 小麦幼苗叶片中硝酸盐转运蛋白和家族基因对氮饥饿响应的表达分析. 麦类作物学报, 2014, 34: 1019–1028. Xuan H M, Wang Y H, Wei L T, Yang Y Y, Wang L N, Kang G Z, Guo T C. Transcription analysis of the genes encoding nitrate transporterandfamily in response to nitrogen starvation in wheat seedlings leaves., 2014, 34: 1019–1028 (in Chinese with English abstract).
[21] 马翠. 水稻硝酸盐转运蛋白基因和超量表达材料的功能鉴定. 南京农业大学硕士论文, 江苏南京, 2011.Ma C. Characteristics of Over-expression for Nitrate Transporter Genesandin Rice. MS Thesis of Nanjing Agricultural University, Nanjing, Jiangsu, China, 2011 (in Chinese with English abstract).
[22] 李红. 拟南芥转运蛋白NRT1.5/NPF7.3调控K+在木质部装载的分子机制研究. 中国农业大学博士学位论文, 北京, 2016. Li H. Mechanism analyses of NRT1.5/NPF7.3-mediated K+release into the xylem in. PhD Dissertation of China Agricultural University, Beijing, China, 2016 (in Chinese with English abstract).
[23] Wang X B, Wu J, Liang J L, Cheng F, Wang X W.database (BRAD) version 2.0: integrating and miningspecies genomic resources., 2015, 1–8.
[24] Gasteiger E, Hoogland C, Gattiker A, Duvaud S, Wilkins M R, Appel R D, Bairoch A. Protein identification and analysis tools on the ExPASy server. In: Walker J M ed. The Proteomics Protocols Handbook. Totowa, NJ, USA: Humana Press Inc, 2005. pp 571−607.
[25] Hu B, Jin J, Guo A Y, Zhang H, Luo J, Gao G. GSDS 2.0: an upgraded gene feature visualization server., 2015, 31: 1296–1297
[26] Smith T F, Waterman M S. Identification of common molecular subsequences., 1981, 147, 195–197.
[27] Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0., 2013, 30: 2725–2729.
[28] Larkin M A, Blackshields G, Brown N P, Chenna R, McGettigan P A, McWilliam H, Valentin F, Wallace I M, Wilm A, Lopez R, Thompson J D, Gibson T J, Higgins D G. Clustal W and clustal X version 2.0., 2007, 23: 2947–2948.
[29] Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees., 1987, 4: 406–425.
[30] Bailey T L, Boden M, Buske F A, Frith M, Grant C E, Clementi L, Ren J, Li W W, Noble W S. MEME SUITE: tools for motif discovery and searching., 2009, 37(Web Server issue): W202–W208.
[31] Hofmann K, Stoffel W. TMBase: a database of membrane spanning protein segments., 1993, 374: 166
[32] Hoagland D R, Arnon D I. The water culture method for growing plants without soil., 1950, 347: 1–32.
[33] Morin R D, Bainbridge M, Fejes A, Hirst M, Krzywinski M, Pugh T J, McDonald H, Varhol R, Jones S J, Marra M A. Profiling the HeLa S3 transcriptome using randomly primed cDNA and massively parallel short-read sequencing., 2008, 45: 81–94.
[34] Hua Y P, Zhou T, Xu F S. Genome-scale mRNA transcriptomic insights into the responses of oilseed rape (L.) to varying boron availabilities., 2017, 416: 205–225.
[35] Guruprasad K, Reddy B V, Pandit M W. Correlation between stability of a protein and its dipeptide composition: a novel approach for predicting in vivo stability of a protein from its primary sequence., 1990, 4: 155–161.
[36] 王占军, 金伦, 徐忠东, 欧祖兰. 麻风树基因的生物信息学分析. 生物学杂志, 2014, 31(4): 68–72. Wang Z J, Jin L, Xu Z D, Ou Z L. Bioinformatics analysis of genefrom., 2014, 31(4): 68–72 (in Chinese with English abstract).
[37] Hua Y P, Zhou T, Song H X, Guan C Y, Zhang Z H. Integrated genomic and transcriptomic insights into the two-component high-affinity nitrate transporters in allotetraploid rapeseed., 2018, 427: 245–268.
[38] Cheng F, Wu J, Fang L, Wang X. Syntenic gene analysis betweenand otherspecies., 2012, 3: 198.
[39] Nekrutenko A, Makova K D, Li W H. The Ka/Ks ratio test for assessing the protein-coding potential of genomic regions: an empirical and simulation study., 2002, 12: 198–202.
[40] Forde B G. Nitrate transporters in plants: structure, function and regulation., 2000, 1465: 219–235.
[41] Chakrabarti S, Bryant S H, Panchenko A R. Functional specificity lies within the properties and evolutionary changes of amino acids., 2007, 373: 801–810.
[42] Tsay Y F, Chiu C C, Tsai C B, Ho C H, Hsu P K. Nitrate transporters and peptide transporters., 2007, 581: 2290–2300.
[43] 尹辉, 牟书勇, 李冠. 植物硝酸盐转运体的功能及其调控. 南方农业学报, 2012, 43: 425–430. Yin H, Mu S Y, Li G. Function and regulation of nitrate transporters in plants., 2012, 43: 425–430 (in Chinese with English abstract).
[44] 汪进, 添先凤, 江昌俊, 李叶云. 茶树硝酸盐转运蛋白基因的克隆和表达分析. 植物生理学报, 2014, 50: 983–988. Wang J, Tian X F, Jiang C J, Li Y Y. Cloning and expression analysis of nitrate transporter gene in., 2014, 50: 983–988 (in Chinese with English abstract).
[45] Almagro A, Lin S H, Tsay Y F. Characterization of thenitrate transporterreveals a role of nitrate in early embryo development., 2008, 20: 3289–3299.
[46] 童依平, 蔡超, 刘全友, 李继云, 李振声. 植物吸收硝态氮的分子生物学进展. 植物营养与肥料学报, 2004, 10: 433–440. Tong Y P, Cai C, Liu Q Y, Li J Y, Li Z S. Recent advances in molecular biology of nitrate transporters in higher plants., 2004, 10: 433–440 (in Chinese with English abstract).
[47] Theissen G, Becker A, Di Rosa A, Kanno A, Kim J T, Münster T, Winter K U, Saedler H. A short history of MADS-box genes in plants., 2000, 42: 115–149.
[48] Kaufmann K, Melzer R, Theissen G. MIKC-type MADS-domain proteins: structural modularity, protein interactions and network evolution in land plants., 2005, 347: 183–198.
[49] Wang P, Wang Z L, Cai R G, Li Y, Chen X G, Yin Y P. Physiological and molecular response of wheat roots to nitrate supply in seedling stage., 2011, 10: 695–704.

附图1 NRT1.5和NRT1.8基因在白菜、甘蓝、甘蓝型油菜及拟南芥中的染色体定位
图a表示基因在拟南芥、甘蓝型油菜、甘蓝及白菜中的染色体定位;图b表示基因在拟南芥、甘蓝型油菜、甘蓝及白菜中的染色体定位。
Fig. a shows the chromosomal localization ofgenes in,and. Fig. b shows the chromosomal localization ofgenes inand.
Bioinformatics analysis and response to nitrate-cadmium stress ofandfamily genes in
LIANG Gui-Hong1,2, HUA Ying-Peng1,2, ZHOU Ting1,2, LIAO Qiong1,2, SONG Hai-Xing1,2, and ZHANG Zhen-Hua1,2,*
1College of Resource and Environment, Hunan Agricultural University, Changsha 410128, Hunan, China;2Southern Regional Collaborative Innovation Center for Grain and Oil Crops in China, Changsha 410128, Hunan, China
The absorption and transportation of nitrate in plants require the assistance of nitrate transporters (NRTs). The expression of two members of thefamily, includingandgenes was strongly induced by nitrateandregulated the long-distance transport and distribution of nitrate between roots and shoots in.andhomologous genes in,, andwere identified by bioinformatics with the basic sequences ofandand analyzed in gene structures and proteins molecular characterization, gene copy number variations, chromosome locations, evolutionary relationship tree, proteins conservative sequence alignment and the transmembrane domains.andresponsive to the low concentration nitrate and cadmium stress were also determined by transcriptome analysis and co-expression network analysis, showing that NRT1.5 and NRT1.8 proteins belong to major facilitator superfamily (MFS) and peptide transporter (PTR) with the conservative transmembrane domains and motifs (F-Y-L-A-L-N-L-G-S-L) in,, and. High-throughput transcriptome analysis showed that the expression ofgene was up-regulated and thewas down-regulated by low concentration nitrate treatment for 72 h in roots, which caused more nitrate transferred from roots to shoots. On the contrary, the ethylene/jasmonic acid-NRT signaling modulecould promoteup-regulation and inhibit the expression ofby cadmium treatment. So that more nitrate transported from shoots to roots and improved the ability of plants to resist cadmium stress. This study is valuable for the research of biological functions ofandfamily genes inand the responses to nitrate-cadmium stress. Our results also provide references for the bioinformatic study ofandfamily genes in other plant species.
; bioinformatics;;; nitrate; cadmium
2018-07-18;
2018-10-08;
2018-11-02.
10.3724/SP.J.1006.2019.84099
张振华, E-mail: zhzh1468@163.com
E-mail: ghliang1119@163.com
本研究由国家重点研发计划项目(2017YFD0200100, 2017YFD0200103)和国家现代农业(油菜)产业技术体系建设专项资助。
This study was supported by the National Key R&D Program of China (2017YFD0200100, 2017YFD0200103) and the China Agriculture Research System.
URL: http://kns.cnki.net/kcms/detail/11.1809.s.20181031.1457.002.html
