刺萼龙葵土壤种子库特征及其对替代控制的响应*
2019-03-08张瑞海张国良王忠辉付卫东尹树红翟洪凯席坤鹏
张瑞海, 宋 振, 张国良, 王忠辉, 付卫东**, 尹树红, 翟洪凯, 席坤鹏
刺萼龙葵土壤种子库特征及其对替代控制的响应*
张瑞海1, 宋 振1, 张国良1, 王忠辉1, 付卫东1**, 尹树红2, 翟洪凯2, 席坤鹏2
(1. 中国农业科学院农业环境与可持续发展研究所 北京 100081; 2. 白城市农业环境保护与农村能源管理站 白城 137000)
土壤种子库在退化生态系统植被的恢复和演替中起重要作用, 为明确外来入侵植物刺萼龙葵(Dunal.)土壤种子库特征, 评价植物替代控制刺萼龙葵的效果, 调查了河滩及农田边缘两个典型生境中的刺萼龙葵土壤种子库特征, 分析了刺萼龙葵种子在土壤中的垂直分布与季节变化动态, 并利用多种多年生禾本科与豆科牧草对刺萼龙葵进行替代控制研究, 以期为刺萼龙葵生态调控提供理论依据。结果表明: 1)两个生境中共鉴定出23种植物, 禾本科和菊科为优势科; 河滩及农田边缘刺萼龙葵种子总储量分别达347粒∙m-2和2 600粒∙m-2, 占整个种子库的2.46%及35.16%。2)河滩生境中刺萼龙葵种子多集中于表层(0~2 cm)土壤, 占种子总量的64.3%, 且随土层深度的增加而减少; 而农田边缘生境0~2 cm、2~5 cm和5~10 cm 3层种子数量差异不大(>0.05), 分别占种子总量的32.7%、38.2%及29.1%。3)两个生境中刺萼龙葵种子主要集中于4月份采集的土样中, 分别为273粒∙m-2(河滩)和1 970粒∙m-2(农田边缘), 显著高于6月份与8月份采集的土样种子数(<0.05); 4)从替代控制第2年起, 刺萼龙葵的密度、生物量及土壤种子储量即被控制在较低水平, 均显著低于同期对照(<0.05), 沙打旺+苇状羊茅+冰草+羊草组合对刺萼龙葵控制效果最佳, 同时还可获得牧草鲜重20 396.1 kg∙hm-2, 干重7 710.6 kg∙hm-2(2017年), 经济效益可观。5)刺萼龙葵种子库与刺萼龙葵密度(<0.01)、生物量(<0.05)呈正相关, 牧草产量与刺萼龙葵种子库储量、密度、生物量均呈负相关, 但相关性不显著(>0.05); 降雨显著影响刺萼龙葵种子库储量(<0.05), 可用幂函数模型=2.6190.001(20.822,=18.486,=0.013)描述二者之间的关系。
生物入侵; 刺萼龙葵; 入侵植物; 土壤种子库; 时空特性; 替代控制
土壤种子库(soil seed bank)是指存在于确定面积的土壤上层凋落物和土壤中全部种子的总和[1-2], 是植物潜在的种群, 研究土壤种子库的组成、动态及其在植被恢复和演替中的作用, 有助于人类在退化生态系统修复方面做出合理的决策和措施[3]。近年来外来植物入侵已成为全球性生态问题[4], 许多入侵植物繁殖系数高, 其种子的萌发、种苗的补充以及群落的重建是其繁殖扩散的关键[5]。在受外来物种入侵危害地区, 土壤种子库结构的变化决定着入侵植物未来的危害程度, 而且对当地生态系统的演替和植物群落的结构、功能及组成的影响尤为重要[6-7]。
刺萼龙葵(Dunal.)为茄科(Solanaceae)茄属一年生草本植物, 原产于北美洲[8]。1981年首次在我国辽宁朝阳被发现[9], 随后扩散至吉林、山西、河北、北京、新疆、内蒙古等地, 并形成优势群落, 造成本地生物多样性降低, 入侵农田后造成农作物减产[10-13]。刺萼龙葵全株带刺且有毒, 牲畜误食后可引起中毒死亡[14]。刺萼龙葵繁殖力强, 单株结实量可达1万~2万粒, 其种子具有物理休眠(种皮的机械阻碍作用)和生理休眠(胚中存在抑制萌发的物质)混合休眠特性[15-16], 这使得刺萼龙葵的防除难度增加。目前针对刺萼龙葵的研究侧重于分布与危害、生物学与生态学特性、转化利用、风险评估等方面[17-23], 为防控刺萼龙葵提供科学依据。
关于刺萼龙葵防治主要集中于化学防治与替代控制方面的研究, 其中化学防治对刺萼龙葵能起到一定的控制作用[24], 但没有专一性强的除草剂, 防除刺萼龙葵的同时, 易对当地环境造成危害, 因此, 化学防治刺萼龙葵并不是有效的可持续性控制技术。农业部环境保护科研监测所曾公布过利用草木樨(L.)+冰草(L.)+羊草[(Trin.) Tzvel.]控制刺萼龙葵的方法, 对刺萼龙葵取得了较好的控制效果[25], 但并未报道替代控制对刺萼龙葵种子库的影响。本研究以此为切入点, 首先调查了农田边缘与河提两个典型入侵生境中的刺萼龙葵土壤种子库分布特征, 并对替代控制措施后刺萼龙葵种子库进行了研究, 明确刺萼龙葵种子在土壤分布中的规律及利用替代控制措施对刺萼龙葵种子库的影响, 为有效综合治理刺萼龙葵提供科学依据。
1 材料与方法
1.1 刺萼龙葵入侵生境种子库特征调查
选取两个刺萼龙葵发生的典型生境, 生境一为河滩, 位于辽宁省朝阳市大凌河(41°32′N, 120°27′E),海拔170 m, 全年平均气温9.0 ℃, 年平均降水量475 mm, 土壤类型为风砂土。生境二为农田边缘, 位于河北省万全(44°44′N, 114°45′E), 海拔716 m, 年平均气温6.9 ℃, 年平均降水量464 mm, 土壤类型为潮土。
在上述两个生境中, 采取对角线取样方法取样, 在每个生境中选取30个样方(面积为100 cm×100 cm), 在样方内随机取1个10 cm×10 cm的小样方, 分层取样, 取样深度依次为0~2 cm(上层)、2~5 cm(中层)和5~10 cm(下层)。根据刺萼龙葵生长周期确定3次取样时间, 分别为: 刺萼龙葵种子萌发前(2012年4月5日)、刺萼龙葵种子萌发中期(2012年6月29日)及刺萼龙葵种子成熟脱落前(2012年8月28日)。
将取回的土样过40目筛滤出枯枝、石块等杂物, 均匀平铺于20 cm×15 cm(内直径×高)的盆钵中, 置于温室中, 隔离外界种子干扰, 定期浇水, 逐日记录种子发芽数, 对已萌发出的幼苗进行种类鉴定、计数后清除。如连续两周无种子萌发, 再将土壤搅拌混合, 继续观察, 直至连续两周不再有种子萌发, 然后将土壤过筛, 统计未萌发的刺萼龙葵种子数量。
土壤种子库储量以密度来表达, 在本研究中, 刺萼龙葵土壤种子库大小表达为单位面积中有活力的刺萼龙葵种子数量。
1.2 替代控制对刺萼龙葵种子库的影响
试验区设在吉林省白城市洮北区平台镇(45°57′N, 122°85′E), 位于科尔沁自然草原北部, 海拔180 m, 全年平均气温4.9 ℃, 年平均降水量408 mm, 土壤类型为淡黑钙土。试验开始前, 于2012年6月27日调查试验区已出苗的刺萼龙葵密度[(292.4±102.5)株∙m-2]以及土壤中刺萼龙葵种子库储量[0~2 cm: (4 109±2 826) 粒∙m-2; 2~5 cm: (2 065± 1 099) 粒∙m-2; 5~10 cm: (3 827±1 820) 粒∙m-2], 此时刺萼龙葵处于2~6叶期, 株高1~4 cm。
试验地翻耕后, 划定10个面积为384 m2(24 m×16 m)的试验区, 每个试验区分为3个面积为128 m2(24 m×8 m)的小区。根据当地气候及前期筛选, 选取披碱草(Turcz., Ed)、无芒雀麦(Leyss., Bi)、苇状羊茅(Schreb., Fa)、羊草(Lc)、冰草(Ac)、紫羊茅(L., Fr)、紫花苜蓿(L., Ms)及沙打旺(Pall., Aa) 8种牧草作为替代植物, 每小区中替代植物组合及播种量见表1。播种方法为混匀后撒播, 播种完毕后, 在整个试验区四周设置围栏及标牌, 以防止人为及牲畜破坏。对照区组(与试验区间隔100 m)不进行撒播替代植物种子。整个试验过程中不施肥, 不浇灌水(自然降水), 试验从2012年6月始到2017年10月止, 每年10月在每个小区中随机选择5个1 m2的样点, 按照1.1方法统计各小区土壤中刺萼龙葵种子库大小, 计算平均值。

表1 替代植物试验处理及播种方法
其余指标测定方法: 2012—2017年每年8月在每个小区中随机选择5个1 m2的样点, 调查测定刺萼龙葵密度及单株生物量。刺萼龙葵密度计算方法: 在每个小区内, 随机选择5个1 m2的样点, 统计该样点内的刺萼龙葵植株数量, 取平均值。生物量测定方法: 用烘箱在105 ℃杀青15 min后, 在80 ℃烘干至恒重。并于2015年8月及2017年8月, 在每个小区中随机选择5个1 m2的样点进行刈割处理, 收获全部牧草, 测量牧草鲜重(FW)和干重(DW)。
1.3 数据分析
多组数据采用One-Way ANOVA方差分析, 所有数据统计与分析、相关分析及回归分析利用Microsoft Excel 2013和SPSS 19.0软件完成, 并在Origin 9.1完成作图。
2 结果与分析
2.1 土壤种子库种类特征
2种生境土壤种子库共鉴定出23种植物, 隶属于10科22属。其中禾本科(Poaceae)、菊科(Asteraceae)植物为优势科。河滩生境有18种植物, 耕地生境有19种植物, 两个样地共有物种有14种。表2和表3分别列出了两种生境样地中土壤种子库各物种组成及其所占储量比例。河滩生境中刺萼龙葵种子量较少, 而农田边缘生境中刺萼龙葵种子库储量最多, 为优势种群。两个生境中, 刺萼龙葵种子量分别占种子总储量的2.46%和35.16%。刺萼龙葵种子库表现为农田边缘(2 600粒·m-2)>河滩(347粒∙m-2)。除刺萼龙葵外, 河滩生境样地中另一种外来入侵植物少花蒺藜草(Cav.)种子量最多, 占总储量的36.70%, 种子库大小达5 187粒∙m-2。
2.2 刺萼龙葵种子在土壤中的垂直分布
从图1可以看出, 在垂直方向上, 河滩生境中随着土壤深度的增加刺萼龙葵种子数量减少(<0.05), 0~2 cm、2~5 cm 和5~10 cm 3层土壤平均分别为223粒∙m-2、86粒∙m-2及38粒∙m-2, 其中上层(0~2 cm)刺萼龙葵种子占种子总量的64.3%; 而在农田边缘生境3层土壤中刺萼龙葵种子数差异不大(>0.05), 平均分别为850 粒∙m-2(0~2 cm)、994粒∙m-2(2~5 cm)和756粒∙m-2(5~10 cm), 分别占种子总量的32.7%、38.2%和29.1%。
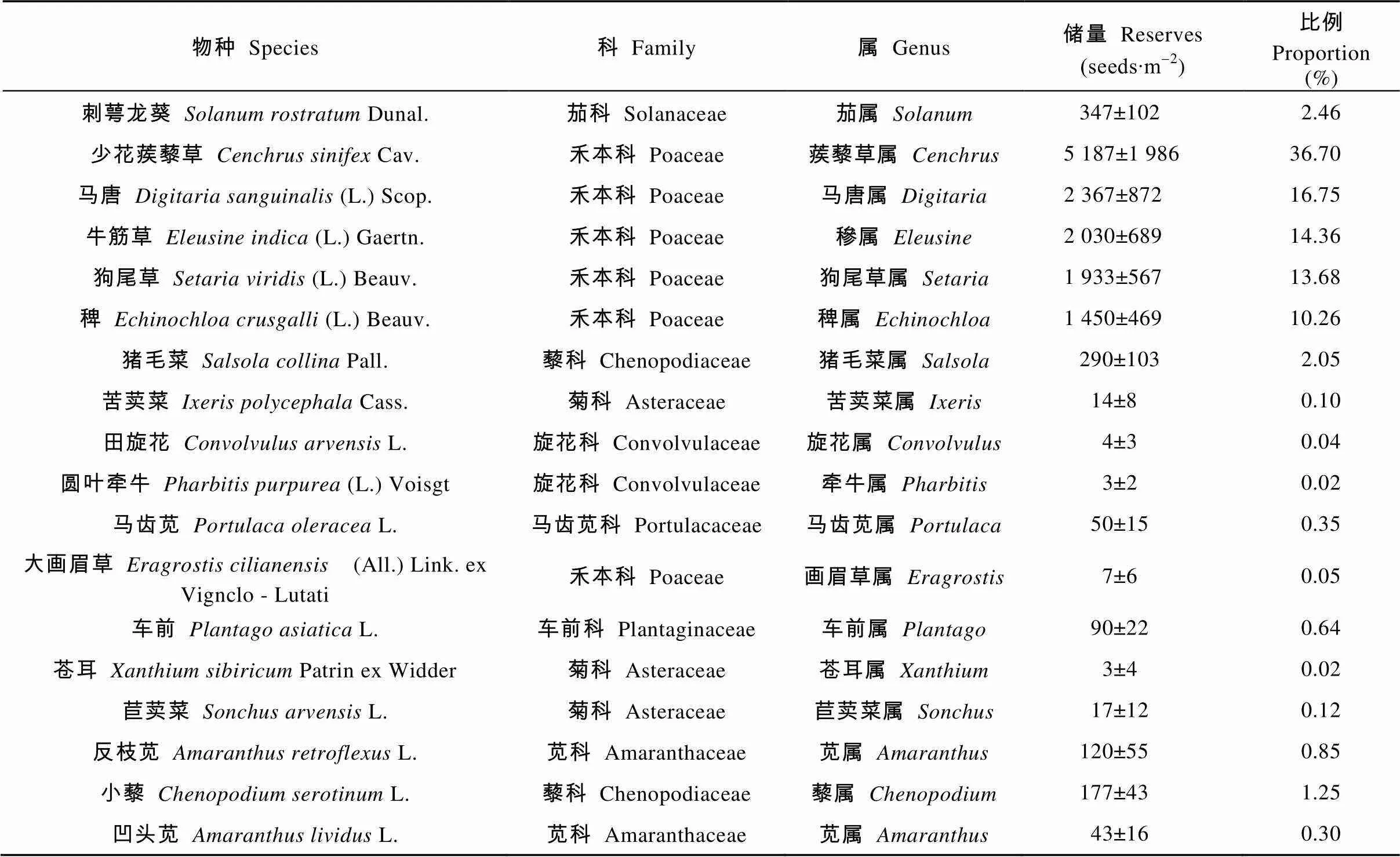
表2 朝阳河滩土壤种子库各物种组成及其所占储量比例

表3 万全农田边缘土壤种子库各物种组成及其所占储量比例

图1 不同生境中刺萼龙葵土壤种子库的垂直分布
*表示同一生境不同土层在0.05水平差异显著。*indicates significant differences among different soil layers within the same habitat at 0.05 level.
2.3 两种生境中刺萼龙葵种子库季节动态变化
从图2可知, 河滩与农田边缘两个生境中刺萼龙葵种子库随着时间的推移逐渐变小。其中4月最多, 种子库分别平均为273粒∙m-2(河滩)和1 970粒∙m-2(农田边缘), 显著高于另外两次采样(<0.05); 6月分别为44粒∙m-2和472粒∙m-2, 8月分别为30粒∙m-2和158粒∙m-2。河滩生境中3个时间采集的刺萼龙葵种子量分别占土壤种子库总量的78.7%、12.7%和8.6%, 而在农田边缘生境中3个时间采集的刺萼龙葵种子占总量的比例分别为75.8%、18.2%和6.1%。

图2 不同生境中刺萼龙葵种子库季节动态变化
不同小写字母表示同一生境不同月份间在0.05水平差异显著。Different lowercase letters in the same habitat indicate significant differences among different months at 0.05 level.
2.4 替代控制对刺萼龙葵种子库的影响
由图3可知, 在替代控制第1年(2012年), 刺萼龙葵控制效果并不显著(>0.05), 除T5外, 其余各小区0~10 cm(图3d)土层中刺萼龙葵种子库储量均高于对照, 且刺萼龙葵种子大多存在于2~10 cm(图3b、3c),这可能是由于在试验开始前对替代控制区土壤进行翻动, 使得刺萼龙葵种子进入中下层。从替代控制第2年(2013年)开始, 各处理表层土壤(0~2 cm)中刺萼龙葵种子库储量均显著低于CK, 而中下层土壤(2~10 cm)中的刺萼龙葵种子储量与对照差异不显著(>0.05)。除2015年T8处理总种子库储量(0~10 cm)高于对照外, 其余处理均低于对照, 截至2017年, T6处理中刺萼龙葵种子库总储量最低, 为432 粒∙m-2, 显著低于对照种子库储量(<0.05), 控制效果明显。
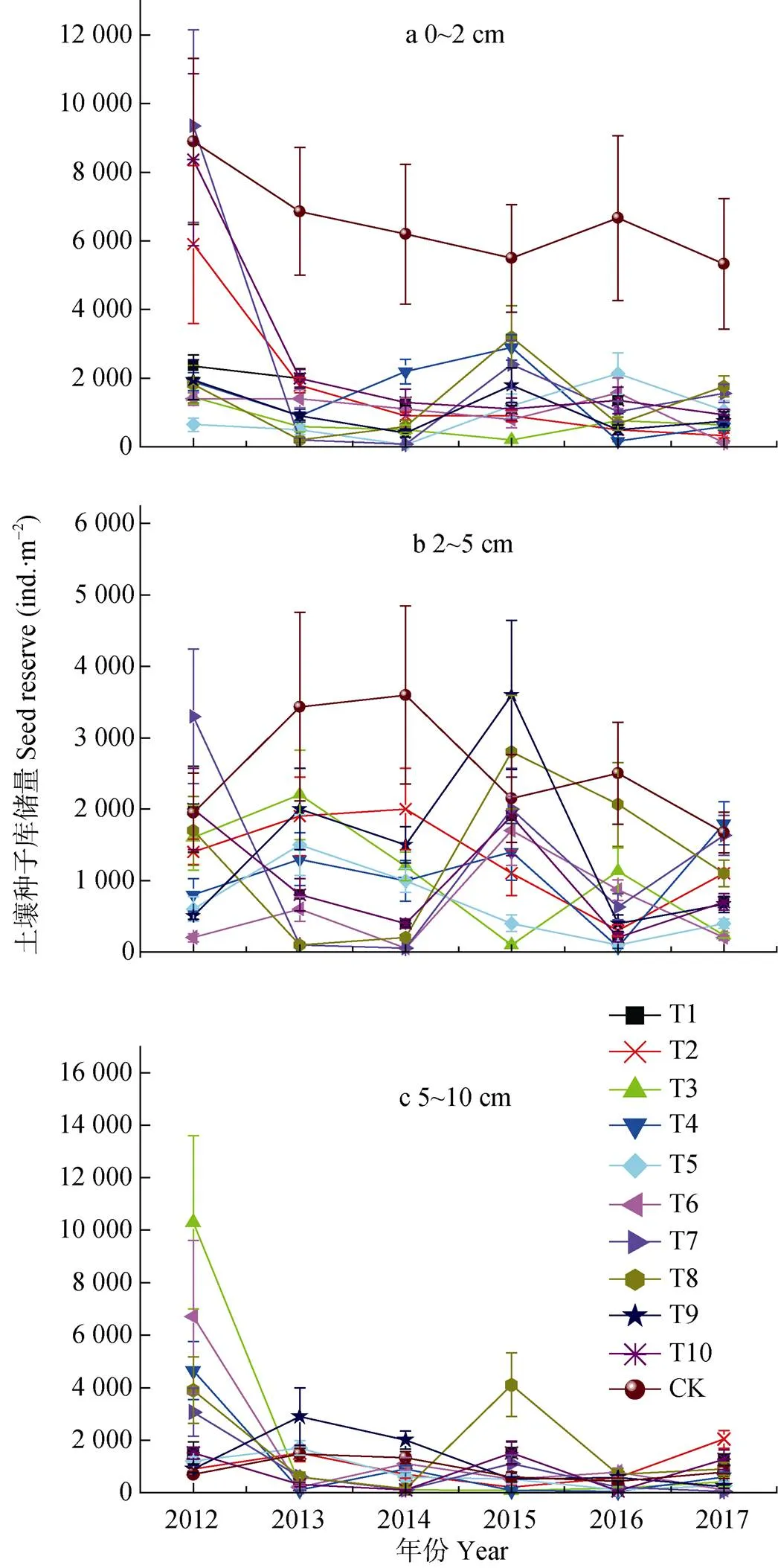
图3 不同植物替代控制处理对不同土层刺萼龙葵种子库动态变化的影响
对刺萼龙葵植株密度及生物量的调查结果表明, 在替代控制第1年(2012年), 各处理中刺萼龙葵密度即迅速下降(表4), 最低为18.0株∙m-2(T5), 较对照(199.5株∙m-2)下降91%(<0.05)。随着替代控制时间的延长, 各处理中牧草覆盖度增大, 与刺萼龙葵争夺资源, 挤占生态位, 使刺萼龙葵的生长受到显著限制, 截至2017年, 各小区中刺萼龙葵密度为4.0~16.8株∙m-2, 显著低于对照(96.0株∙m-2) (<0.05)。同样, 由表5可以看出, 刺萼龙葵的单株生物量从替代控制第1年开始即受到显著影响, 各小区中刺萼龙葵的单株生物量均显著低于同期对照(<0.05)。通过调查牧草产量可以看出(图4), 在2015年T1、T2、T4、T6、T10处理中牧草鲜重分别达14 207.1 kg∙hm-2、12 806.4 kg∙hm-2、12 206.1 kg∙hm-2、18 409.2 kg∙hm-2、15 007.5 kg∙hm-2, 干重也达4 000 kg∙hm-2以上, 其中T6处理在2017年鲜重达20 396.1 kg∙hm-2, 干重达7 710.6 kg∙hm-2。由此可见, 利用替代控制的方法防控刺萼龙葵, 经济效益可观。
通过各处理刺萼龙葵种子库储量、密度、生物量及牧草产量(鲜重)相关性分析发现(表6), 刺萼龙葵种子库与刺萼龙葵密度(<0.01)、生物量(<0.05)呈正相关, 而牧草产量与刺萼龙葵各指标均呈负相关, 但相关性不显著(>0.05)。以T6为例, 对不同年份土壤种子库与年降雨量进行回归分析(表7), 结果发现, 降雨量对刺萼龙葵种子库的影响较大, 从曲线拟合情况看, 幂函数和指数函数拟合效果好于线性函数和对数函数(<0.05), 从各回归模型的2值、值显著性综合分析, 刺萼龙葵种子库与降雨量之间的关系符合幂函数模型=2.6190.001。
3 讨论
3.1 刺萼龙葵种子库特征
种子库是杂草生命周期中重要组成部分, 是未来杂草种群的惟一来源, 因此, 了解种子库中物种组成变化对防除和控制杂草危害及生态修复尤为重要[26-27]。外来入侵植物影响了本地生物多样性和生态系统功能, 调查入侵植物土壤种子库以及入侵后本地植物种子库的组成和结构特征可以揭示植物入侵对生态系统的长期影响[28-29]。外来入侵杂草的发生与危害很大程度上依赖于土壤种子库储量的大小[30-31]。通过调查河滩和农田边缘两个刺萼龙葵入侵的典型生境中的土壤种子库特征, 发现农田边缘生境种子库储量大于河滩生境, 这可能是由于农田边缘生境中土壤较河滩更肥沃, 使得刺萼龙葵更容易生长。在农田边缘生境中刺萼龙葵种子库储量达2 600粒∙m-2, 因此, 即使把地上刺萼龙葵种群全部灭除, 土壤种子库这个潜在的种群仍可导致再度入侵; 另外, 河滩生境中另一种入侵植物少花蒺藜草种子库储量占总储量的36.70%, 这也增加了防除入侵杂草的难度。
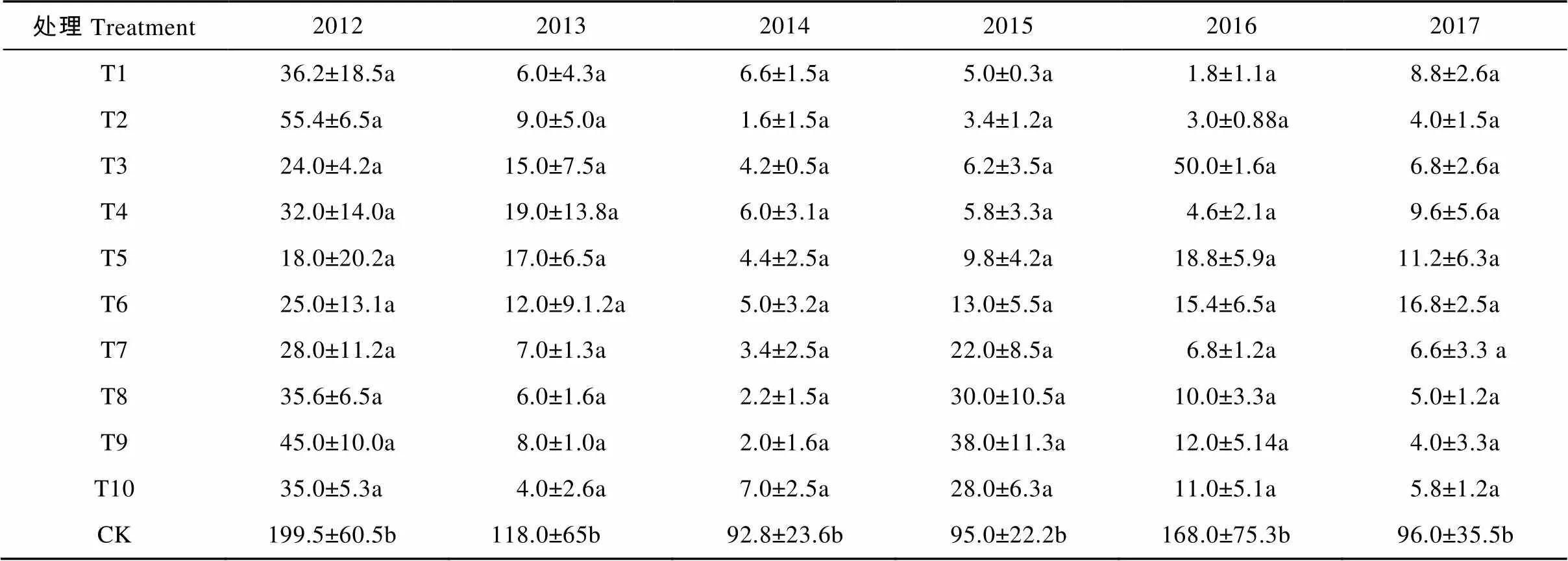
表4 2012—2017年连续不同植物替代控制处理对对刺萼龙葵密度的影响
同列不同小写字母表示不同植物替代处理间0.05水平差异显著。Different lowercase letters in the same column indicate significant differences among different plant replacement treatments at 0.05 level.

表5 2012—2017年连续不同植物替代控制处理对对刺萼龙葵生物量的影响
同列不同小写字母表示不同植物替代处理间0.05水平差异显著。Different lowercase letters in the same column indicate significant differences among different plant replacement treatments at 0.05 level.
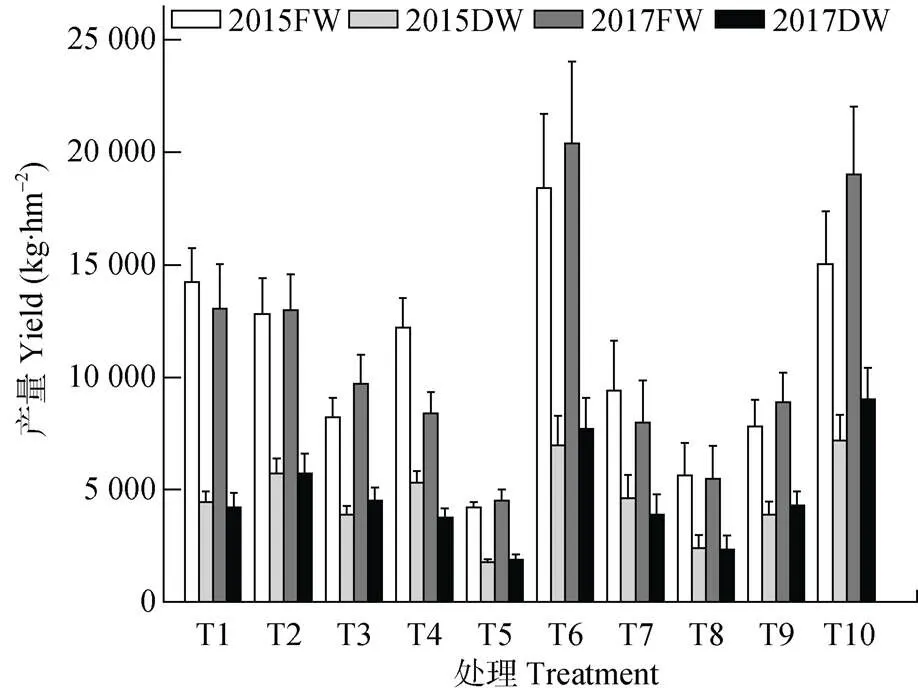
图4 替代控制试验过程中各处理替代牧草产量变化

表6 刺萼龙葵相关指标与牧草产量相关性分析
*和**分别表示在0.05和0.01水平显著相关。* and ** indicate significant correlation at 0.05 and 0.01 level, respectively.

表7 不同年份土壤种子库与年降雨量的回归分析——以T6处理为例
众多研究发现, 随土层加深, 种子储量逐渐变少[31-33]。在本研究中, 河滩生境中刺萼龙葵种子库体现了类似的规律, 刺萼龙葵种子主要分布在土壤上层(0~2 cm); 但河滩生境人为干扰较少, 大部分刺萼龙葵种子保留在中上层, 而农田边缘生境中翻耕、施肥、灌溉等农事操作频繁, 使更多的刺萼龙葵种子进入中下层(2~10 cm), 达到67.3%, 这与前人研究农田中入侵植物土壤种子库类似[30,34]; 当杂草种子进入种子库后, 种子可以感知种子库中的周围环境, 进入休眠状态或萌发, 人为扰动如农事操作、农作物管理等可以直接影响种子库中种子的环境, 可影响种子寿命及萌发行为[26]。因此, 要注意农田边缘生境土壤中层与下层(2~10 cm)中刺萼龙葵种子的潜在危害。
在我国刺萼龙葵发生区, 刺萼龙葵种子在4—5月气温达10 ℃时即开始萌发[35]。本研究也体现了类似的结果, 大部分刺萼龙葵种子存在于4月份土样中, 在两个生境中均达75%以上, 由于刺萼龙葵种子具有休眠特性[18], 因此, 两个生境中, 6月份与8月份采集的土样中仍分别有21.3%(河滩)和24.3%(农田边缘)的刺萼龙葵种子萌发。刺萼龙葵种子的休眠特性, 延长了其在入侵地的存留时间, 一旦环境适宜, 即可萌发, 完成生活史, 从而大大增加了防除难度。
3.2 替代控制措施对刺萼龙葵种子库的影响
减少杂草种子进入土壤种子库是有效控制杂草的最有效的措施[36-37]。目前能够有效控制刺萼龙葵种子产量的方法主要有物理防治与化学防治, 但物理防治需多年多次连续进行, 需要投入大量人力物力[38], 而化学防治虽然对刺萼龙葵的清除效果好, 见效快, 由于没有专一性强的除草剂, 特别是针对科尔沁草原等自然生态系统, 如果使用不当则很容易产生二次污染, 对当地环境造成破坏。在生态学原则的基础上, 除对原生态系统的保护外, 通过植物间竞争等手段, 采用植物替代控制和增加植物多样性是一种有效控制刺萼龙葵的方法[39-40]。
国内关于刺萼龙葵替代控制的研究较少, 且未对替代控制后的刺萼龙葵土壤种子库展开调查。由于刺萼龙葵种子具有休眠特性, 故不适合选择一年生替代植物控制刺萼龙葵; 针对发生在科尔沁草原的刺萼龙葵, 我们选取了竞争能力强、生物量大且生长迅速的多年生豆科牧草(紫花苜蓿和沙打旺)作为先锋植物, 选取多年生禾本科牧草(披碱草、无芒雀麦、苇状羊茅、冰草、紫羊茅和羊草)作为草场恢复植物, 通过设置一定的组合替代控制刺萼龙葵, 结果表明, 替代控制第1年, 替代植物生长较刺萼龙葵缓慢, 且刺萼龙葵密度大, 控制效果并未较好地体现, 但次年替代植物发芽早(如沙打旺、紫花苜蓿等), 生长迅速, 很快占据了生态位, 光照会一定程度抑制萼龙葵种子萌发[18], 且高郁闭度的替代植物挤占了刺萼龙葵的生长空间, 与其争夺光照、水分、养分等条件, 挤占刺萼龙葵的资源生态位, 有效地阻止了次年刺萼龙葵的生长, 进而表现为刺萼龙葵密度及生物量从第2年开始较对照显著降低, 有效减少了土壤中刺萼龙葵种子库储量。另外, 本文研究结果发现, 在2015年, 刺萼龙葵种子库储量、密度及生物量均有不同程度的升高, 除受替代控制影响外, 还与降雨量有关, 且这种关系可用幂函数进行拟合, 这也体现了生态控制的复杂性。
本文通过测定替代植物产量可以看出, T4和T7两个处理中替代植物产量在2017年有所下降, 这可能由于豆科牧草在生长3~7年后开始衰退造成的[41-42], 豆科植物作为先锋植物在荒漠治理[43]、尾矿修复[44-45]等方面已有大量报道, 故本研究部分小区选择豆科牧草与禾本科牧草作为组合, 将豆科牧草作为先锋植物在替代控制前期对刺萼龙葵起控制作用, 随着替代控制的进行及豆科牧草的衰退, 替代控制区逐渐恢复为禾本科植物为主的生态系统。另外, 本研究发现, 替代控制并未将刺萼龙葵种群完全清除, 其种子库储量及植株密度维持在一定水平, 因此, 若在科尔沁草原彻底清除刺萼龙葵, 应以替代控制为主, 辅以物理防除及化学防除, 逐步恢复生态系统健康。
4 结论
1)河滩及农田边缘刺萼龙葵发生的两个典型生境中刺萼龙葵种子库较大, 其中河滩生境中随着土壤深度的增加刺萼龙葵种子数量减少, 主要分布于中上层(0~5 cm), 农田边缘生境3层种子数量差异不大, 但需注意中下层(2~10 cm)种子潜在危害; 两种生境中刺萼龙葵种子库大小随着时间的推移逐渐变小。
2)本文选用豆科与禾本科牧草作为替代植物, 可将刺萼龙葵种子库储量及密度控制在较低水平, 沙打旺+苇状羊茅+冰草+羊草组合对刺萼龙葵控制效果最佳, 同时又可获得可观的经济效益; 替代控制受环境(如降雨)影响较大, 因此, 可辅以物理、化学防控等手段形成刺萼龙葵综合防控体系, 对刺萼龙葵进行根除, 可用于控制草场、农田周边、荒地刺萼龙葵的发生与蔓延。
[1] KEELEY J E. Seed production, seed populations in soil, and seedling production after fire for two congeneric pairs of sprouting and nonsprouting Chaparal shrubs[J]. Ecology, 1977, 58(4): 820-829
[2] ROBERTS H A. Seed banks in the soil[J]. Advance in Applied Biology, 1981, 6: 1-55
[3] 张志权. 土壤种子库[J]. 生态学杂志, 1996, 15(6): 36-42 ZHANG Z Q. Soil seed bank[J]. Chinese Journal of Ecology, 1996, 15(6): 36-42
[4] 徐高峰, 申时才, 张付斗, 等. 外来入侵植物小子虉草研究进展与展望[J]. 中国生态农业学报, 2015, 23(9): 1083-1092 XU G F, SHEN S C, ZHANG F D, et al. Research progress and prospect about exotic invasive speciesRetz[J]. Chinese Journal of Eco-Agriculture, 2015, 23(9): 1083-1092
[5] MARQUES A R, COSTA C F, ATMAN A P F, et al. Germination characteristics and seedbank of the alien species(Fabaceae) in Brazilian forest: Ecological implications[J]. Weed Research, 2014, 54(6): 576-583
[6] 白文娟, 章家恩, 全国明. 土壤种子库研究的热点问题及发展趋向[J]. 土壤, 2012, 44(4): 562-569 BAI W J, ZHANG J E, QUAN G M. Hot topics and developing trends in soil seed bank[J]. Soils, 2012, 44(4): 562-569
[7] STREHLOW T, DEKEYSER S, KOBIELA B. Managing seedbank composition to enhance wetland restoration[J]. Ecological Restoration, 2017, 35(1): 12-16
[8] SINGH R P, BAGNALL R H.Dunal., a new test plant for the potato spindle tuber virus[J]. American Potato Journal, 1968, 45(9): 335-336
[9] 关广清, 高东昌. 又有五种杂草传入我国[J]. 植物检疫, 1982, 6: 2 GUAN G Q, GAO D C. Five other weeds spread to China[J]. Plant Quarantine, 1982, 6: 2
[10] 王维升, 郑红旗, 朱殿敏, 等. 有害杂草刺萼龙葵的调查[J]. 植物检疫, 2005, 19(4): 247-248 WANG W S, ZHENG H Q, ZHU D M, et al. Investigation of harmful weedDunal.[J]. Plant Quarantine, 2005, 19(4): 247-248
[11] 车晋滇, 刘全儒, 胡彬. 外来入侵杂草刺萼龙葵[J]. 杂草学报, 2006, (3): 58-60 CHE J D, LIU Q R, HU B. Alien invasive weed —Dunal.[J]. Weed Science, 2006, (3): 58-60
[12] 贺俊英, 哈斯巴根, 孟根其其格, 等. 内蒙古新外来入侵植物——黄花刺茄(Dunal.)[J]. 内蒙古师范大学学报: 自然科学汉文版, 2011, 40(3): 288-290 HE J Y, KHASBAGAN, MONGENQIQIG, et al.Dunal.: A newly invaded alien plant of Inner Mongolia[J]. Journal of Inner Mongolia Normal University: National Science Edition, 2011, 40(3): 288-290
[13] 向俊, 李翠妮, 刘全儒, 等. 北京外来入侵植物刺萼龙葵的生态状况[J]. 生态学杂志, 2011, 30(3): 453-458 XIANG J, LI C N, LIU Q R, et al. Ecological state of invasive alien plantin Beijing[J]. Chinese Journal of Ecology, 2011, 30(3): 453-458
[14] BAH M, GUTIÉRREZ D M, ESCOBEDO C, et al. Methylprotodioscin from the Mexican medical plant(Solanaceae)[J]. Biochemical Systematics and Ecology, 2004, 32(2): 197-202
[15] 张少逸, 魏守辉, 张朝贤, 等. 刺萼龙葵种子休眠和萌发特性研究进展[J]. 杂草学报, 2011, 29(2): 5-9 ZHANG S Y, WEI S H, ZHANG C X, et al. Research advances on seed dormancy and germination of buffalobur ()[J]. Weed Science, 2011, 29(2): 5-9
[16] 苏秋霞, 李青丰. 黄花刺茄种子休眠及发芽特性[J]. 草业科学, 2014, 31(7): 1298-1301 SU Q X, LI Q F. Seed dormancy and germination of[J]. Pratacultural Science, 2014, 31(7): 1298-1301
[17] 钟艮平, 沈文君, 万方浩, 等. 用GARP生态位模型预测刺萼龙葵在中国的潜在分布区[J]. 生态学杂志, 2009, 28(1): 162-166 ZHONG G P, SHEN W J, WAN F H, et al. Potential distribution areas ofin China: A prediction with GARP niche model[J]. Chinese Journal of Ecology, 2009, 28(1): 162-166
[18] WEI S H, ZHANG C X, LI X J, et al. Factors affecting buffalobur () seed germination and seedling emergence[J]. Weed Science, 2009, 57(5): 521-525
[19] 邵华. 外来入侵植物刺萼龙葵的化感作用研究[J]. 种子, 2015, 34(8): 101-104 SHAO H. Study on the allelopathic effect ofDunal[J]. Seed, 2015, 34(8): 101-104
[20] 赵晓红, 张国良, 张瑞海, 等. 刺萼龙葵入侵对不同生境土壤特征的影响[J]. 生态环境学报, 2017, 26(6): 924-930 ZHAO X H, ZHANG G L, ZHANG R H, et al. Effects ofinvasion on soil characteristics in different habitats[J]. Ecology and Environment Sciences, 2017, 26(6): 924-930
[21] HUANG H J, LING T J, WANG H M, et al. One new flavonoid from[J]. Natural Product Research, 2017, 31(5): 1831-1835
[22] CHANG L, SHAO Q, XI X J, et al. Separation of four flavonol glycosides fromDunal using aqueous two-phase flotation followed by preparative high-performance liquid chromatography[J]. Journal of Separation Science, 2017, 40(3): 804-812
[23] ZHAO J L, LOU A R. Genetic diversity and population structure of the invasive plantin China[J]. Russian Journal of Ecology, 2017, 48(2): 134-142
[24] 张少逸, 张朝贤, 杨连喜, 等. 茎叶除草剂对刺萼龙葵的防治效果评价[J]. 植物保护, 2012, 38(5): 170-173 ZHANG S Y, ZHANG C X, YANG L X, et al. Efficacy evaluation of foliar-applied herbicides on[J]. Plant Protection, 2012, 38(5): 170-173
[25] 农业部环境保护科研监测所. 恶性杂草刺萼龙葵入侵天然植被快速恢复方法: 中国, CN201210355171[P]. 2013-01-09 Agro-Environmental Protection Institute, Ministry of Agriculture. Method for quickly recovering natural vegetation invaded by malignant weed: CN, CN201210355171[P]. 2013-01-09
[26] GULDEN R H, SHIRTLIFFE S J. Weed seed banks: Biology and management[J]. Weeds, Herbicides and Management, 2009, 2: 46-52
[27] 张广帅, 邓浩俊, 杜锟, 等. 汶川地震生态治理区土壤种子库及其与地上植被的关系[J]. 中国生态农业学报, 2015, 23(1): 69-79 ZHANG G S, DENG H J, DU K, et al. Soil seed bank and its correlations with aboveground vegetation in ecological restoration zones of Wenchuan Earthquake Region[J]. Chinese Journal of Eco-Agriculture, 2015, 23(1): 69-79
[28] MOSTERT E, GAERTNER M, HOLMES P M, et al. Impacts of invasive alien trees on threatened lowland vegetation types in the Cape Floristic Region, South Africa[J]. South African Journal of Botany, 2017, 108: 209-222
[29] RUSTERHOLZ H P, KÜNG J, BAUR B. Experimental evidence for a delayed response of the above-ground vegetation and the seed bank to the invasion of an annual exotic plant in deciduous forests[J]. Basic and Applied Ecology, 2017, 20: 19-30
[30] 张衍雷, 张瑞海, 付卫东, 等. 不同农作措施对少花蒺藜草(Benth)种子库及其繁殖能力的影响[J]. 农业资源与环境学报, 2015, 32(3): 312-320 ZHANG Y L, ZHANG R H, FU W D, et al. Effects of Different cultivation practices on the amount of seeds in the soils and seed production ofBenth[J]. Journal of Agricultural Resources and Environment, 2015, 32(3): 312-320
[31] 张瑞海, 付卫东, 宋振, 等. 河北地区黄顶菊土壤种子库特征及其对替代控制的响应[J]. 生态环境学报, 2016, 25(5): 775-782ZHANG R H, FU W D, SONG Z, et al. Characteristics of Soil seed bank ofand its response to control with replacement plants in Hebei, China[J]. Ecology and Environmental Sciences, 2016, 25(5): 775-782
[32] 张志明, 沈蕊, 张建利, 等. 元江流域干热河谷灌草丛土壤种子库与地上植物群落的物种组成比较[J]. 生物多样性, 2016, 24(4): 431-439 ZHANG Z M, SHEN R, ZHANG J L, et al. Comparisons of species composition between soil seed banks and aboveground plant communities in the dry-hot valley of the Yuanjiang River[J]. Biodiversity Science, 2016, 24(4): 431-439
[33] RUWANZA S. Soil seed bank depletion as a mechanism ofL. invasion[J]. South African Journal of Plant and Soil, 2016, 33(4): 303-308
[34] 章超斌, 马波, 强胜. 江苏省主要农田杂草种子库物种组成和多样性及其与环境因子的相关性分析[J]. 植物资源与环境学报, 2012, 21(1): 1-13 ZHANG C B, MA B, QIANG S. Analyses of species composition and diversity of weed seed bank of main crop fields in Jiangsu Province and its correlation with environmental factors[J]. Journal of Plant Resources and Environment, 2012, 21(1): 1-13
[35] 林玉, 谭敦炎. 一种潜在的外来入侵植物: 黄花刺茄[J]. 植物分类学报, 2007, 45(5): 675-685LIN Y, TAN D Y. The potential and exotic invasive plant:[J]. Acta Phytotaxonomica Sinica, 2007, 45(5): 675-685
[36] REID A M, MORIN L, DOWNEY P O, et al. Does invasive plant management aid the restoration of natural ecosystems?[J]. Biological Conservation, 2009, 142(10): 2342-2349
[37] GIORIA M, OSBORNE B. Similarities in the impact of three large invasive plant species on soil seed bank communities[J]. Biological Invasions, 2010, 12(6): 1671-1683
[38] 庞立东, 孙余卓. 刺萼龙葵的入侵机理与控制策略研究进展[J]. 中国植保导刊, 2016, 36(8): 20-25 PANG L D, SUN Y Z. Research progress on invasive mechanism ofDunal. and its control strategy[J]. China Plant Protection, 2016, 36(8): 20-25
[39] SHELEY R L, KRUEGER-MANGOLD J. Principles for restoring invasive plant-infested rangeland[J]. Weed Science, 2003, 51(2): 260-265
[40] WAN F H, LIU W X, GUO J Y, et al. Invasive mechanism and control strategy of(Sprengel)[J]. Science China Life Sciences, 2010, 53(11): 1291-1298
[41] 孙启忠, 桂荣, 那日苏, 等. 赤峰地区不同生长年限沙打旺生产力的研究[J]. 中国草地学报, 1999, (5): 30-35SUN Q Z, GUI R, NA R S, et al. Study on productivity ofPall. with different growth periods[J]. Chinese Journal of Grassland, 1999, (5): 30-35
[42] 陈志怡, 李金月. 不同生长年限对紫花苜蓿产量及土壤养分的影响[J]. 山东农业大学学报: 自然科学版, 2015, 46(2): 214-220 CHEN Z Y, LI J Y. Effects of growth years on the yield ofL. and soil nutrient[J]. Journal of Shandong Agricultural University: Natural Science Edition, 2015, 46(2): 214-220
[43] ZHANG B, GUI D W, GAO X P, et al. Controlling soil factor in plant growth and salt tolerance of leguminous plantShap. in saline deserts, Northwest China[J]. Contemporary Problems of Ecology, 2018, 11(1): 111-121
[44] 慈恩, 高明. 环境因子对豆科共生固氮影响的研究进展[J]. 西北植物学报, 2005, 25(6): 1269-1274CI E, GAO M. Research advances in the effects of environmental factors on the symbiotic nitrogen fixation of legumes[J]. Acta Botanica Boreali-Occidentalia Sinica, 2005, 25(6): 1269-1274
[45] 赵俊峰, 肖礼, 安韶山, 等. 永利煤矿复垦区植物叶片和枯落物生态化学计量学特征[J]. 生态学报, 2017, 37(9): 3036-3045ZHAO J F, XIAO L, AN S S, et al. Ecological stoichiometry characteristics of leaves and litter in plant communities in the Yongli colliery reclamation area[J]. Acta Ecologica Sinica, 2017, 37(9): 3036-3045
Characteristics of soil seed bank forDunal.and its response to control with replaceable plants*
ZHANG Ruihai1, SONG Zhen1, ZHANG Guoliang1, WANG Zhonghui1, FU Weidong1**, YIN Shuhong2, ZHAI Hongkai2, XI Kunpeng2
(1. Institute of Agricultural Environment and Sustainable Development, Chinese Academy of Agricultural Sciences, Beijing 100081, China; 2. Agricultural Environmental Protection and Rural Energy Management Station of Baicheng, Baicheng 137000, China)
Soil seed bank is important in the restoration and succession of vegetation in degraded ecosystems. In order to determine the characteristics of soil seed bank of the alien invasive plantDunal. and to evaluate the effects of replacement control of, soil seed bank ofin two typical habitats (riverside and farmland margin) were studied and its’ vertical distribution and seasonal dynamics analyzed. In addition, a study on replacement control ofwas conducted using varieties of perennial forages of Gramineae and Leguminosea. 1) Based on the results, 23 plants species were identified, and Gramineae and Compositae were the most dominant families. The total reserve ofseed banks in the riverside and farmland margin were respectively 347 seeds·m-2and 2 600 seeds·m-2, which accounted for 2.46% and 35.16% of the whole seed bank.was the dominant family in farmland but another invasive plant (Cav.) was the main species in riverside; reaching 5 187 seeds∙m-2and accounting for 36.70% of the seed reserve. 2)seeds were mainly stored in the 0–2 cm soil layer in riverside region, accounting for 64.3% and decreasing with increasing soil depth. There was no significant difference in the three soil layers of farmland margin in terms ofseed reserve (> 0.05), which accounted for 32.7% (0-2 cm), 38.2% (2-5 cm) and 29.1% (5-10 cm) of the total seed bank. However, it was need to pay attention to potential hazards of seeds in the middle and lower layers (2-10 cm). 3) For the three times of samplings,seeds mainly were collected in April reached 273 seeds∙m-2and 1 970 seeds∙m-2in riverside and farmland margin, significantly fewer in June and August. 4) In the second year of the controlled replacement, the coverage of forage gradually increased, the resources and niches were seized by the forage, and the growth ofsignificantly inhibited. The density, biomass and soil seed reserve ofwere controlled at a low level, significant lower than CK (< 0.05).Pall.,Schreb.,(L.) Gaertn., combined with(Trin.) Tzvel. had the best control effect on. At the same time, it showed obvious economic benefits, which produced forage grass of 20 396.1 kg·hm-2(fresh weight) and 7 710.6 kg·hm-2(dry weight). 5) Soil seed reserves ofwas positively correlation with density (< 0.01) and biomass (< 0.05) of, while forage yield was negatively correlation with soil seed reserve, density and biomass ofHowever, this was not significant (> 0.05). Nevertheless, rainfall significantly affected soil seed reserve of(< 0.05), which was described by the power function model= 2.6190.001(20.822,= 18.486,= 0.013). Therefore, plant replacement combined with physical and chemical methods can be used to establish integrated system to controlgrowth in habitats such as grassland, farmland margin and wasteland.
Biological invasion;Dunal.; Invasive plant; Soil seed bank; Temporal and spatial dynamics; Replacement control
, E-mail: fuweidong@caas.cn
Jul. 26, 2018;
Oct. 19, 2018
Q948
A
2096-6237(2019)03-0337-11
10.13930/j.cnki.cjea.180697
张瑞海, 宋振, 张国良, 王忠辉, 付卫东, 尹树红, 翟洪凯, 席坤鹏. 刺萼龙葵土壤种子库特征及其对替代控制的响应[J]. 中国生态农业学报(中英文), 2019, 27(3): 337-347
ZHANG R H, SONG Z, ZHANG G L, WANG Z H, FU W D, YIN S H, ZHAI H K, XI K P. Characteristics of soil seed bank forDunal. and its response to control with replaceable plants[J]. Chinese Journal of Eco-Agriculture, 2019, 27(3): 337-347
* 国家重点研发计划项目(2016YFC1201203)资助
付卫东, 主要研究方向为外来入侵植物入侵机制与防控。E-mail: fuweidong@caas.cn
张瑞海, 主要研究方向为入侵植物防控。E-mail: ruihai.321@163.com
2018-07-26
2018-10-19
* This study was supported by the National Key R&D Project of China (2016YFC1201203).
