通过分子标记辅助选择将耐储藏主效QTL qSS-9Kas转入宁粳4号提高其种子贮藏能力
2019-03-18姜一梅曹鹏辉张福鳞伍洪铭蔡梦颖刘世家田云录万建民
张 平 姜一梅 曹鹏辉 张福鳞 伍洪铭 蔡梦颖 刘世家 田云录 江 玲,* 万建民,2
通过分子标记辅助选择将耐储藏主效QTLqSS-9转入宁粳4号提高其种子贮藏能力
张 平1姜一梅1曹鹏辉1张福鳞1伍洪铭1蔡梦颖1刘世家1田云录1江 玲1,*万建民1,2
1南京农业大学作物遗传与种质创新国家重点实验室 / 农业部长江中下游粳稻生物学与遗传育种重点实验室 / 江苏省植物基因工程技术研究中心, 江苏南京 210095;2中国农业科学院作物科学研究所 / 农作物基因资源与基因改良国家重大科学工程, 北京 100081
利用置换系SL36作为位点上Kasalath等位基因的供体亲本, 各方面农艺性状较好的宁粳4号作为轮回亲本, 通过回交和自交, 并使用4个与紧密连锁的分子标记Y-10、Y-11、Y-14、Y-13进行基因型的检测筛选, 对宁粳4号进行耐贮性的遗传改良, 获得了既耐贮藏、大多农艺性状又接近于宁粳4号的较为稳定的优良新品系, 克服了宁粳4号耐贮藏性差的弱点。获得的新品系种子在人工老化和自然老化条件下均表现出发芽率明显提高、丙二醛含量显著降低、TTC染色效果更明显, 表明转入耐储藏主效QTL-9的宁粳4号新品系耐贮性显著提高。
水稻; 置换系; 分子标记辅助选择; 耐贮性; 遗传改良
水稻是我国最重要的粮食作物, 60%以上人口以稻米为主食。众所周知, 稻谷在一般贮藏条件下第2年就会产生陈化变质现象, 随着贮藏过程中温度和湿度的加大, 陈化过程也会加快。而具有良好耐贮藏特性的种子可以在同样条件下保存更长的时间, 并且在长时间贮藏后, 仍会表现出较高的萌发与出苗能力和优良的稻米品质, 可减少稻米陈化引起的损失[1]。种子耐贮性是一个受遗传因素、种子发育成熟度和贮藏期间环境条件影响的复杂数量性状, 与种子寿命和种子活力密切相关[2]。在同等贮藏条件下, 不同水稻品种的成熟种子在耐贮性上存在明显差异, 如贮藏1年的武育粳3号的发芽率只有14%, 而耐贮的W017仍保持87.3%的发芽能力, 表明遗传因素在种子耐贮性上有重要贡献。
目前, 利用自然或人工老化处理方法, 已检测到众多的水稻种子耐贮性相关QTL, 除了第8和第10染色体外, 在其他染色体上均有相关QTL分布, 在第9染色体上存在一个在不同遗传背景、不同生长环境下均稳定表达的主效QTL。Miura等[3]在Nipponbare/Kasalath//Nipponbare衍生的回交重组自交系(backcross inbred lines, BILs)群体中检测到主效QTL, 即。Ren等[4]用Zhenshan 97/Minghui 63衍生的重组自交系群体(recombinant inbred lines, RILs)检测到的与Miura等[3]检测到的位置一致, 可能为同一个QTL。Xue等[5]在Asominori/IR24衍生的RIL群体中检测到的与Miura等[3]报道的位点位置一致。Li等[6]利用Koshihikari/Kasalath//Koshihikari BIL群体在第9染色体检测到主效QTL, 来自Kasalath的对耐贮性起增效作用。Lin等[7]用Nipponbare/ Kasalath//Nipponbare BIL群体检测到3个QTL, 其中位于第9染色体上的对表型变异的贡献率最大, 该结果与Miura等[3]检测到的位点位置一致。这些结果说明,、和可能是同一个控制种子耐贮性的稳定的主效QTL。
本实验室Lin等[7]进一步通过自然老化和人工老化两种处理方法, 通过构建次级F2分离群体和一系列近等基因系, 对来自耐贮性极强的籼稻品种Kasalath等位基因(简写为)的遗传分析和精细定位, 将限定在146 kb之间, 获得了与耐贮性紧密连锁的分子标记Y10、Y11、Y14和Y13。本研究即是在此基础上, 利用这些紧密连锁的分子标记, 以宁粳4号为轮回亲本, 以耐贮性的置换系SL36为供体, 进行杂交和连续回交, 将转育宁粳4号, 创制携带耐贮的宁粳4号。
1 材料与方法
1.1 供试材料和主要农艺性状调查
“宁粳4号”是本实验室利用日本优质水稻品种“越光”和我国高产品种“镇稻99”杂交选育而成, 表现出高产稳产、综合抗性较强、适应性广、生育期适中、米质优良等特点[8], 但其耐贮性较弱。为提高宁粳4号的耐贮性, 以宁粳4号为轮回亲本, 以携带的日本晴(Nipponbare)为背景的染色体片段置换系SL36为供体(遗传背景见图1-A, 由日本农业生物资源研究所提供), 进行杂交和连续回交, 然后自交获得BC5F2代。
如图1-B, 2013年5月中旬将置换系SL36、宁粳4号种植于南京农业大学土桥水稻试验站(南京市江宁区淳化街道土桥社区), 将宁粳4号与置换系SL36杂交获得F1杂交种; 当年冬季南繁获得BC1F1代; 2014年5月中旬回交获得BC2F1代杂交种; 冬季南繁获得BC3F1代杂交种, 在2015年5月于土桥将BC3F1杂交收种后种植获得BC3F2自交种, 下文命名为Y6647。
根据4个与紧密连锁的分子标记Y-10、Y-11、Y-14、Y-13[9](表1)对收取119份BC3F2群体进行基因型的筛选, 将BC3F2在温度40℃、湿度80%的人工气候老化箱中处理25 d, 挑选种子活力显著高于宁粳4号的10份进行后代杂交繁育。2015年11月南繁选育获得BC4F1杂交种; 2016年5月于土桥获得BC4F2群体; 2016年11月南繁获得BC5F1杂交种。2017年5月于土桥收获BC5F2群体, 下文命名为Y3059, 均进一步通过MAS和耐储性鉴定, 选育出耐储性较稳定的品系, 成熟期调查株高、单株有效分蘖数、千粒质量等农艺性状。
1.2 种子耐贮性表型鉴定
1.2.1 自然老化处理 参考Li等[6], 略作改动。将同年正季在南京试验基地种植收获的亲本(置换系SL36、宁粳4号)和119份BC3F2代种子在室温<30℃、湿度40%~60%的环境中存放18个月。
1.2.2 人工老化处理 参照Zeng等[10], 略作修改。将同年正季在南京种植收获的置换系SL36和宁粳4号在智能型种子老化试验箱(江苏省南京研奥仪器设备有限公司, 型号为LH-300S)中40℃和80%湿度的条件下分别处理20、25和30 d。根据同年亲本预处理结果, 确定人工老化处理的时间。
1.2.3 发芽率测定 将人工老化处理后的种子发芽率作为种子耐贮性的间接评价指标[3]。随机选取50粒健康饱满经老化处理后的种子, 放入铺有两层滤纸的9 cm培养皿, 加8 mL水, 于30℃光照培养箱中培养7 d。以胚芽超过半粒种子长和胚根超过种子长度为发芽标准, 统计发芽率, 以发芽率的高低来判断种子的耐贮性强弱。设2个重复。
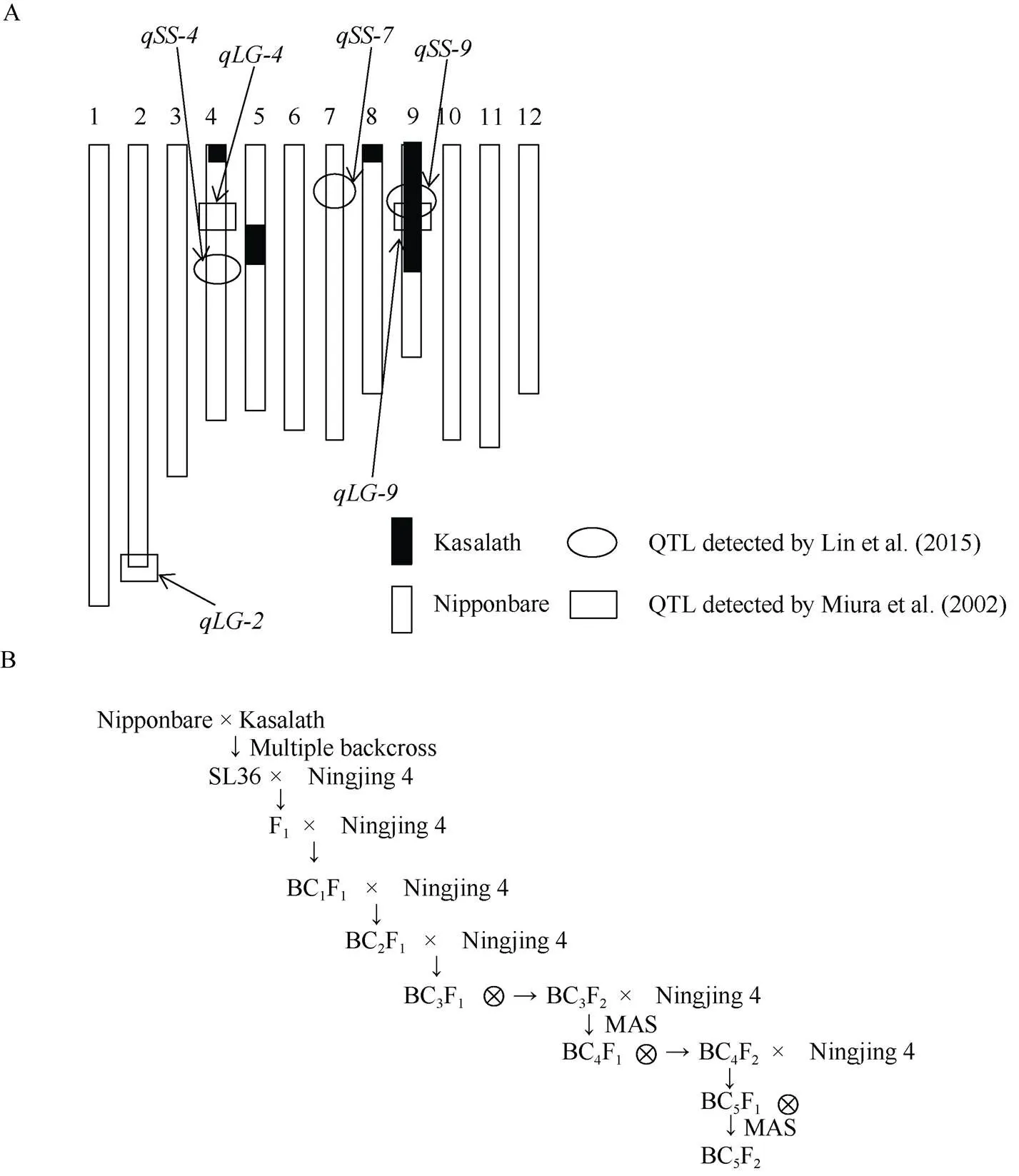
图1 染色体片段置换系SL36的基因型示意图(A)和回交育种流程图(B)
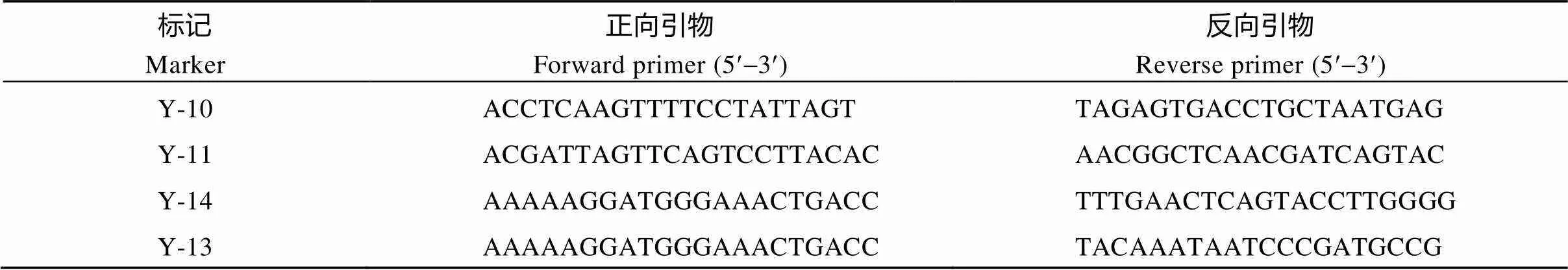
表1 本文中qSS-9的筛选标记
1.3 TTC (2,3,5-氯化三苯基四氮唑)法测定种子生活力
参考Lopez[11]的方法。取50粒脱皮的种子在30℃水中浸泡48 h, 然后在含有0.5% (w/v) TTC (pH7.0)的PBS (磷酸盐缓冲液)中黑暗染色2 h, 使用红色的强度作为活力的量度。
通过改进的测定法测量TTC还原含量[12]。TTC染色后, 将种子在滤纸上干燥并放入盛有10mL无水乙醇的15mL试管。在60℃水浴中浸泡6 h将种胚中红色物质提取出来, 在室温下以12,000 r min–1离心2 min, 测定上清液在分光光度计490 nm处OD值。在所有步骤中, 用未经TTC染色的种子作空白对照。设3个重复。
1.4 生理生化指标测定
1.4.1 直链淀粉含量测定 按照国家标准GB-7648-87法经改进简化, 称取过0.15 mm筛的待测精米粉样品和标准样品0.1000 g放入100 mL容量瓶, 加1 mL 95%乙醇和9 mL 1 mol L–1NaOH溶液, 沸水浴10 min后, 加蒸馏水定容至100 mL。吸取5 mL 样品溶液至100 mL容量瓶中, 加1 mL 1 mol L–1乙酸溶液和1.5 mL碘液显色并定容, 静止20 min后在620 nm 波长下测定吸光值, 计算宁粳4号和置换系后代的直链淀粉含量。设3个重复。
1.4.2 MDA (丙二醛)含量测定 通过以下步骤从过完100目筛的糙米粉中提取MDA。取6粒左右健康饱满的稻谷手工去壳后, 收集胚及糊粉层的混合物。在2.0 mL离心管中, 加入1 mL 0.05 mol L–1NaH2PO4-Na2HPO4缓冲液(pH 7.4), 在混合仪(Retsch MM301, 德国)上以1/30 s的周期振荡2 min后, 提取0.1 g糙米粉。测定采用MDA测定试剂盒(南京建成生物工程研究所), 以nmol mg–1表示。
2 结果与分析
2.1 携带qSS-9Kas的染色体片段置换系具有强的耐贮性
室温下贮藏18个月的宁粳4号的种子发芽率仅为26.70%±0.03%, 而置换系SL36和F1杂交种(SL36×宁粳4号)的发芽率分别为91.10% ±0.05%和80.90% ±0.08%, 均与宁粳4号存在极显著差异(< 0.01)(图2-A, B), 表明位点上的Kasalath等位基因表现为强的耐贮性, 且为显性效应。TTC染色结果表明, 大部分宁粳4号种子的胚部均未染上红色, 部分种子胚部虽已着色, 但着色程度较浅, 表明宁粳4号种子已基本丧失生活力。而置换系SL36和F1种子的种胚大多染上红色且着色程度深, 表现出较强的种子生活力(图2-C)。该结果与种子发芽测定结果一致, 说明两种方法均能检测种子的耐贮性。
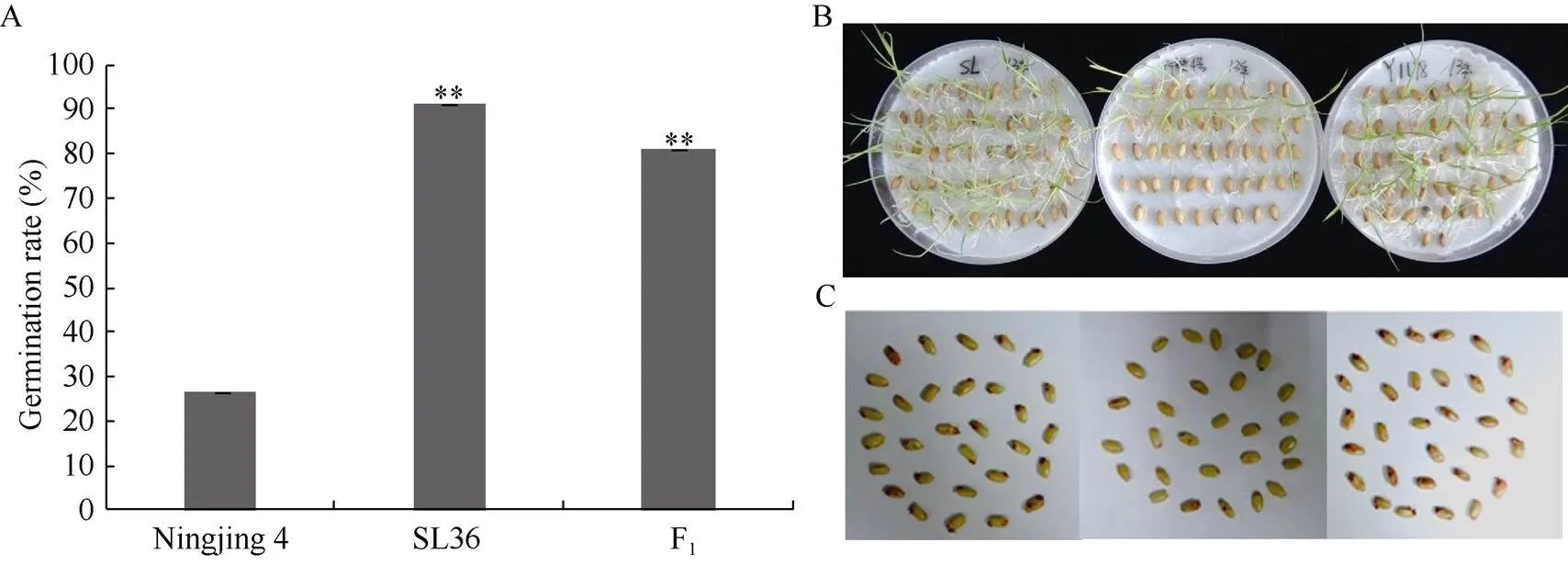
图2 亲本及F1种子在自然环境下贮藏18个月后的种子发芽率和TTC法测定的种子生活力
A: 亲本及F1种子发芽率测定; B、C: 亲本及F1发芽表现和TTC法种子生活力表现, 从左到右依次是SL36、宁粳4号和F1。**表示与宁粳4号的发芽率相比, 在0.01水平上存在极显著差异。
A: Germination rates of parents and F1; B,C: Seed germination and tetrazolium assay of parents SL36, Ningjing 4 and F1(from left to right). ** means significant at the 0.01 probability level compared with the germination rate of Ningjing 4.
2.2 利用MAS方法, 将控制耐贮性的主效QTL qSS-9Kas导入宁粳4号
利用与紧密连锁的分子标记Y-10、Y-11、Y-14、Y-13对各亲本进行基因型分析。分子标记电泳图谱显示宁粳4号与不耐贮的粳稻品种Nipponbare一样, 在位点上不同于Kasalath的基因型, 即不携带qSS-9, 而置换系SL36则含有该QTL的Kasalath等位基因qSS-9(图3-A)。在收取的200份置换系后代BC5F2的基因型鉴定和目的单株选育(图3-B)中, 带型和SL36一致的有194个。
共选择212个均匀分布于12条染色体的多态性标记对2017年正季BC5F2代单株进行置换系背景鉴定, 发现BC5F2中宁粳4号遗传背景达到约97.6%, 表明经过高代回交,qSS-9已被成功转育到宁粳4号中, 遗传背景已较纯合。
2.3 携带qSS-9Kas的宁粳4号改良系老化后丙二醛含量降低
种子中MDA的含量是衡量脂质过氧化的主要指标, 随着稻米贮藏时间的延长, 种子活性降低, MDA的含量不断增加。自然老化处理一年后测定种子发现, SL36、Y6647中MDA的含量(1.60~1.85 nmol mg–1)明显低于对照宁粳4号(5.65 nmol mg–1)(图4-A), 说明与对照相比, 在常温条件储存下,携带qSS-9的置换系和改良系种子的劣变及衰老过程延缓, 脂质降解产生的MDA较少。
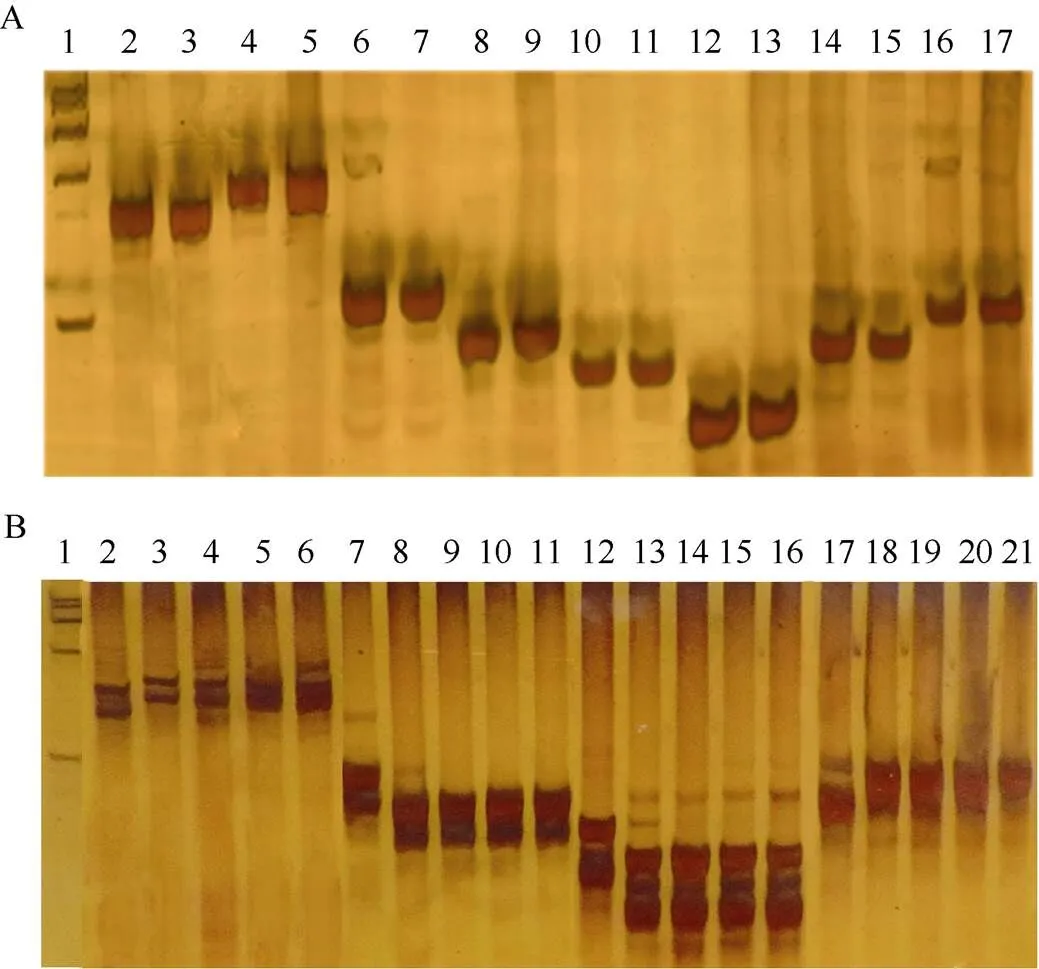
图3 各亲本和BC5F2在与qSS-9紧密连锁的分子标记Y-10、Y-11、Y-14、Y-13下的基因型鉴定
A: 利用与紧密连锁的分子标记Y-10、Y-11、Y-14和Y-13对各亲本的基因型进行鉴定; 其中, 1:DNAmarker; 泳道2~5、6~9、10~13和14~17分别代表各品系在Y-10、Y-11、Y-14和Y-13的基因型; 泳道2、6、10和14为Nipponbare; 泳道3、7、11和15为宁粳4号; 泳道4、8、12和16为置换系SL36; 泳道5、9、13和17为Kasalath。B: 利用与紧密连锁的分子标记Y-10、Y-11、Y-14和Y-13对宁粳4号、SL36及BC2F1间的基因型进行鉴定: 其中1:DNA marker; 泳道2~6分别代表宁粳4号、SL36及BC2F1在Y-10下的基因型; 7~11分别代表宁粳4号、SL36及BC5F2在Y-11下的基因型; 12~16分别代表宁粳4号、SL36及BC2F1在Y-14下的基因型; 17~21分别代表宁粳4号、SL36及BC2F1在Y-13下的基因型。
A: Genotype identification of parents by molecular markers Y11 and Y14 close linkage to; 1:DNA marker; 2-5, 6-9, 10-13, and 14-17 indicate genotype of markers Y-10, Y-11, Y-14, and Y-13 respectively; 2, 6, 10, and 14 indicate genotype of Nipponbare; 3, 7, 11, and 15 indicate genotype of Ningjing 4; 4, 8, 12, and 16 indicate genotype of SL36; 5, 9, 13, and 17 indicate genotype of Kasalath, respectively. B: Genotype identification of Ningjing 4, SL36 and BC5F2by molecular markers Y11 and Y14 close linked to; 1:DNA marker; 2-6 : Genotype of Ningjing 4, SL36 and BC5F2under marker Y-10; 7-11: Genotype of Ningjing 4, SL36 and BC5F2under marker Y-11; 12-16: Genotype of Ningjing 4, SL36 and BC5F2under marker Y-14; 17-21: Genotype of Ningjing 4, SL36 and BC5F2under marker Y-13, respectively.
2.4 携带qSS-9Kas的宁粳4号改良系耐储性显著增强
自然条件储存一年后, 宁粳4号的发芽率为78.0%±5.7%, 置换系SL36的发芽率为93.0%±1.4%, 置换系后代BC3F2发芽率为73.0%±12.7%, 但是继续储存一年后, 宁粳4号的发芽率降到20%以下,而置换系SL36的发芽率为77.0%±4.2%, BC3F2的发芽率仍保持在57.7%±8.4%, 活力相比宁粳4号显著提高(图4-B)。
2016年海南收获的宁粳4号、置换系SL36和BC5F1代的破除休眠后种子在温度40℃和湿度80%条件下老化35 d后, 宁粳4号的发芽率已降至69% ±1%, 而置换系SL36和BC5F1的发芽率仍分别保持在95%±2.5%和96.5%±3.0%, 表明qSS-9导入宁粳4号后, 显著提高了其抗人工老化的能力(图4-C)。
将2017年正季收获的宁粳4号、置换系SL36和BC5F2代的种子破除休眠后, 人工老化处理30 d, 宁粳4号的发芽率为55.6%±1.7%, 而置换系SL36的发芽率为85.6%±2.6%, 两者之间存在极显著差异(0.01)。从194份含有目的片段的单株中挑选出发芽率较宁粳4号显著提高的186份, 其中农艺性状与宁粳4号基本一致的3份种子发芽率分别为78%± 4%、91%±1%、76%±2%(图4-D, E, F, G, H), 显著高于宁粳4号, 表明qSS-9导入宁粳4号, 显著提高其种子耐贮性。
2.5 携带qSS-9Kas的宁粳4号改良系种子活力增强
TTC在种子脱氢酶的作用下, 可还原产生TTCH (三苯甲簪), 以还原力的高低来判定种子的活力。TTC被还原越多, 产生红色的TTCH就越多, 种子的活力越高, 用无水乙醇将不溶于水的TTCH从种胚中提取出来, 测量提取液在TTCH最大吸收波长490 nm处的吸光值, 以检测种子的活力。取人工老化30 d的宁粳4号及携带qSS-9的宁粳4号改良家系种子, 进行TTC染色, 如图5-A所示, 改良系种子被染色的部分更多更深, 对TTCH含量测定发现, 改良系种子中TTCH的含量是对照宁粳4号的1.5~2.0倍(图5-B)。
2.6 携带qSS-9Kas的宁粳4号改良家系的农艺性状表现
由于稻米直链淀粉含量是决定品质的最重要性状之一, 其含量与米饭的黏性、柔软性、光泽和食味品质密切相关。为此, 我们测定了宁粳4号和Y3059-1-7、Y3059-1-8、Y3059-1-9老化处理前后直链淀粉的含量随种子老化程度加深, 伴随着种子发芽率的降低, 种子中直链淀粉的含量也有所降低(图5-C), 但具体的变化机制仍需进一步探究。

图4 亲本和后代种子在不同老化处理下的发芽率和丙二醛的含量
A: 宁粳4号、SL36、BC3F2(Y6647)在自然老化一年MDA含量; B: 宁粳4号、SL36、BC3F2在自然老化一年和两年的发芽率; C: 亲本和BC5F1种子在人工老化处理35 d后的发芽率; D: 宁粳4号、Y3059-1-7、Y3059-1-8、Y3059-1-9的发芽率; E~H: 老化处理30 d后的发芽情况。E、F、G、H分别代表宁粳4号、Y3059-1-7、Y3059-1-8、Y3059-1-9。*表示在= 0.05水平上差异显著; **表示在= 0.01水平上差异极显著。
A: Contents of MDA in seeds of Ningjing 4, SL36, and BC3F2after natural aging for one year; B: Germination rate of Ningjing 4, SL36 and BC3F2after natural aging forone year and two years; C: Germination rates of parents and BC5F1after artificial aging for 35 days; D: Seed germination of BC5F2and Ningjing 4 after accelerated aging for 30 d and storage at 40°C and 80% RH; E-H: Seed viability of the control after aging treatment; E, F, G, and H represent Ningjing 4, Y3059-1-7, Y3059-1-8, Y3059-1-9, respectively. Significant differences relative to the control were assessed by-tests. * and **, significant at= 0.05 and= 0.01, respectively. Values are means ± SD (= 2).
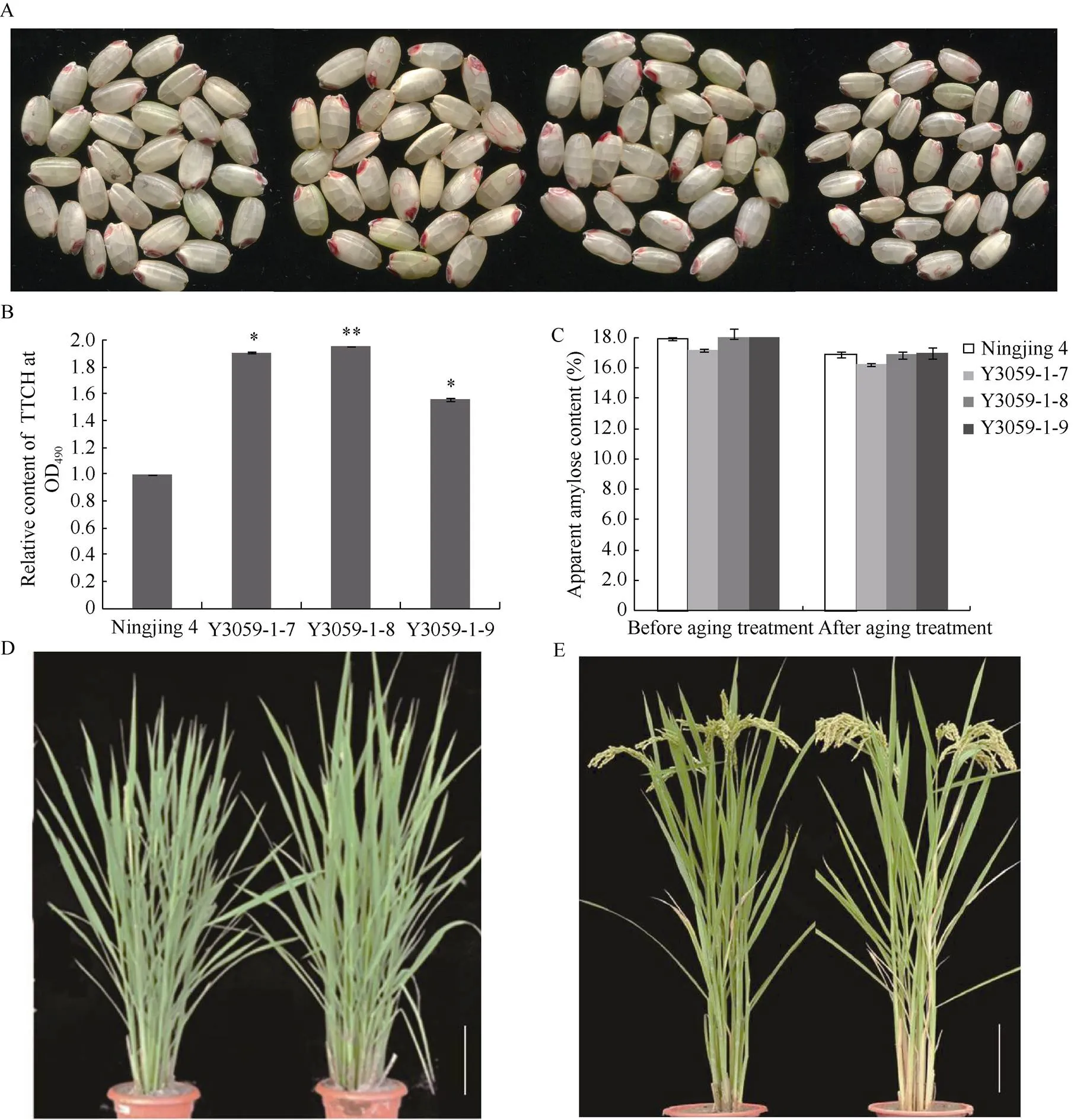
图5 宁粳4号和Y3059的TTC试验、直链淀粉含量和株型的比较
A: 老化处理30 d后宁粳4号和Y3059的TTC染色情况, 从左到右分别代表宁粳4号、Y3059-1-7、Y3059-1-8、Y3059-1-9; B: Y3059及宁粳4号种子中TTCH的含量; C: 宁粳4号和Y3059老化处理前后表观淀粉含量的变化; D: 抽穗时期宁粳4号(左)和BC5F2(右)株型; E: 抽穗35 d后宁粳4号(左)和BC5F2(右)株型。*表示在= 0.05水平上差异显著;**表示在= 0.01水平上差异极显著。
A: Tetrazolium assay of Ningjing 4 and Y3059; From left to right represent Ningjing 4, Y3059-1-7, Y3059-1-8, Y3059-1-9, respectively; B: The content of TTCH of Y3059and Ningjing 4; C: The change of apparent amylose content in Ningjing 4 and Y3059;D: Plant type of Ningjing 4 and BC5F2at heading;E: Plant type of Ningjing 4 and BC5F2at 35 days after heading; Significant differences relative to the control were assessed by-tests. * and **, significant at= 0.05 and= 0.01, respectively. Values are means ± SD (= 2).
对改良家系进行农艺性状考察, 如图5-D, E所示, 改良系和宁粳4号的抽穗期、结实率、千粒重、单株产量都没有显著差异(表2)。表明在提高宁粳4号耐贮性的同时并没有影响其主要的农艺性状。

表2 宁粳4号和Y3059农艺性状的比较
*表示在= 0.05水平上差异显著;**表示在= 0.01水平上差异显著。
*and**, significant at= 0.05 and= 0.01, respectively. HD: heading date; PH: plant height;NT: number of tillers;SSR: seed setting rate; TGW: 1000-grain weight; YPP: yield per plant.
3 讨论
本实验室Li等[6]利用Koshihikari/Kasalath// Koshihikari BIL群体在南京、连云港、金湖3个不同地方种植并进行种子耐贮性相关QTL分析, 在第9染色体标记R10783S和R1751区间检测到qSS-9, 并且利用CSSL群体验证了qSS-9的真实性。该位点与Miura等[3]检测到的位置一致, 在此基础上, 林秋云[9]利用以Nipponabre为背景, Kasalath为置换片段的染色体片段置换系SL36与Nipponabre回交, 构建次级F2分离群体, 对qSS-9进行遗传分析, 同时进行F3后代验证和构建的近等基因系进行验证, 获得了与qSS-9紧密连锁的Indel标记Y11和Y14, 在Nipponbare基因组上的物理距离约为147 kb。同时用以Koshihikari为背景, Kasalath为置换片段的染色体片段置换系SL226与Koshihikari回交, 构建次级F2分离群体, 对qSS-9进行精细定位, 将qSS-9定位在Indel标记Y7和Y13之间, 在Nipponbare基因组上的物理距离约为478 kb。两种qSS-9的精细定位区间重叠。经过多年多点的检测, 发现qSS-9是真实存在并稳定表达的主效QTL。本研究在前期定位基础上, 利用4个Indel标记Y10、Y11、Y14和Y13, 对本实验室选育的主栽粳稻品种宁粳4号与耐贮性的染色体片段置换系SL36的回交和自交后代单株, 进行分子标记的选择, 再经过老化后种子活力检测及目标性状的选育, 已将qSS-9转育到宁粳4号中, 获得耐贮性的宁粳4号。
影响种子耐贮性的因素很多, 如种子自身的遗传因素、种子灌浆成熟时的气候条件、收获干燥条件、种子贮藏时的外界环境等。本研究发现在同一型号的同一台老化箱中, 每次老化的时间均有一定的波动, 如南繁收获的种子, 可能由于海南热带季风气候, 种子灌浆时期温度较高, 收获时较为干燥, 种子的生长状况良好, 耐贮性稍强, 人工老化时间也会相应增加, 说明水稻自身的状况对种子耐贮性有一定的影响。本研究中虽然不同年份不同批次的老化试验结果略有差异, 但总体趋势是一致的, 转育了qSS-9的宁粳4号, 其耐贮性均明显高于背景亲本宁粳4号。同时, 本文使用了两种不同的老化方法, 在短时间内用自然老化方法较难评价种子的耐贮性[13]; 人工老化是在人为控制的环境条件下加快种子老化速率, 高温高湿是目前最常用的人工老化方法[14]。通过人工老化处理后种子在短时间内发芽率下降明显, 因此可以快速评价种子的耐贮性, 弥补自然老化所需较长时间的缺点。Rajjou等[15]认为人工加速老化可以模拟自然老化从而提高鉴定种子耐贮性的效率。许惠滨等[16]认为短时间的高温高湿加速老化条件不能完全替代自然老化。从本文的研究结果看, 人工老化虽然与自然老化结果有一定的差异, 但两种方法均可验证, 在通过不断回交转育的后代中, 种子的耐贮性的确得到提高, 因此, 人工老化同样可适用于分子标记辅助育种。我们已成功地将耐储藏主效QTLqSS-9引入到主栽品种宁粳4号中。
生产上, 人们一般通过改善贮藏条件, 如修建气调库或低温库、选择合适的包装材料和微波处理种子等[17], 虽然可以延缓稻米贮藏期间的陈化, 但需要消耗大量的人力和财力。从遗传角度改良种子本身的耐贮性, 能更加经济、有效地解决种子在贮藏过程中的陈化变质问题。因此, 水稻种子耐贮性的分子育种已经成为水稻育种的重要目标之一。本研究采用人工老化为主, 自然老化为辅, 结合MAS方法, 将来源于Kasalath的控制种子耐贮性的主效QTLqSS-9转移到宁粳4号中, 从种子自身的遗传基因来提高种子的耐贮性, 经过多年多种方法验证, 表明该手段能稳定有效提高其种子的耐贮性。如自然条件下贮藏2年的种子, 宁粳4号的发芽率已低于20%, 而改良家系Y6647的发芽率仍保持在57.7% ±8.4%。为耐贮性分子育种奠定了基础。
4 结论
利用分子标记辅助选择技术将来源于Kasalath的控制种子耐贮性的主效QTLqSS-9转育到主栽品种宁粳4号中, 获得的宁粳4号改良系表现出更高的耐贮性, 同时大多数农艺性状和宁粳4号无显著差异。
[1] Yamauchi M, Winn T. Rice seed vigor and seedling establishment in anaerobic soil., 1996, 36: 680–686.
[2] 沈圣泉, 庄杰云, 王淑珍, 杨国花, 夏英武. 水稻种子耐贮藏性QTL主效应和上位性效应分析. 分子植物育种, 2005, 3: 323–328. Shen S Q, Zhuang J Y, Wang S Z, Yang G H, Xia Y W. Analysis of QTLs with main and epistatic effects of the seed storability in rice., 2005, 3: 323–328 (in Chinese with English abstract).
[3] Miura K, Lin S Y, Yano M, Nagamine T. Mapping quantitative trait loci controlling seed longevity in rice (L.)., 2002, 104: 981–986.
[4] 任淦, 彭敏, 唐为江, 徐才国, 邢永忠. 水稻种子衰老相关基因定位. 作物学报, 2005, 31: 183–187. Ren G, Peng M, Tang W J, Xu C G, Xing Y Z. QTL associated with seed aging in rice., 2005, 31: 183–187 (in Chinese with English abstract).
[5] Xue Y, Zhang S Q, Yao Q H, Peng R H, Xiong A S, Li X, Zhu W M, Zhu Y Y, Zha D S. Identification of quantitative trait loci for seed storability in rice (L.)., 2008, 164: 739–744.
[6] Li L F, Lin Q Y, Liu S J, Liu X, Wang W Y, Ngo T H, Liu F, Zhao Z G, Jiang Ling, Wan, J M. Identification of quantitative trait loci for seed storability in rice (L.)., 2012, 131: 739–743.
[7] Lin Q Y, Jiang Y M, Sun A L, Cao P H, Li L F, Liu X, Tian Y L, He J, Liu S J, Chen L M, Jiang L. Fine mapping of, a major and stable quantitative trait locus, for seed storability in rice (L.)., 2015, 134: 293–299.
[8] 王学明, 吴爱国. 中熟中粳宁粳4号种植表现及高产栽培技术. 耕作与栽培, 2010, (5): 58. Wang X M, Wu A G, Planting performance and high-yielding cultural techniques of medium-maturity Ningjing 4., 2010, (5): 58 (in Chinese).
[9] 林秋云. 水稻种子耐贮性相关基因的定位. 南京农业大学博士学位论文, 江苏南京, 2012. Lin Q Y. Gene Location for Seed Storability in Rice (L.). PhD Dissertation of Nanjing Agricultural University, Nanjing, Jiangsu, China, 2012 (in Chinese with English abstract).
[10] Zeng D L, Guo L B, Xu Y B, Yasukumi K, Zhu L H, Qian Q. QTL analysis of seed storability in rice., 2006, 125: 57–60.
[11] Lopez D E L, David N M, Victor M H, Toorop P E, Iannetta P P M. A spectrophotometric assay for robust viability testing of seed batches using 2,3,5-triphenyl tetrazolium chloride: usingL. as a model., 2017, 8: 747.
[12] Zhao P F, Zhu Y H, Wang W. Evaluation and improvement of spectrophotometric assays of TTC reduction: maize () embryo as an example., 2010, 32: 815–819.
[13] Sasaki K, Fukuta Y, Sato T. Mapping of quantitative trait loci controlling seed longevity of rice (L.) after various periods of seed storage., 2005, 124: 361–366.
[14] Ngo T H, Lin Q Y, Liu L L, Liu X, Liu S J, Wang W Y, Li L F, He N Q, Liu Z, Jiang L, Wan J M. Mapping QTLs related to rice seed storability under natural and artificial aging storage conditions., 2015, 203: 673–681.
[15] Rajjou L, Lovigny Y, Groot S P C, Belghaz, M, Job C, Job D. Proteome-wide characterization of seed aging in Arabidopsis: a comparison between artificial and natural aging protocols., 2008, 148: 620–641.
[16] 许惠滨, 魏毅东, 连玲, 朱永生, 谢华安, 王宗华, 张建福. 水稻种子人工老化与自然老化的分析比较. 分子植物育种, 2013, 11: 552–556. Xu H B, Wei Y D, Lian L, Zhu Y S, Xie H A, Wang Z H, Zhang J F. Comparative analysis of artificial aging and natural aging with rice seeds., 2013, 11: 552–556 (in Chinese with English abstract).
[17] Sharp R N, Timme L K. Effects of storage time, storage- temperature, and packaging method on shelf life of brown rice., 1986, 63: 247–251.
Introducinginto Ningjing 4 by molecular marker-assisted selection to improve its seed storage ability
ZHANG Ping1, JIANG Yi-Mei1, CAO Peng-Hui1, ZHANG Fu-Lin1, WU Hong-Ming1, CAI Meng-Ying1, LIU Shi-Jia1, TIAN Yun-Lu1, JIANG Ling1,*, and WAN Jian-Min1,2
1State Key Laboratory of Crop Genetics and Germplasm Enhancement / Key Laboratory of Biology, Genetics and Breeding of Japonica Rice in Mid-lower Yangtze River, Ministry of Agriculture / Research Center of Jiangsu Plant Gene Engineering, Nanjing Agricultural University, Nanjing 210095, Jiangsu, China;2National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China
In this paper, the chromosome segment substitution line SL36 was used as the donor parent with the Kasalath allele at thelocus. Ningjing 4, a commercial cultivar with ideal agronomic traits, was used as a recurrent parent, through continuous self-pollination and backcrossing for four consecutive years. Four molecular markers Y-10, Y-11, Y-14, Y-13 closely linked towere used to screen genotypes, and molecular marker-assisted selection (MAS) was used for seed storage ability breeding for Ningjing 4. Through genetic improvement, we obtained inheritable lines with high seed storage ability. Most agronomic traits of the lines were nearly the same as those of Ningjing 4. These lines showed significantly higher germination rate, lower malondialdehyde content and more obvious TTC staining effects under artificial aging and natural aging conditions compared with Ningjing 4, indicating that the new lines withhave high seed storage ability.
rice; chromosome segment substitution line; molecular marker-assisted selection; seed storage ability; genetic improvement
2018-07-06;
2018-12-24;
2019-01-03.
10.3724/SP.J.1006.2019.82035
江玲, E-mail: jiangling@njau.edu.cn
E-mail: 2016101096@njau.edu.cn
本研究由国家转基因生物新品种培育重大专项(2016ZX08001006), 江苏省重点研发项目(BE2018388, BE2017368)和江苏省现代作物生产协同创新中心项目资助。
This study was supported by the National Major Project for Developing New GM Crops (2016ZX08001006), the Key Science and Technology Project of Jiangsu Province (BE2018388, BE2017368), and Jiangsu Collaborative Innovation Center for Modern Crop Production.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20181229.1351.002.html
