香蕉-甘蔗轮作模式防控香蕉枯萎病的持续效果与土壤微生态机理(Ⅱ)*
2019-03-08林威鹏曾莉莎伍朝荣周建坤刘文清蔡昆争杜彩娴
林威鹏, 曾莉莎, 吕 顺**, 伍朝荣, 王 芳, 周建坤, 刘文清, 蔡昆争, 杜彩娴, 夏 玲
香蕉-甘蔗轮作模式防控香蕉枯萎病的持续效果与土壤微生态机理(Ⅱ)*
林威鹏1,2,3†, 曾莉莎1†, 吕 顺1**, 伍朝荣2, 王 芳1, 周建坤1, 刘文清1, 蔡昆争2, 杜彩娴1, 夏 玲1
(1. 东莞市香蕉蔬菜研究所 东莞 523061; 2. 华南农业大学资源环境学院 广州 510642; 3. 广东省农业科学院茶叶研究所 广州 510642)
香蕉枯萎病是由尖孢镰刀菌古巴专化型[f. sp.]侵染引起维管束坏死的一种毁灭性真菌病害, 是世界范围内分布最广、毁灭性最强的植物病害之一。我们在古籍《广东新语》找到有关香蕉-甘蔗轮作的记载, 经大田试验证明, 连作蕉地轮作甘蔗2年后, 后茬香蕉枯萎病发病率大幅下降至1.79%。在本研究中, 我们试图探明连作蕉地轮作甘蔗, 后茬香蕉的持续抑病效果及其机理。连作蕉地轮作甘蔗2年后, 测定回种不同年限香蕉枯萎病发病率和土壤可培养微生物数量, 并通过高通量测序技术分析土壤细菌微生物群落结构和组成的变化。结果表明: 连作蕉地轮作甘蔗2年后, 甘蔗地后茬香蕉枯萎病发病率仅为1.79%, 后续3年发病率逐年递增, 分别为21.93%、25.80%和28.81%, 但均低于多年连作蕉地的平均发病率(49.15%)。土壤放线菌和尖孢镰刀菌总数随香蕉种植年限增加而显著增加。高通量测序结果表明, 酸杆菌门(32.86%)、变形菌门(28.85%)和绿弯菌门(12.33%)3个菌门为香蕉-甘蔗轮作系统香蕉根围土细菌的优势菌门。酸杆菌目(Acidobacteriales)细菌相对丰度随回种香蕉逐年增加, 并与香蕉枯萎病发病率呈显著正相关的关系; 而假单胞菌目(Pseudomonadales)、浮霉菌目(Planctomycetales)、球杆菌目(Sphaerobacterales)、乳酸杆菌目(Lactobacillales)和土壤红杆菌目(Solirubrobacterales)细菌随香蕉回种年限的增加呈下降趋势, 并与香蕉枯萎病发病率呈显著负相关的关系。连作蕉地轮作甘蔗2年后, 4年内仍能保持较好的抑病效果。本模式具有轮作时间短、抑病效果好、持续时间长、经济效益高等特点。
香蕉-甘蔗轮作; 香蕉枯萎病; 尖孢镰刀菌; 持续效果; 控病机理
香蕉枯萎病是由尖孢镰刀菌古巴专化型f. sp(FOC)侵染引起的维管束类疾病(又称香蕉黄叶病、巴拿马枯萎病), FOC在土壤中可存活15年以上, 生存和传播能力极强[1]。由于气候条件的限制, 我国可种植香蕉(spp.)的耕地面积有限, 加之该病害在我国的迅速蔓延, 已严重阻碍现代香蕉产业的可持续发展[2-4]。
轮作、间作、套作等种植模式是我国传统农业主要种植方式, 在一定程度上能有效防控作物土传病害的发生。前人研究表明, 轮作能减少土传病害的发生[5-6], 促进土壤养分的释放[7-8], 改善根际微生物环境[9-10]。对黄瓜()、当归()、马铃薯()等作物的研究表明, 轮作均能有效缓解连作障碍[11-13]。已有研究表明, 连作香蕉地轮作水稻()[14]、木薯()[15]、玉米()[5]、菠萝[5]()、韭菜()[16]均能不同程度降低后茬香蕉枯萎病的发病率。因此, 合适的轮作模式可以降低香蕉枯萎病的发病率, 最终获得香蕉种植的最大经济效益。目前与香蕉进行轮作的作物, 经济效益远低于香蕉。寻找一种轮作周期短、抑病效果好、持续时间长、经济效益高、前后茬操作模式相近的轮作模式对当前解决香蕉连作障碍具有重要的现实需求与经济意义。清代屈大均所著文献《广东新语》(成书于康熙17年左右)记载的香蕉-甘蔗()轮作模式为香蕉轮作甘蔗2年再复种香蕉3~4年, 周而复始[17]。我们的同期研究表明[18], 连作蕉地分别轮作甘蔗1年、2年和3年后, 下茬香蕉枯萎病的发病率分别为17.86%、1.79%和1.79%, 即香蕉轮作甘蔗2年即可达到较优抑病效果。承接前文的研究, 本试验试图探明甘蔗轮作2年后茬的持续抑病效果。通过测定土壤可培养微生物数量, 以及通过高通量测序技术揭示土壤微生物群落结构的变化, 探明其中的原理及香蕉-甘蔗轮作模式抑制后茬香蕉枯萎病发病的可能机制, 为建立合理的香蕉轮作体系提供科学的理论依据。
1 材料与方法
1.1 试验设计
田间试验位于广东省东莞市沙田镇中围村, 面积1.4 hm2, 为同一农户耕种同一幅地块平坦的多年连作香蕉地。土质为轻黏壤土, 土层深厚, 土质疏松, 有机质丰富。起始时土壤耕层含有机质12.30 g∙kg-1、全氮164.9 mg∙kg-1, pH 5.2。2003年个别植株出现香蕉枯萎病, 至2007年枯萎病发病率达80%以上, 2008年开始改种中等抗病品种‘农科1号’香蕉, 当年‘农科1号’枯萎病发病率达30%以上, 2009年开始进行香蕉-甘蔗轮作试验, 样品测定在东莞市香蕉蔬菜研究所及华南农业大学进行。
试验共设4个处理(表1), 每个处理3个重复, 每个重复250 m2, 种植42棵香蕉。CK: 轮作甘蔗2年处理(连作香蕉多年—轮作甘蔗2年—种植香蕉0年); XJ1: 轮作后回种香蕉1年处理(香蕉连作多年—轮作甘蔗2年—种植香蕉1年); XJ2: 轮作后回种香蕉2年处理(香蕉连作多年—轮作甘蔗2年—种植香蕉2年); XJ3: 轮作后回种香蕉3年处理(香蕉连作多年—轮作甘蔗2年—种植香蕉3年)。
轮作后种植的香蕉品种仍为‘农科1号’(AAA Cavendish, 广州市农业科学研究院选育), 采用高畦深沟, 双行春植的栽培方式。每公顷种植1 650株, 行距为2.60 m, 株距为2.40 m; 畦面宽4.5 m, 畦间沟宽0.5 m、深0.7 m。采用常规方法进行种植管理。种植的甘蔗品种为‘广东黄皮果蔗’, 每米植蔗沟约种12个芽, 每公顷有效茎约75 000条。种茎以品字形或双轨窄幅排放, 两行种茎之间距离10 cm左右, 种茎与土壤紧贴, 芽排向两侧。种前施足基肥, 采用常规方法进行种植管理。
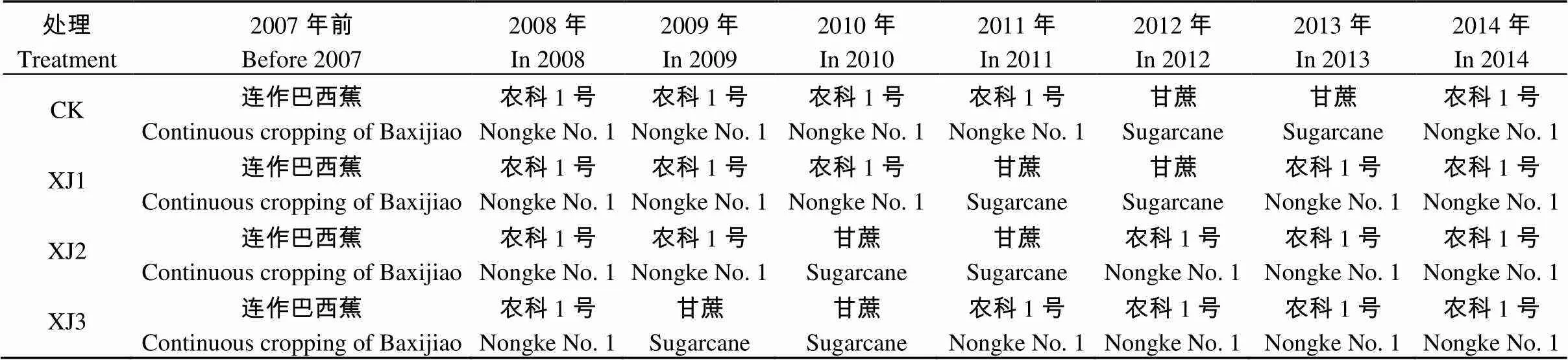
表1 不同处理2007—2014年香蕉和甘蔗的轮作模式
‘巴西蕉’和‘农科1号’为香蕉品种。‘Baxijiao’ and ‘Nongke No. 1’ are varieties of banana.
1.2 土壤样品的采集
在香蕉抽蕾挂果阶段(甘蔗收获期)的枯萎病发生高峰期, 于2013年10月14日统一采集土壤样本, 用于测定轮作甘蔗2年后回种不同年限香蕉[0年(CK)、1年(XJ1)、2年(XJ2)、3年(XJ3)]土壤可培养微生物及土壤微生物群落结构。于试验地每个小区内采用5点取样法取样, 去掉0~5 cm的表土, 采距植株20~30 cm的根围土壤, 深约20 cm, 每个处理采集3个重复小区共12个土壤样品, 将所采每个小区的土样混匀后分成2份, 一份于-20 ℃冷冻保存用于土壤DNA提取, 一份用于测定土壤中可培养微生物数量。
1.3 测定指标与方法
1.3.1 发病率及相对防效
2014年11月3日(采集土壤1年以后), 在香蕉抽蕾期跟踪调查香蕉枯萎病的发病情况, 计算各处理的香蕉枯萎病发病率及相对防效。
枯萎病发病率(%)=发病植株数/供试植株总数×100 (1)
相对防效(%)=(连作蕉地的发病率-处理的发病率)/连作蕉地的发病率×100 (2)
1.3.2 可培养微生物
土壤微生物的测定参照林先贵[19]方法, 采用平板培养法测定土壤中的微生物数量。细菌测定用牛肉汁蛋白胨琼脂, 真菌用马丁氏琼脂, 放线菌用高氏1号琼脂, 尖孢镰刀菌采用Komada改良培养基[20-21]。
1.4 土壤细菌群落结构的检测方法及测序流程
1.4.1 土壤DNA的提取
称取0.5 g新鲜土样置于Fast DNA cube中, 使用FastDNA土壤DNA提取试剂盒提取土壤DNA。提取的DNA使用微量紫外分光光度计测定浓度, 用0.8%琼脂糖凝胶电泳检测样品DNA片段大小。
1.4.2 基因扩增与纯化
选用引物27F(5¢-AGAGTTTGATCCTGGCTCAG-3¢)和533R(5¢-TTACCGCGGCTGCTGGCAC-3¢)[22]对原核生物16s片段的V1-V3片区进行扩增, 产物进行纯化、定量和均一化形成测序文库, 建好的文库先进行文库质检, 质检合格的文库用Roche 454 FLX+系统进行测序。
1.4.3 生物信息学分析
对测序得到的原始数据进行拼接、过滤, 得到有效序列和优质序列[23-24]。基于优质序列按照97%相似度进行OTUs(operational taxonomic units)聚类和物种分类分析[25], 并将OTUs和物种注释结合, 从而得到每个样品的OTUs和分类谱系的基本分析结果。再对OTUs进行丰度、多样性指数等分析[26-27], 同时对物种注释在各个分类水平上进行群落结构的统计分析。由上海派森诺生物科技有限公司完成测序及初步分析, 广州基迪奥生物科技有限公司完成进一步分析。
1.5 数据分析
采用Microsoft Excel 2016进行数据的整理, 使用SPSS 21.0进行数据的统计分析, 统计前对数据进行正态性检验和组件方差齐性分析, 如果无法满足以上两个条件, 则对数据进行log10转化, 然后对数据进行方差分析(one-way ANOVA),采用最小显著极差法(LSD法)进行数据的多重比较。
2 结果与分析
2.1 香蕉地轮作甘蔗2年对后茬香蕉枯萎病发病率的影响
连作香蕉地轮作甘蔗2年后, 回种不同年限香蕉, 后茬香蕉枯萎病发病率如图1所示。根据同期研究[18],连作蕉地枯萎病平均发病率为49.15%。轮作2年甘蔗后, 甘蔗地(CK)的后茬香蕉枯萎病发病率为1.79%, 回种香蕉1年(XJ1)的后茬香蕉发病率为21.93%, 回种香蕉2年(XJ2)的后茬香蕉发病率为25.80%, 回种香蕉3年(XJ3)的后茬香蕉发病率为28.81%。与香蕉连作地相比, 轮作甘蔗两年后, 甘蔗地CK的相对防效高达96.36%, XJ1、XJ2、XJ3的相对防效分别为55.38%、47.50%和41.38%。以上结果表明连作蕉地轮作甘蔗2年后, 回种香蕉4年内对香蕉枯萎病均具有一定的防控效果, 但防控效果随着回种香蕉年数的增加而逐步降低, 但仍低于连作蕉地的平均发病率。
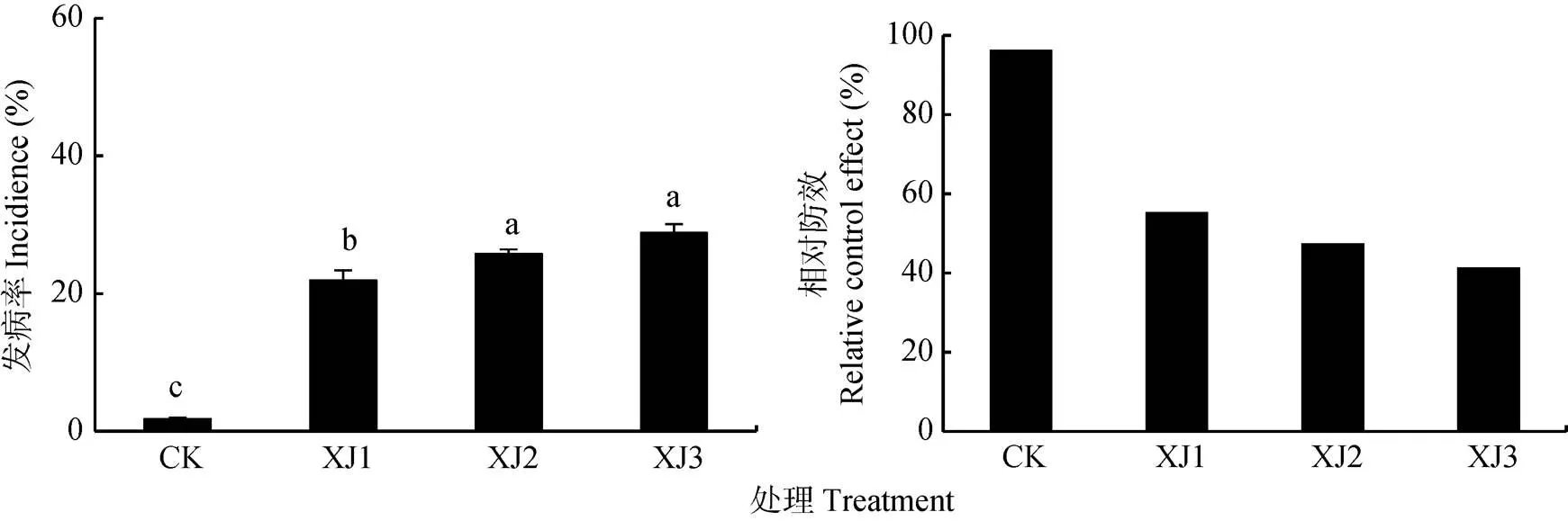
图1 连作香蕉地轮作2年甘蔗后茬香蕉枯萎病发病率和相对防效
不同字母表示处理间差异显著(<0.05)。Different letters indicate significant differences among treatments at 0.05 level.
2.2 香蕉地轮作甘蔗2年复种香蕉对土壤可培养微生物的影响
连作香蕉地轮作2年甘蔗后, 回种不同年限香蕉, 土壤可培养微生物数量如图2所示。与甘蔗地CK相比, 回种香蕉后土壤微生物总数增加, 其中土壤细菌数量在回种香蕉1年(XJ1)有所提高, 回种2年(XJ2)显著提高, 回种3年(XJ3)开始回落。回种香蕉1年, 真菌数量并没有显著变化, 回种2年开始显著增加, 回种3年回落。放线菌在回种香蕉1和2年无显著提高, 回种3年显著提高, 并随回种香蕉年数的增加而显著提高。尖孢镰刀菌随着回种香蕉年数的增加而逐年增加。
2.3 香蕉地轮作甘蔗2年复种香蕉对土壤细菌群落结构的影响
2.3.1 微生物多样性
轮作2年甘蔗后回种不同年限香蕉, 土壤微生物多样性如表2所示。结果表明, 土壤原核微生物群落丰度指数(Chao1、ACE)随香蕉种植年数的增加而显著提高,群落多样性Shannon指数也随香蕉种植年数的增加而显著提高。即与甘蔗地(CK)相比, 连作香蕉地轮作2年甘蔗后, 回种香蕉时间越长, 土壤原核微生物群落的丰度和多样性越高。
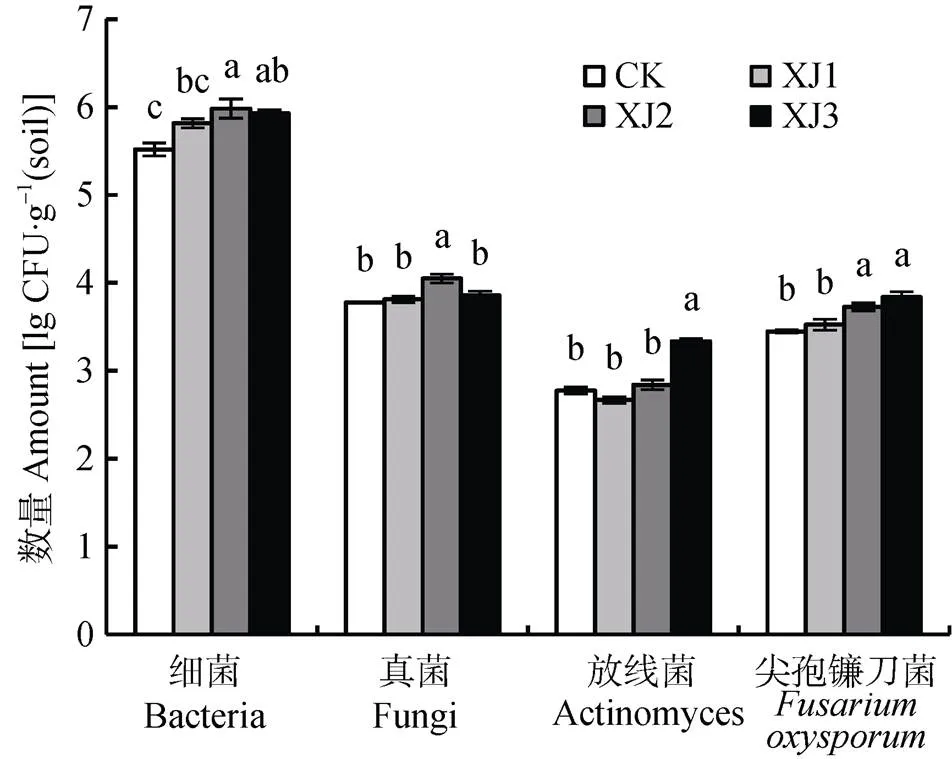
图2 连作香蕉地轮作2年甘蔗后茬土壤可培养微生物数量
不同字母表示处理间差异显著(<0.05)。Different letters indicate significant differences among treatments at 0.05 level.
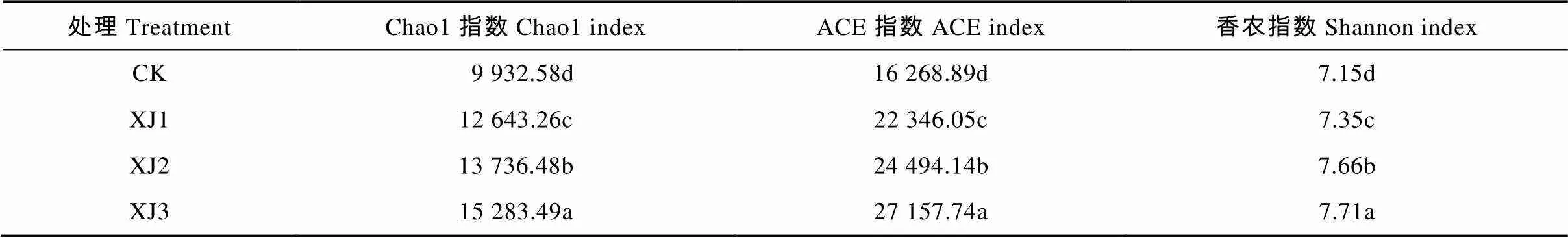
表2 连作香蕉地轮作2年甘蔗后种植不同年限香蕉土壤细菌多样性
同列不同字母表示处理间差异显著(<0.05)。Different letters in the same column indicate significant differences among treatments at 0.05 level.
2.3.2 样品间OTU聚类
根据OTU的表达谱数据, 本研究对回种不同年限香蕉土壤细菌构成进行分析(以OTU包含tags数量达总tags数量的0.1%作为阈值), 结果如图3所示。轮作2年甘蔗后回种香蕉1年(XJ1)土壤细菌构成与轮作2年甘蔗(CK)更加相似, 而轮作2年甘蔗后回种香蕉2年(XJ2)和3年(XJ3)具有相似的土壤细菌构成, 这表明随着香蕉种植年数的增加, 土壤细菌群落构成特征由原来的甘蔗主导型向香蕉主导型递变。
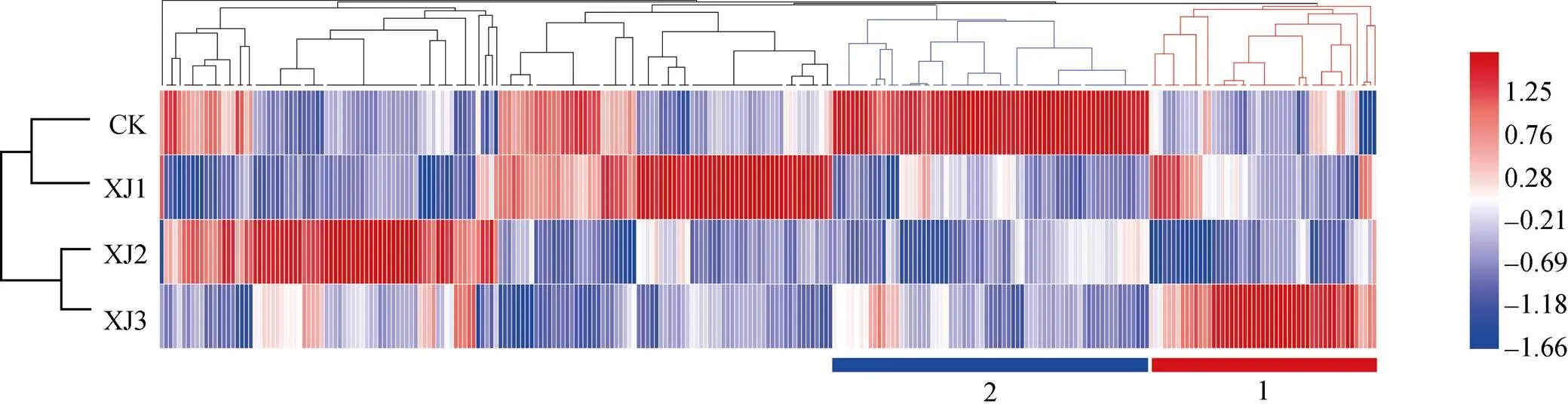
图3 连作香蕉地轮作2年甘蔗后茬土壤16s rDNA(细菌OTUs)丰富度热图
红色表示微生物丰度上调, 蓝色表示微生物丰度下调。Red means microbial abundance increased, blue means microbial abundance decreased.
根据OTU丰度变化趋势可划分为3大类: 第1类(红色纵向聚类)的OTU丰度呈现蓝-白-红的趋势, 表明这类细菌呈现随回种香蕉年数增加而逐年增加的趋势; 而第2类(蓝色纵向聚类)的OTU丰度呈现红-白-蓝的趋势, 表明该类细菌呈现随种植年数的增加而逐年减少的趋势。该结果表明, 土壤中存在特定的微生物会随着香蕉回种年数而呈逐年递增或者递减少的趋势。这些微生物的变化可能与枯萎病的发病率相关。另外, 第3类则是OUT丰度没有随香蕉种植年数增加而呈现规律性变化。
2.3.3 物种分类表达谱
根据OTU的物种注释信息以及OTU在不同样品中的表达(图4), 按照界、门、纲、目、科统计了每一个分类水平上各样品的表达情况, 并绘制出OTU分类树。结果表明, 4个处理的土壤中, 细菌的优势门分别是变形菌门(Proteobacteria, 29.27%)、酸杆菌门(Acidobacteria, 28.20%)和绿弯菌门(Chloroflexi, 12.36%), 这3个门的相对丰度占比高达69%。以上各门微生物向下注释, 优势目分别为酸杆菌目(Acidobacteriales, 6.76%)、黄单胞菌目(Xanthomonadales, 10.23%)、假单胞菌目(Pseudomonadales, 4.29%)、红螺菌目(Rhodospirillales, 7.16%)、纤线杆菌目(Ktedonobacterales, 4.08%)和Thermogemmatisporales (3.31%), 这些优势目的相对丰度占比达35%。
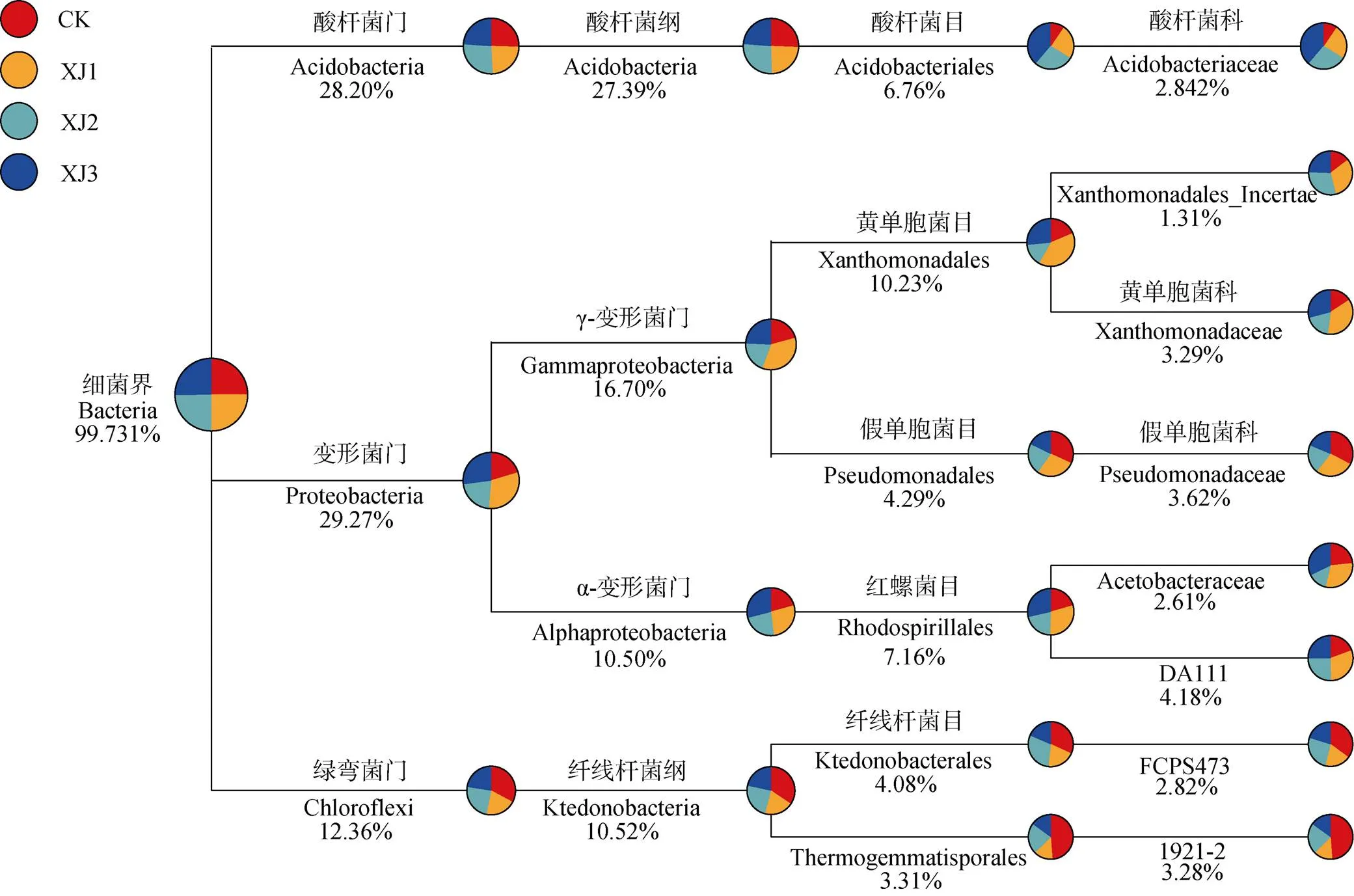
图4 连作香蕉地轮作2年甘蔗后茬土壤16s rDNA(细菌OTUs)表达谱
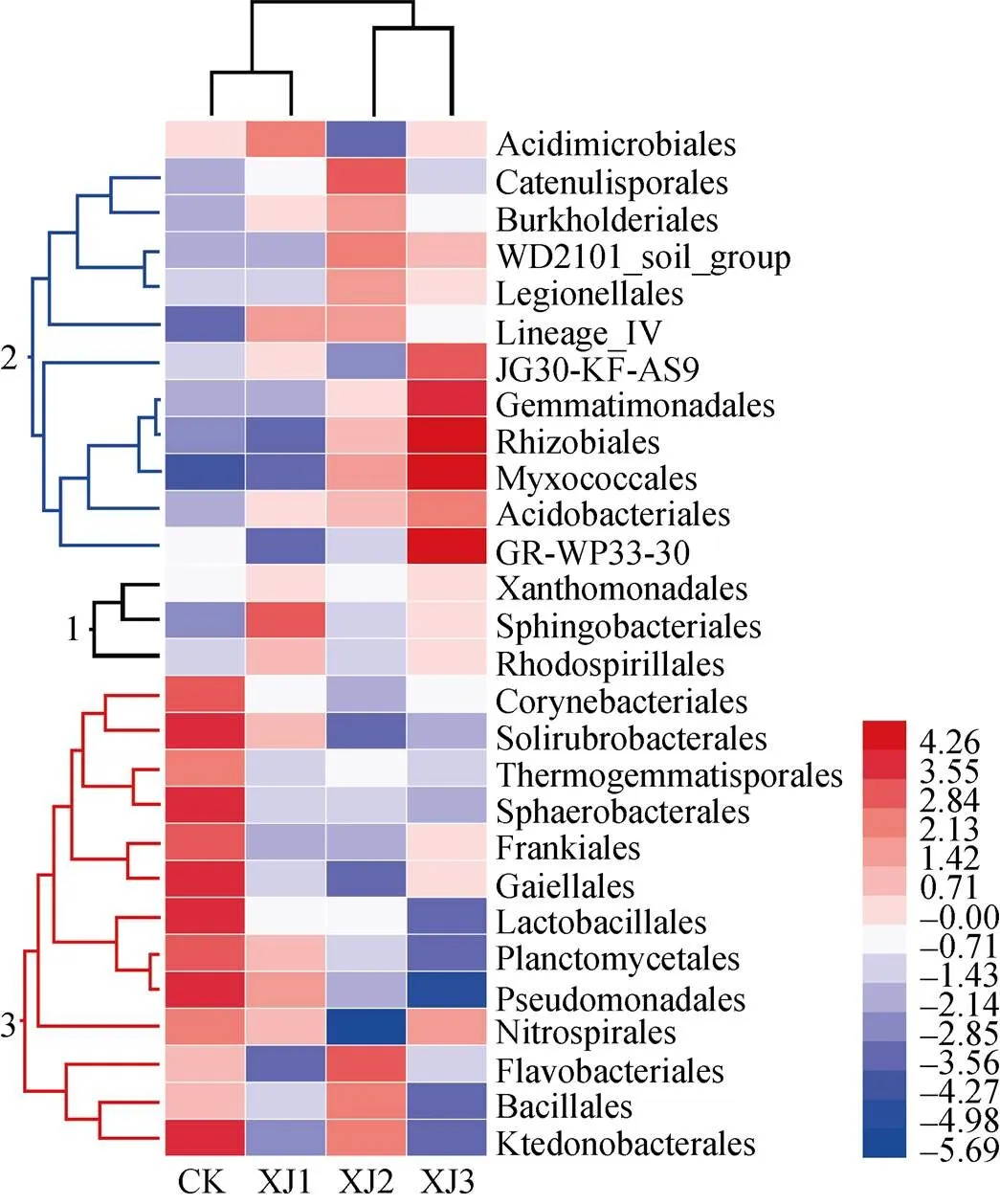
图5 连作香蕉地轮作2年甘蔗后茬土壤细菌目水平上的物种分类热图
