Jmjd3和Ezh2酶活性抑制剂对人间充质干细胞成脂分化的影响及机制研究
2019-03-13武晓慧李蓉黄振鹏杨帆陈佳琪
武晓慧 李蓉 黄振鹏 杨帆 陈佳琪


摘要:目的 研究EZH2和JMJD3对人脂肪间充质干细胞成脂分化的影响及可能机制。方法 体外培养人脂肪来源间充质干细胞并进行诱导分化。分化过程中分别加入EZH2酶活性抑制剂GSK126(6 μM)和JMJD3酶活性抑制剂GSKJ4(20 μM),对照组加入DMSO。采用Realtime PCR方法检测脂肪细胞分化基因PPARγ、FABP4和ADIPOQ;棕色化标志基因UCP1、PRDM16和CIDEA,以及米色脂肪独有的标志基因CD137、TMEM26和TBX1,并计算各组与对照组的相对表达量。采用Western Blot法检测H3K27me3、UCP1、JAK2、STAT3和pSTST3的蛋白表达量。结果 GSK126组的脂滴形成略减少,GSKJ4组的脂滴形成明显减少。GSK126组的H3K27me3含量减少,GSKJ4组的H3K27me3含量增加;GSK126组的UCP1、PRDM16、TMEM26和JAK2基因表达均明显增加(P<0.05);GSKJ4组的PPARγ、UCP1和JAK2的基因表达量均明显降低(P<0.05);GSK126组的UCP1蛋白表达增加而GSKJ4组UCP1蛋白表达减少。GSK126组的JAK2和pSTST3均增加,GSKJ4组的JAK2和pSTST3蛋白表达均减少。结论 EZH2酶活性抑制剂GSK126促进人MSC向棕色或米色脂肪细胞分化。JMJD3酶活性抑制剂GSKJ4抑制人MSC向棕色或米色脂肪细胞分化;EZH2和JMJD3的酶活性影响H3K27的三甲基化修饰,可能通过JAK2-STAT信号通路,发挥调控人脂肪细胞分化的作用。
关键词:间充质干细胞;分化;EZH2;JMJD3;UCP1;JAK2c
中图分类号:R364.2 文献标识码:A DOI:10.3969/j.issn.1006-1959.2019.02.020
文章编号:1006-1959(2019)02-0063-06
肥胖是当今世界最常见的慢性非传染性疾病之一[1],我国的肥胖人数已达世界之最,2010年的调查结果显示,有41.7%以上的人群处于超重或者肥胖状态[2]。肥胖可以引发多种疾病,如胰岛素抵抗、2型糖尿病、心脑血管疾病、骨关节炎、部分癌症等[3,4]。多项研究表明,啮齿动物和人体内的棕色和米色脂肪细胞可以通过产热消耗能量,对抗肥胖和胰岛素抵抗,因而这两类脂肪细胞的分化成为肥胖治疗领域的研究热点[5]。
目前对于脂肪细胞分化的研究已由针对单个基因功能的研究转为基因表达调控的研究,如表观遗传学调控机制。组蛋白修饰是表观遗传调控基因表达的方式之一,研究发现,组蛋白H3K27me3去甲基化酶jmjd3可以促进小鼠棕色脂肪细胞的分化[6],而H3K27me3甲基转移酶Ezh2可以在体外促进小鼠白色和棕色脂肪细胞的分化[7],但在人体细胞尚无相关报道,其调控机制亦尚不清楚。研究发现,JAK2可通过JAK2/STAT途径调控脂肪细胞分化[8]。本实验拟以人脂肪来源间充质干细胞(mesenchymal stem cells,MSC)为研究对象,在诱导分化过程中使用EZH2酶活性抑制剂GSK126或JMJD3酶活性抑制剂GSKJ4处理细胞,观察细胞成脂分化情况,并检测JAK2及STAT3的表达,揭示Jmjd3和Ezh2对人间充质干细胞向脂肪细胞分化过程的影响及可能机制。
1材料与方法
1.1主要试剂 人脂肪来源间充质干细胞(中科院上海细胞库);间充质干细胞培养液(MSCM,7501,美国Sciencell);DMEM培養液(美国Sigma);胎牛血清(四季青);人用生物合成短效胰岛素(诺和灵R,诺和诺德);3-异丁基-1-甲基黄嘌呤(IBMX)(美国Sigma);地塞米松(美国Sigma);三碘甲腺原氨酸(T3,上海麦克林);罗格列酮(麦克林)。用于western blot的一抗:兔Ezh2 抗体(1∶1000;CST);兔Jmjd3抗体(1∶1000,Abcam);兔H3K27me3抗体(1∶1000;Abcam);兔JAK2抗体(1∶1000;Abcam);兔STAT3抗体(1∶1000;Abcam);兔磷酸化STAT3抗体(1∶1000;Abcam);小鼠β-Tubulin(1∶1000;天津三箭)。二抗:HRP-羊抗兔IgG、HRP-羊抗鼠IgG(1∶5000;北京中杉金桥)。全蛋白提取试剂盒、BCA蛋白浓度测定试剂盒(碧云天)。用于Realtime PCR的RNA提取试剂盒、反转录试剂盒、Syber Green Master Mix(日本Takara)。小分子抑制剂GSK126、GSKJ4(美国MCE);油红0粉末(Sigma)。
1.2细胞的分化方法 实验设3个组,分别为对照组,GSK126组和GSKJ4组,将细胞种于12孔板内,每个组设3个复孔。细胞扩增达100%汇合时出现接触抑制,细胞停止分裂进入分化阶段(G0期),设为分化第2天。接触抑制2 d后,开始分化,为分化第0天。每孔加入分化液,并且按照设计分别加入GSK126、GSKJ4或DMSO;分化液培养2 d后换用维持培养液培养,每隔2 d换液1次,换液后根据分组加入GSK126或GSKJ4,对照组加入等体积DMSO。分化第8天时,对照组已经全部出现脂滴,此时结束分化。
1.3分化液的配制
1.3.1分化液的成份 经摸索实验条件,MSC的扩增培养和传代使用MSCM培养液,分化时换用DMEM培养液分化更好。分化液的配制:在50 ml DMEM完全培养液中加入胰岛素、IBMX、地塞米松、罗格列酮和T3;维持液的配制:在50 ml DMEM完全培养液中加入胰岛素、罗格列酮和T3。所有药品加入后0.22 μm濾器过滤于新的灭菌50 ml离心管,4 ℃保存。
1.3.2分化用药品的配制 短效胰岛素:50 ml培养液中加入60 μl。IBMX的工作浓度为0.5 mM,称取IBMX粉剂 222.2 mg,溶于2 ml DMSO,配成1000×母液,用时50 ml培养液中加入IBMX母液50 μl。地塞米松工作浓度为5 μm,称取3.92 mg地塞米松,加入1 ml DMSO,配成10000×母液,用时在50 ml培养液中加入地塞米松母液5 μl。罗格列酮的工作浓度为1 μm,称取0.357 mg罗格列酮,加入1 ml DMSO,配成1000×母液,用时在50 ml培养液中加入罗格列酮母液5 μl;T3工作浓度为1 nm,称取0.0651 mg T3,加入10 ml DMSO,配成10000×母液。用时在50 ml培养液中加入T3母液 5 μl。
1.3.3 GSK126及GSKJ4储存液的配制及使用 GSK126的分子量为526.6 g/mol。称取GSK126粉剂1.05 mg,加入DMSO 1 ml,0.22 μm微孔滤膜过滤,工作浓度为6 μm,用时每1 ml培养液加3 μl母液。GSK J4的分子量为417.5 g/mol。称取GSKJ4粉剂2.8 mg,加入DMSO 1 ml,充分溶解后,0.22 μm微孔滤膜过滤,工作浓度为20 μm,用时每1 ml培养液加3 μl母液。因培养液中DMSO>1‰,为排除影响,对照组每1 ml培养液加3 μl DMSO。
1.4细胞的油红染色 配制油红O染液:称取油红O 粉末0.5 g,溶于100 ml异丙醇,滤纸过滤,避光4℃保存。使用时配制工作液,用时现配:取6 ml储存液加ddH2O 4 ml,混匀后静置10 min,用滤纸过滤。染色时,细胞用预冷的4%甲醛固定10 min,PBS缓冲液洗1次。油红O工作液避光染色30 min;60%异丙醇浸洗1~2次,除去多余的油红O染液。PBS浸洗2次,在吸水纸上拍干,滴加50%甘油封片剂(甘油:PBS缓冲液体积比=1∶1),在倒置显微镜下观察并照相。
1.5 Realtime PCR检测mRNA的表达 Trizol法提取总RNA。将总RNA反转录为cDNA后进行Realtime PCR反应。实验所需引物由大连宝生公司设计并合成,序列信息见表1。以小鼠β-actin为内参。反应体系为20 μl。反应程序为:95 ℃预变性30 s,随后42个循环:95 ℃变性5 s,56 ℃退火及延伸30 s。每个基因设3个复孔,采用2-△△Ct法计算基因的相对表达量。实验重复3次,采用Graphpad prism 5软件绘制成柱状图。
1.6 Western blot 法检测蛋白表达 倒掉培养液,向培养皿中加入RIPA裂解液(已经加入PMSF),冰上放置裂解30 min;细胞刮刀刮下细胞,并吸出液体至1.5 ml离心管,13000 rpm 4 ℃离心5 min。将离心后的上清转移至新的1.5 ml的离心管中,上清液取5 μl采用BCA法做蛋白浓度测定,其余按照体积加入5×loading Buffer上样缓冲液(内含β-巯基乙醇),混匀,95 ℃金属浴10 min,-20 ℃保存备用。根据待测蛋白的分子量配制不同浓度的SDS-Page分离凝胶,蛋白上样量为20~50 μg,电泳完成后100 V 1 h湿转至PVDF膜,5%脱脂奶粉封闭后4 ℃孵育一抗过夜,相应的二抗室温孵育1 h,ECL发光液发光。
1.7统计学方法 数据用SPSS 20.0 统计软件处理,所有数据采用(x±s)表示,组间比较采用独立样本均数t检验,以P<0.05为统计学意义显著,P<0.01为统计学意义极显著。
2结果
2.1 GSK126对人MSC成脂分化影响较小,GSKJ4明显减少MSC的成脂分化 实验设置3个组,分别是GSK126组,GSKJ4组和溶剂对照组(DMSO)。我们在培养液中根据文献提供的浓度,在分化诱导和维持期持续加入GSK126和GSKJ4。分化8 d后进行拍照,发现加入抑制剂后,细胞分化仍可进行,但脂滴形成有差异。GSK126组脂滴形成略减少,GSKJ4组脂滴形成明显减少,见图1(上)。经油红染色证实,对照组脂滴较多,GSK126组的脂滴形成略减少,GSKJ4组脂滴形成明显减少,见图1(下)。
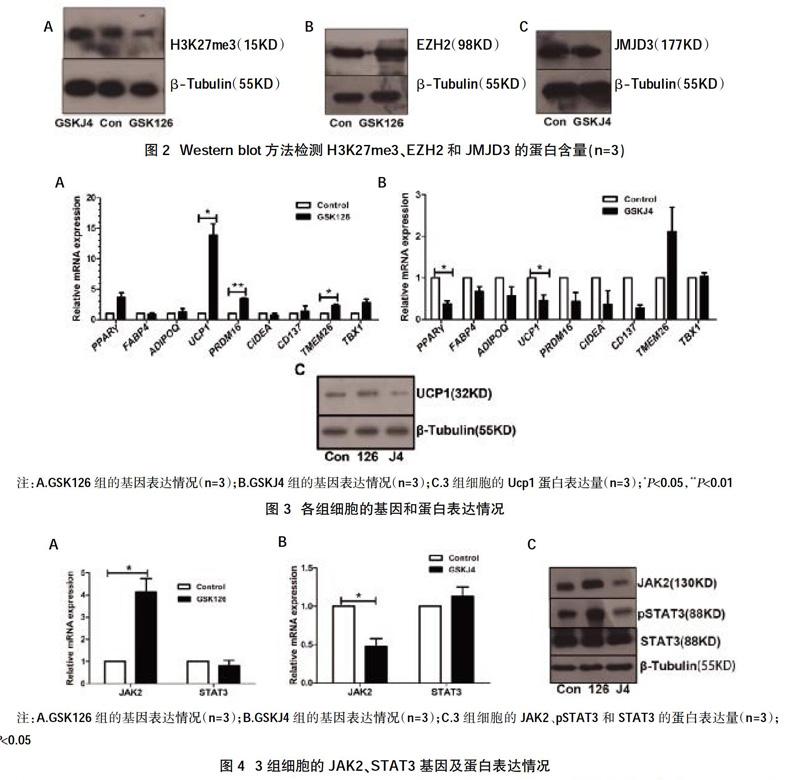
2.2抑制剂改变了H3K27me3甲基化修饰水平 经Western Blot检测,发现给药组H3K27me3的含量出现相应的改变,GSKJ4组该蛋白含量增加,H3K27三甲基化修饰水平增高;GSK126组该蛋白含量减少,H3K27三甲基化修饰水平降低,见图2A。接着我们测定了H3K27me3的甲基转移酶Ezh2和去甲基化酶Jmjd3的蛋白表达情况,结果显示,加入酶活性抑制剂后Ezh2和Jmjd3的蛋白表达均无明显改变,表明在酶蛋白含量无明显改变的情况下,两种蛋白的酶活性被部分抑制,见图2B,图2C。
2.3 GSK126组棕色脂肪标志基因表达增加,GSKJ4组的脂肪分化基因和棕色脂肪标志基因表达减少 我们检测了三组细胞的脂肪细胞分化基因PPARγ、FABP4和ADIPOQ;棕色化标志基因UCP1、PRDM16和CIDEA,以及米色脂肪特有的标志基因CD137、TMEM26和TBX1,并计算各组与对照组的相对表达量。结果显示,GSK126组的脂肪细胞分化基因没有明显改变,而GSKJ4组的PPARγ表达明显减少(P<0.05),见图3B。GSK126组的UCP1和PRDM16的表达明显升高(P<0.05,P<0.01),见图3A。GSK J4组的Ucp1表达明显减少(P<0.05),见图3B。另外,GSK126组的米色脂肪特有基因TMEM26的表达明显升高(P<0.05),见图3A。经Western blot检测,GSK126组的Ucp1表达升高,而GSKJ4组的Ucp1表达降低,见图3C。这些结果表明,GSK126促进了人MSC向棕色脂肪方向的分化,而GSKJ4抑制了MSC向棕色脂肪方向分化。
综上所述,H3K27me3的甲基化修饰在体外對于人脂肪来源间充质干细胞的成脂分化发挥了重要的调控作用:EZH2催化的H3K27me3修饰增加抑制MSC向棕色和米色脂肪细胞分化;JMJD3催化的H3K27me3修饰减少促进MSC向棕色和米色脂肪细胞分化。这些效应可能通过JAK2-STAT3信号通路发挥作用。
参考文献:
[1]Winik CL,Bonham CE.Implementation of a screening and management of overweight and obesity clinical practice guideline in an ambulatory care setting[J].Mil Med,2018,183(1-2):32-39.
[2]王醴湘,吕筠,郭彧,等.中国慢性病前瞻性研究:10个项目地区成年人超重/肥胖现况分析[J].中华流行病学杂志,2015,36(11):1190-1194.
[3]Gustafson B,Hedjazifar S,Gogg S,et al.Insulin resistance and impaired adipogenesis[J].Trends Endocrinol Metab,2015,26(4):193-200.
[4]Akinyemiju T,Moore JX,Pisu M,et al.A prospective study of obesity, metabolic health, and cancer mortality[J].Obesity,2018,26(1):193-201.
[5]Gonzalez-Hurtado E,Lee J,Choi J,et al.Fatty acid oxidation is required for active and quiescent brown adipose tissue maintenance and thermogenic programing[J].Molecular Metabolism,2018(7):45-56.
[6]Pan D,Huang L,Zhu LJ,et al.Jmjd3-mediated h3k27me3 dynamics orchestrate brown fat development and regulate white fat plasticity[J].Dev Cell,2015,35(5):568-583.
[7]Wang L,Jin Q,Lee JE,et al.Histone h3k27 methyltransferase ezh2 represses wnt genes to facilitate adipogenesis[J].Proc Natl Acad Sci USA,2010,107(16):7317-7322.
[8]Shi SY,Zhang W,Luk CT,et al.Jak2 promotes brown adipose tissue function and is required for diet- and cold-induced thermogenesis in mice[J].Diabetologia,2016,59(1):187-196.
[9]Gathirua-Mwangi WG,Song Y,Monahan PO,et al.Associations of metabolic syndrome and c-reactive protein with mortality from total cancer, obesity-linked cancers and breast cancer among women in nhanes iii[J].International Journal of Cancer,2018,143(3):535-542.
[10]Ishibashi J,Seale P.Medicine. Beige can be slimming[J].Science,2010,328(5982):1113-1114.
[11]Jaksic VP.Adipose tissue as an endocrine organ[J].Molecular & Cellular Endocrinology,2014,316(2):129-139.
[12]Cypess AM,Lehman S,Williams G,et al.Identification and importance of brown adipose tissue in adult humans[J].N Engl J Med,2009,360(15):1509-1517.
[13]Lidell ME,Betz MJ,Dahlqvist Leinhard O,et al.Evidence for two types of brown adipose tissue in humans[J].Nat Med,2013,19(5):631-634.
[14]汤其群.生物化学与分子生物学[M].上海:复旦大学出版社,2015:149-157.
[15]Ohno H,Shinoda K,Ohyama K,et al.Ehmt1 controls brown adipose cell fate and thermogenesis through the prdm16 complex[J].Nature,2013,504(7478):163-167.
[16]Harms MJ,Ishibashi J,Wang W,et al.Prdm16 is required for the maintenance of brown adipocyte identity and function in adult mice[J].Cell Metab,2014,19(4):593-604.
[17]Garcia RA,Roemmich JN,Claycombe KJ.Evaluation of markers of beige adipocytes in white adipose tissue of the mouse[J].Nutr Metab,2016,13(1):24-37.
[18]Cypess AM,Lehman S,Williams G,et al.Identification and importance of brown adipose tissue in adult humans[J].New England Journal of Medicine,2009,360(15):1509-1517.
[19]Asano H,Kanamori Y,Higurashi S,et al.Induction of beige-like adipocytes in 3t3-L1 cells[J].J Vet Med Sci,2014,76(1):57-64.
[20]Krone WJA,Stegehuis F,Koningstein G,et al.Ppar gamma agonists induce a white-to-brown fat conversion through stabilization of prdm16 protein[J].Cell Metab,2012,15(3):395-404.
[21]Sharp LZ,Shinoda K,Ohno H,et al.Human BAT Possesses Molecular Signatures That Resemble Beige/Brite Cells[J].PLOs One,2012,7(11):49452-49462.
[22]Wu J,Bostrom P,Sparks LM,et al.Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human[J].Cell,2012,150(2):366-376.
[23]Unno Y,Yamamoto H,Takatsuki S,et al.Palmitoyl lactic acid induces adipogenesis and a brown fat-like phenotype in 3t3-l1 preadipocytes[J].Biochim Biophys Acta Mol Cell Biol Lipids,2018,1863(7):772-782.
[24]Han DH,Nolte LA,Ju JS,et al.Ucp-mediated energy depletion in skeletal muscle increases glucose transport despite lipid accumulation and mitochondrial dysfunction[J].American Journal of Physiology Endocrinology & Metabolism,2004,286(3):347-353.
[25]Shi SY,Luk CT,Brunt JJ,et al.Adipocyte-specific deficiency of janus kinase (jak) 2 in mice impairs lipolysis and increases body weight, and leads to insulin resistance with ageing[J].Diabetologia,2014,57(5):1016-1026.
收稿日期:2018-11-3;修回日期:2018-11-13
編辑/杨倩
