Liver stem cells: Plasticity of the liver epithelium
2019-03-11AtsunoriTsuchiyaWeiYuLu
Atsunori Tsuchiya, Wei-Yu Lu
Abstract The liver has a high regenerative capacity after acute liver injury, but this is often impaired during chronic liver injury. The existence of a dedicated liver stem cell population that acts as a source of regeneration during chronic liver injury has been controversial. Recent advances in transgenic models and cellular reprogramming have provided new insights into the plasticity of the liver epithelium and directions for the development of future therapies. This article will highlight recent findings about the cellular source of regeneration during liver injury and the advances in promoting liver regeneration.
Key words: Liver regeneration; Stem cells; Cellular plasticity
INTRODUCTION
The liver is the largest internal organ in humans, and has exceptional regenerating capacity. The liver epithelium mainly consists of hepatocytes and cholangiocytes, with 70%-85% of the liver consisting of hepatocytes. Hepatocytes function in drug metabolism, protein synthesis and bile secretion. The biliary tree is a threedimensional branching structure lined by cholangiocytes to form tubular networks.These networks transport bile for storage in the gall bladder, or to the duodenum for the breakdown of fats. The liver is relatively quiescent compared to other epithelial organs such as the skin and intestines, with minimal hepatocyte proliferation during homeostasis[1,2]. The average lifespan of a hepatocyte ranges from 200-400 d[3]. It is believed that the liver has a ‘’two-tiered’’ regeneration system, where hepatocytes proliferate and regenerate the liver upon acute liver damage, and the liver progenitors or oval cells orchestrate the regeneration process during chronic liver injury when hepatocyte proliferation is impaired[4]. Recent advances in lineage-tracing and imaging techniques have provided new perspectives in addition to the original ‘’twotiered’’ idea. This review will focus on recent advances in stem cell biology and tissue repair to highlight the plasticity of the liver epithelium during regeneration.
REGENERATIVE POTENTIAL OF CHOLANGIOCYTES
The existence and the regenerative potential of hepatic progenitor cells (HPCs) or oval cells have been debatable. HPCs are a subpopulation of cells in the liver characterised by their oval shape and high nucleus to cytoplasm ratio when hepatocyte proliferation is inhibited[5]. Expansion of HPCs was observed in rat models of chronic liver injury in which hepatocyte proliferation is inhibited by the administration of 2-acetylaminofluorene (2-AAF). This was also observed upon hepatic injury such as carbon tetrachloride (CCl4) or partial hepatectomy (PHH)[6,7]. In addition to rat studies,HPC activation was also observed in mouse studies and human chronic liver disease[8-11]. In particular, the presence and contribution of a facultative liver stem cell population in human liver regeneration are highlighted by studies showing shared mitochondrial DNA mutations between HPC and the regenerative nodules in cirrhotic patients, suggesting their common origin[12,13].
HPC activation is a part of the dynamic change of the liver in response to chronic liver injury called ductular reaction (DR). Besides HPC activation, DR also includes immune cell infiltration, remodelling of the extracellular matrix, and myofibroblast activation[10,14,15]. Label-retaining assays that determine the cycling speed of the ductular cell populations have suggested multiple potential niches for slower cycling ductular cells after injury. These include the intralobular bile ducts, areas around periductal mononuclear cells, peribiliary hepatocytes, and the Canal of Hering[16].However, most studies that characterised DR relied on immunohistochemistry of paraffin-embedded sections, which has the limitation of being two-dimensional and lacking spatial information. This leads to the confusion that the DR observed is the activation of endogenous liver stem cells that leave their stem cell niche, migrate towards the site of injury and differentiate into hepatocytes. However, an elegant study performed by Kamimoto and colleagues used three-dimensional imaging techniques to demonstrate that instead of leaving their niche and migrating, the biliary tree undergoes complex remodelling without diverging from the main structure[17,18]. Furthermore, they also observed that cholangiocytes do not proliferate uniformly, which fits with the previous label-retaining studies suggesting heterogeneity in the proliferative capacity of cholangiocyte populations. These indicate the presence of HPC populations within the biliary epithelium with different regenerative potentials. However, the identity of HPCs within cholangiocytes remains elusive, as there are no specific markers to differentiate HPCs from cholangiocytes[19,20]. Nevertheless, subpopulations of cholangiocytes have been identified with markers such as Leucine-rich repeat-containing G-protein coupled receptor 5 (LGR5), ST14, Trop2, neural cell adhesion molecule (NCAM), MIC-1C3,CD133,etc[21-26].
Advances in transgenic mice and gene editing technologies in the past decades have facilitated lineage-tracing studies of cholangiocytes in the background of chronic liver injury, which is mainly induced by the administration of special diets. For example, the choline-deficient, ethionine-supplemented (CDE) diet and the methionine choline-deficient (MCD) diet can induce hepatocellular injury[8,27].Alternatively, cholestatic injury can be induced by the administration of a 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) diet[28]. Lineage-tracing studies using the Cre-lox system and traceable proteins such as GFP, RFP and B-galactosidase facilitate the identification of the origin and regenerative potential of cholangiocytes during homeostasis, chronic liver injury and post-recovery. However, lineage-tracing studies performed by multiple groups showed mixed results about the regenerative potential of cholangiocytes[24,29-31]. Rodrigo-Torreset al[29]and Espanol-Suneret al[30]used a tamoxifen-inducible Cre system controlled under the Hepatocyte Nuclear Factor 1B (HNF1B) and Osteopontin (OPN) promoters, respectively, to label cholangiocytes during homeostasis. They both showed that cholangiocytes can differentiate into hepatocytes following chronic liver injury. However, the contribution of biliary cells towards hepatocytes are minimal in these studies (<3%)[29,30]. On the contrary, when Cre is controlled under the transcription factor Forkhead box L1 (Foxl1) promoter, the degree of cholangiocyte-derived hepatocytes increases up to 29%, and ablation of the Foxl1 population results in impaired DR and liver regeneration[32-34]. The higher contribution from the Foxl1 population compared to the Hnf1b and the Opn population might be due to the former study’s use of a constitutive Cre system, and the latter study’s use of an inducible Cre system, in which the cell-labelling efficiency can be limited due to variable Cre penetrance. In contrast, the constitutive Cre system will always have the caveat that any cells that transiently expressed the promoter gene will be labelled, causing less accurate results.It is also worth noting that several studies do not show direct differentiation of cholangiocytes into hepatocytes following liver injury, which will be discussed in the sections below (Figure 1).
The existence of HPCs remains controversial, largely due to the mixed results obtained from different lineage-tracing studies[31,35,36]. Technical factors such as the selection of the promoter that controls the expression of Cre recombinase is one of the causes for this discrepancy. For example, it has been shown that some hepatocytes express Sox9 after tamoxifen injection or liver injury, which may result in the labelling of hepatocytes, potentially leading to the erroneous conclusion that overestimates the contribution from cells with biliary origin[35,37-39]. Furthermore, it is known that hepatocytes have a propensity to express certain biliary markers like osteopontin upon stress[40]. It seems that biliary markers such as Krt19 are an exception to this phenomenon, as Krt19 is not expressed by hepatocytes following injury, making it one of the most common markers used for lineage-tracing studies[39]. It is worth investigating whether other biliary markers such as EpCAM or Trop2 share similar characteristics to Krt19 during liver injury[25]. Another potential reason for the discrepancy might be the variable in injury models applied. For example, different studies have used different percentages of ethionine in water (0.1%-0.15%)[33,35]. The level of ethionine controls the severity of injury by inhibiting hepatocyte proliferation,which is crucial for the activation and differentiation of cholangiocytes into hepatocytes. The level of injury can also be affected by age, gender, the intake of ethionine-supplemented water and special diets. As the intake of diets is highly variable across individual mice, this might lead to the discrepancy observed due to the high regenerative capacity of hepatocytes. A recent study performed by Denget al[41]showed that Krt19-positive cells can differentiate into hepatocytes following longterm liver injury induced by the DDC diet. This shows the importance of selecting an appropriate injury model to investigate the true regenerative potential of cholangiocytes.
In comparison to dietary models, lineage-tracing studies involving liver injury induced by transgenic approaches appears to result in a more robust contribution of hepatocytes from non-hepatocyte sources[42,43]. Transgenic approaches to induce hepatocyte senescence through the deletion ofMdm2, a negative regulator of p53, has been developed to simulate human chronic liver disease. In this system, where the inducible Ah-Cre is controlled by the Cyp1a promoter, hepatocytes can be targeted by the administration of B-Naphthoflavone to trigger a senescence phenotype, hence causing a robust DR and the regeneration of the liver parenchyma contributed by a non-hepatocyte source[42,44]. Besides inducing cellular senescence, hepatocyte proliferation can also be disrupted by the deletion of theItgB1gene[45]. Hepatocytes that have defective ItgB1 have impaired proliferation due to disrupted HGF signalling[45]. Overexpression of p21, a cyclin-dependent kinase inhibitor, has also been used to impair hepatocyte proliferation through the delivery of AAV8 vector packaged with the p21 codon[43]. When combining the inhibition of hepatocyte proliferation (Itgb1knockout or AAV8-p21 delivery) with dietary-induced chronic liver injury, biliary cells can commit to both biliary and hepatocyte fates[43]. Although the contribution of cholangiocytes into hepatocytes does not lead to full repopulation,it is still a significant contribution (25%) compared with previous models.Furthermore, suppression of the Wnt/β-catenin pathway in hepatocytes promotes the differentiation of cholangiocytes into hepatocytes[46]. However, it remains unclear whether full repopulation of the liver from cholangiocytes can be achieved. The loss of ItgB1, the Wnt pathway or p21 overexpression in all hepatocytes do not completely resemble the mechanisms of human liver disease. Nevertheless, the impairment of hepatocyte regenerative capacity promotes the differentiation of cholangiocytes as a proof-of-principle to demonstrate the regenerative capacity of cholangiocytes. Despite this, the complex regulatory process that triggers cholangiocytes to differentiate into hepatocytes remains to be identified. It was recently reported that Histone deacetylase 1 (HDAC1) regulates the commitment of cholangiocytes to biliary epithelial and hepatocyte fates through controlling Sox9 expression in zebrafish and mouse. This reveals extra molecular pathways that work in conjunction with the previously identified Wnt and Notch pathways in regulating liver regeneration[14,47]. Nevertheless,it remains to be tested whether promoting endogenous regeneration through the activation and differentiation of biliary cells to repopulate the liver parenchyma is plausible.
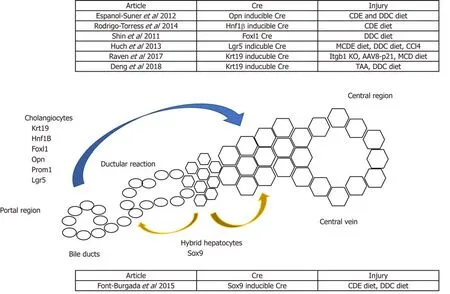
Figure 1 Schematic summarising potential non-hepatocyte sources of regeneration. Lineage-tracing models using Cre recombinase controlled under a cholangiocyte-specific promoter have shown cholangiocytes giving rise to hepatocytes. Alternatively, Sox9+ hybrid hepatocytes have been shown to give rise to both cholangiocytes and hepatocytes.
Besides the advances achieved in lineage-tracing studies, the development of the organoid culture system pioneered by the Clevers group revolutionised the field of regenerative medicine. First established in 2009 by Satoet al[48], organoid culture was used to culture intestinal stem cells that express the Wnt target gene leucine-richrepeat-containing G protein-coupled receptor 5 (Lgr5). This technology was then applied by the same group to the liver to form liver organoids, and is now widely used by researchers forin vitromodelling, drug screening and gene sequencing[24,49,50].Endogenous lineage-tracing of Lgr5-expressing cells using the Lgr5-IRES-creERT2 reporter mice showed that Lgr5-expressing cells can contribute to both cholangiocytes and hepatocytes after liver damage[24]. Interestingly, Lgr5 expression is not detected in healthy liver, but only detected in cholangiocytes after injury. This indicates that Lgr5 is transiently expressed in a subpopulation of cholangiocytes that activate Wnt signalling and repopulate the liver when required. However, the origin and identity of this trans-amplifying population in the liver during quiescence remains to be investigated. Identifying the origin of the Lgr5 population will reveal whether either there are predetermined residential liver stem cells, or cholangiocytes obtain regenerative capacity stochastically and regenerate the liver parenchyma during injury. Although the controversy regarding the existence of a dedicated liver stem cell population remains, subpopulations of cholangiocytes have been identified.Furthermore, it appears that the cells of the liver epithelium (i.e. cholangiocytes and hepatocytes) can obtain cellular plasticity and differentiate into the other compartment when one compartment is significantly damaged.
EXOGENOUS REPOPULATING CAPACITY OF CHOLANGIOCYTES
In contrast to lineage-tracing studies, there are more convincing results in transplanting isolated orin vitro-cultured cholangiocytes[42,51-53]. Transplantation assays have been widely used by the liver research community to prove the repopulating capacity of different cell populations. As the most effective treatment for liver disease is whole organ transplantation, it is common to seek alternatives using cells isolated from the liver to overcome the challenges faced in allogenic transplantations. Most transplantation assays are designed in a way that selective advantages are provided to the donor population, and this is often achieved by causing defects to the host hepatocytes. The most widely used models are the fumarylacetoacetate hydrolasemice(Fah)-deficient and the urokinase-type plasminogen activator-overexpressing (uPA)mice[54,55].
The Fah-deficient mice lack the functional fumarylacetoacetate hydrolase (Fah), a metabolic enzyme required for the last step of the tyrosine catabolism pathway. Fah mice develop liver diseases and die within 12 hours after birth from hypoglycaemia[54].This defect can be corrected by blocking the accumulation of hepatotoxins through the administration of 2-(2-nitro-4-trifluoro-methylbenzyol)-1,3-cyclohexanedione (NTBC)to partially recover liver function[56]. The administration of NTBC prolongs the lifespan of Fah mice and enables this strain to be suitable for liver repopulation[57]. On the other hand, the uPA transgenic mice have the mouse urokinase-type plasminogen activator (uPA) gene under the control of the mouse albumin enhancer/promotor.Hepatocyte-specific expression of uPA causes liver toxicity and chronic liver injury,hence creating a selective advantage for transplanted hepatocytes[55,58]. However, the limitations of this model are the narrow time transplantation window after birth and the poor breeding efficiency of this strain. In addition, only homozygous animals are good recipients for liver cell transplantation[59]. Immunocompromised variants of these strains have also been developed, and transplantation of human or rodent hepatocytes has shown promising repopulation capacity after xenotransplantation[60-62].
With hepatocyte transplantation as the gold-standard for the liver-repopulating transplantation assay, the liver-repopulating capacity of isolated HPCs were investigated using similar assays. HPCs can be enriched by density gradient centrifugation from the liver non-parenchymal fraction after protease digestion[21,25,53].Fluorescence-activated cell sorting (FACS) is often used to isolate a pure HPC population. However, HPCs do not have a specific marker and often share surface markers (e.g., c-kit, Thy-1) with other cell types such as haematopoietic cells[6,63,64]. This raised doubts about whether HPCs originate from the bone marrow, but transplantation assays and lineage-tracing studies have proven that HPCs originate from cholangiocytes and not the bone marrow[42,53]. Multiple surface markers have been used to isolate cholangiocyte subpopulations with liver-repopulating capacity,which include EpCAM, Trop2, CD24, CD133, LGR5, c-kit, CD44, Thy-1,etc[22,25,42,51,53,57].Due to the relatively low numbers of isolated cells, most studies culture the isolated cellsin vitrobefore transplantation. Upon transplantation, cells can differentiate into hepatocytes and self-renew[21,22,24,25,42,51,53,63]. However, the magnitude of repopulation is still relatively low compared to primary hepatocyte transplantation, which remains a challenge for using HPCs in cell therapy[65]. Refinements to increase the degree of repopulation, either by targeting the engraftment efficiency or the differentiation capacity, are required. Nevertheless, an advantage of using biliary-derived cells for transplantation is that cholangiocytes are more resistant to protease digestion than hepatocytes and can be culturedin vitro, which is difficult to achieve for primary hepatocytes. In addition to regenerating damaged hepatocytes, human liver organoids derived from cholangiocytes have also been used to regenerate extrabiliary bile ductsin vivoafter being seeded with biodegradable scaffolds[66]. The use of cultured cholangiocytes to regenerate damaged liver epithelium seems promising.However, the heterogeneity of cells within the culture, and whether prolonged culture alters the characteristics and the long-term stability of the cells, need to be further investigated.
THE PLASTICITY OF HEPATOCYTES
The liver is deemed a highly regenerative organ mainly due to the remarkable regenerative capacity of hepatocytes. The proliferative capacity of hepatocytes is well characterised such that when acute liver injury occurs, hepatocytes initiate a series of pathways to restore the lost mass[67]. These studies are mostly performed using the PHH model, where a substantial portion of the liver is resected. It was commonly presumed that proliferation is the main mechanism used by hepatocytes to compensate for the loss of liver mass. However, a study by Miyaoka and colleagues show that hepatocyte hypertrophy (i.e. increase in hepatocyte size) contributes concurrently with proliferation to compensate for the loss of liver mass[68,69]. However,this regeneration process does not promote regrowth of the original resected lobes,but instead, the replacement of the original mass occurs at the remnant lobes. It has been showed that YAP/Hippo signalling plays a major role in controlling the restoration of the liver mass to its original size[70]. However, it needs to be noted that in the PHH model, the remnant lobes are not affected by any injury, which is uncommon in most clinical cases of acute liver failure. Patients have substantial inflammatory effects or senescence in the liver, such as paracetamol overdose.
The use of the Adeno-associated Virus (AAV) vector, a small (approximately 5 kb long), non-enveloped virus for transgene delivery specifically into hepatocytes, allows researchers to lineage-trace hepatocytes during homeostasis and injury[71]. Gaoet al[72]isolated AAV serotype 8 (AAV8), which has a 10-100-fold higher liver tropism compared to other AAV serotypes. Malatoet al[71]used the hepatotropism characteristic of the AAV8 to introduce Cre recombinase controlled under the hepatocyte-specific transthyretin (Ttr) promoter to target hepatocytes, which has a higher specificity than the widely used Alb-Cre. They lineage-traced the labelled hepatocytes under the context of homeostasis, acute liver injury, and biliary injury.During homeostasis and acute liver injury, they did not find any evidence that hepatocytes are derived from the biliary compartment. However, they observed a small amount of biliary-derived, periportal hepatocytes following bile duct ligation and DDC injury, suggesting a population within the cholangiocyte population that can differentiate into hepatocytes following injury. However, this conclusion is based on the authors’ observation of unlabelled hepatocytes located at the periportal region,instead of positively lineage-traced biliary cells. The same group went further to investigate whether cholangiocytes can differentiate into hepatocytes following chronic liver injury using the CDE diet regime, which predominantly damages hepatocytes[36]. Surprisingly, in contrast to the DDC model, the authors observed minimal (less than 1%) production of hepatocytes by cholangiocytes after chronic liver injury. This was further confirmed with a positive lineage-tracing model using CK19-CreER;R26R-RFP mice. Furthermore, the authors also showed that mesenchymal cells cannot differentiate into hepatocytes after chronic liver injury. Yangeret al[31]further investigated the source of newly formed hepatocytes following chronic liver injury using CK19-CreER; R26R-YFP mice by applying different types of injury, including DDC, CDE and CCl4administration, as well as an alpha-naphthyl-isothiocyanate(ANIT) diet. Similar to what Schaubet al[36]observed, they did not detect any production of hepatocytes from YFP-labelled biliary cells after injury, regardless of the timing of cell labelling before or during injury. These suggest that quiescent cholangiocytes or activated atypical duct cells cannot differentiate into hepatocytes following injury. They further characterised the source of regenerating hepatocytes following liver injury using an AAV8 vector packaged with a Cre recombinase controlled under a hepatocyte-specific Thyroid Binding Globulin promoter (AAV8-TBG-Cre). Hepatocytes from R26R-YFP mice are labelled with YFP after AAV8-TBGCre administration. They observed no reduction in the amount of YFP-labelled hepatocytes following treatment with the panel of injury models mentioned above,including the widely used CDE and DDC. This suggested that the newly formed hepatocytes derived from self-duplication of pre-existing hepatocytes rather than from other cell populations[31].
These observations require further follow-up investigation as to whether there is a population of hepatocytes that possess a higher regenerative capacity[31,35,36,71]. Wanget al[73]reported that a population of pericentral hepatocytes that express the Wnt responsive geneAxin2migrate towards the periportal region during homeostasis.This Axin2+ population also expresses the hepatoblast marker Tbx3, and differentiates into Tbx3-negative hepatocytes. Furthermore, the Axin2+ population proliferates faster, is mostly diploid and its Axin2 expression is dependent on the Wnt signals provided by the endothelium at the central vein. This reawakened the streaming liver hypothesis proposed by Zajiceket al[74]30 years ago, which claims that hepatocytes from the portal region gradually advance towards the central region during homeostasis. In this study, however, the stream starts from the opposite direction instead (Figure 2). However, recent studies showed that despite the presence of Wnt activity in the pericentral hepatocytes, this population does not possess proliferative advantages over other hepatocyte populations during homeostasis and acute injury[75]. Instead, proliferation rates across the three acinar zones are similar,and regeneration is performed by hepatocytes with high levels of telomerase expression[76].
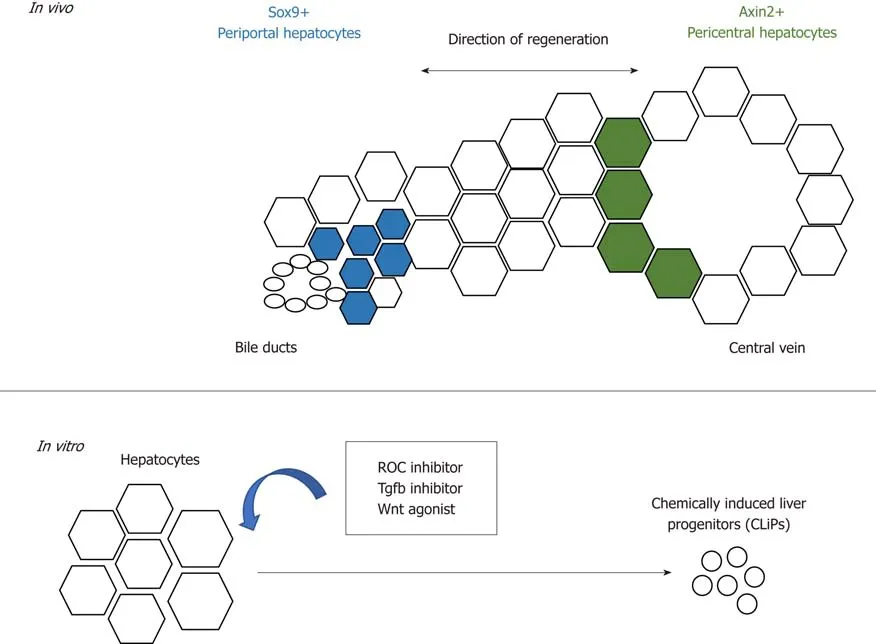
Figure 2 Schematic summarising the regenerative potential of hepatocytes.In vivo, periportal and pericentral hepatocytes regenerate the liver with different regenerative mechanisms. In vitro, hepatocytes can be converted into liver progenitors with extrinsic factors.
Another report by Font-Burgadaet al[77]suggests that a population of hepatocytes named hybrid hepatocytes (HybHP) resides close to the biliary epithelium and expressesSox9during homeostasis. In both chronic liver injury models of CCl4administration as well as the MUP-uPA transgenic model, HybHP repopulate the damaged population and manage to differentiate into zone 3 hepatocytes with the expression of glutamine synthetase. Interestingly, under the CDE diet regime, they found that most HybHP were killed, but no contribution of hepatocytes from the biliary cells was observed albeit the existence of extensive DR. Furthermore, they found HybHP can transdifferentiate into biliary-like cells that express biliary markers like Sox9, Opn, and Ck19 when induced with cholestatic injury from a DDC diet.Transcriptional analysis revealed the hybrid character of HybHP, as this population expresses a cluster but not all of genes that are usually expressed by cholangiocytes.However, it remains to be investigated whether this population can fully differentiate into mature cholangiocytes, and this might explain the differences in the observations made by Malatoet al[71]and Yangeret alabout whether hepatocytes can differentiate into cholangiocytes after injury. Upon transplantation into the immunodeficient Fah-/-Rag2-/-Il2rg-/- transgenic model, HybHP exhibit higher repopulation capacity than conventional hepatocytes or HPC. Despite their regenerative capacity, neither HybHP nor HPC preferentially give rise to hepatocellular carcinoma (HCC), suggesting tumorigenesis is a random event that occurs equally in all cell types rather than preferentially in certain cell populations. In addition, the HybHP phenotype was also observed by Tarlowet al[78]after chronic liver injury, in which hepatocytes can obtain a biliary phenotype with the expression of the cholangiocyte marker MIC-1C3 (hepPD).Although they share similar surface marker expression, the transcriptional profile of hepatocytes and biliary-derived MIC-1C3-expressing cells are different, and these two populations behave differentlyin vitro, with the former maintaining a more hepaticlike profile. These Sox9-expressing hepPD cells can also differentiate back into hepatocytes after injury[78]. Both the Tarlow and Font-Burgada studies demonstrated the plasticity of hepatocytes after injury, and the heterogeneity of hepatocytes within the liver does not only restrict to their zonation and drug metabolic functions, but also occurs in the context of their regenerative potential. Interestingly, the conversion of hepatocytes into biliary-like cells was observed in the DDC model but not in the Thioacetamide (TAA) model[18]. This might due to the pathophysiology caused by different injury models, in which the DDC is a model for cholestatic liver injury whilst the TAA model is a hepatotoxic injury model. This highlights the plasticity in the liver when cholestatic injury occurs, and that hepatocytes can transdifferentiate into biliary-like cells due to damage in the biliary compartment. On the other hand, when hepatocyte proliferation is impaired, cholangiocytes can differentiate into hepatocytes.
The plasticity of hepatocytes after chronic liver injury is also recapitulatedin vitroby the conversion of mature hepatocytes in mice and rats into progenitor-like cells using a cocktail of small molecules that consists of ROCK-inhibitor(Y-27632), TGF-β inhibitor (A-83-01) and Wnt agonist (CHIR99021)[79](Figure 2). The chemicallyinduced liver progenitors (CLiPs) can be cultured long-term, as well as differentiate into hepatocytes and biliary cellsin vitroandin vivoafter transplantation. The development of CLiPs has overcome the limitation that primary hepatocytes could not maintained be in prolonged culturein vitro, and the low cell number of the regenerative progenitor cellsin vivo. This opens a new possibility of using CLiPs for cell therapy. Whether all hepatocytes or only a subpopulation can be reprogrammed into CLiPs, and the applicability of using human hepatocytes for CLiPs generation,remains to be investigated in order to translate this into future therapy.
CELLULAR REPROGRAMMING FOR LIVER REGENERATION
The 2006 discovery of induced pluripotent stem cells (iPSCs) by the expression of Yamanaka factors (Oct3/4, Sox2, Klf4, c-Myc) in fibroblasts sparked broad interest in cellular reprogramming[80]. Hepatocyte-like cells have been generated from embryonic stem cells (ESCs) and iPSCs using multistep protocolsin vitro[81-83]. When co-cultured with other cell types such as mesenchymal and endothelial cells, iPSC-derived hepatocytes can form three-dimensional ‘’liver buds’’. The liver buds can form vascular networks with the host vasculature after cranial transplantation[84]. These cells are potentially promising for drug screening, disease modelling, and cell replacement therapy. However, ethical issues, efficacy and long-term stability of these cellsin vivoremains a challenge to translate these into therapy[85,86]. Besides using ESC/iPSC-derived hepatocytes, researchers also considered the possibility of bypassing the reprogramming into pluripotency by shortening the multistep reprogramming protocol, or even directly converting adult somatic cells into functional hepatocytes[87,88]. The overexpression of two transcription factors including Hnf4a with the combination of either Foxa1, Foxa2, or Foxa3 can induce reprogrammingin vitroof cultured mouse embryonic and adult fibroblasts into hepatocyte-like (iHep) cells. These cells maintained hepatic functionin vitroand contribute to the regeneration of the liver parenchyma after transplantation[88]. Direct reprogramming of hepatic myofibroblasts into hepatocytesin vivowith adenovirus has also shown decreased fibrosis in mice[89]. Advances in cellular reprogramming have opened a new area of regenerative medicine. However, the efficiency, epigenetic memory and stability of transdifferentiated hepatocytes remains a challenge for the therapeutic use of cells generated by genetic reprogramming.
CONCLUSION AND FUTURE DIRECTIONS
A lot has been learned about liver plasticity in recent years, especially regarding hepatocytes. It appears that cellular plasticity in the liver is bidirectional and injurydependent, where hepatocytes are able to transdifferentiate into biliary-like cells under certain conditions. However, when the proliferative capacity of hepatocytes is impaired, cholangiocytes become the facultative liver stem cells that differentiate into hepatocytes. It remains to be investigated whether compartmentalised stem cell populations exist within the liver as in other epithelial organs, or if the acquisition of stem cell properties by cholangiocytes is a stochastic event[90,91]. Nevertheless, it seems that the plasticity of hepatocytes and cholangiocytes reflects what has been observed in other organs such as the skin and intestines, where residential stem cells acquire plasticity and differentiate after injury[90,92]. Future studies involving next generation sequencing at the single-cell level will provide us with new perspectives on cell identity and the involvement of epigenetic alterations during regeneration.Advancements in our understanding of tissue repair, cellular reprogramming and bioengineering will be beneficial for developing models for drug screening, and may even translate into cell therapy. Modulation of the regenerative niche will hopefully enhance the endogenous regenerative capacity of the liver (Figure 3).
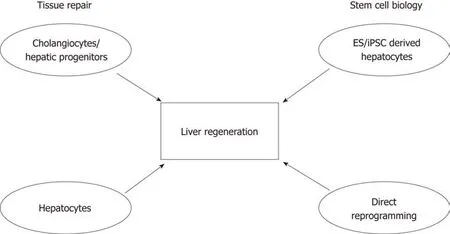
Figure 3 Schematic showing the importance of understanding tissue repair and stem cell biology, and their contributions to developing future therapies to promote liver regeneration. Studies focusing on understanding the mechanisms of tissue repair have suggested that cell populations with high cellular plasticity include both cells of the biliary tree and hepatocytes. The fields of stem cell biology and regenerative medicine have used signals and key factors required during embryonic development to produce hepatocyte-like cells derived from either adult or embryonic sources. In conjunction, these will advance the liver regeneration field.
杂志排行

World Journal of Gastroenterology的其它文章
- Reaction of antibodies to Campylobacter jejuni and cytolethal distending toxin B with tissues and food antigens
- Integrated network analysis of transcriptomic and protein-protein interaction data in taurine-treated hepatic stellate cells
- Computed tomography scan imaging in diagnosing acute uncomplicated pancreatitis: Usefulness vs cost
- Targeted puncture of left branch of intrahepatic portal vein in transjugular intrahepatic portosystemic shunt to reduce hepatic encephalopathy
- Optimized protocol of multiple post-processing techniques improves diagnostic accuracy of multidetector computed tomography in assessment of small bowel obstruction compared with conventional axial and coronal reformations
- Comprehensive lifestyle intervention vs soy protein-based meal regimen in non-alcoholic steatohepatitis