灌浆初期高温胁迫对不同耐热性小麦品种形态和产量的影响*
2019-03-08李升东李华伟王宗帅王法宏孔令安
冯 波, 李升东, 李华伟, 王宗帅, 张 宾, 王法宏, 孔令安
灌浆初期高温胁迫对不同耐热性小麦品种形态和产量的影响*
冯 波, 李升东, 李华伟, 王宗帅, 张 宾, 王法宏**, 孔令安**
(山东省农业科学院作物研究所 济南 250100)
研究灌浆初期高温胁迫对不同耐热性小麦品种的影响, 有助于为耐热稳产性小麦品种选育提供方法, 也可为小麦丰产抗逆栽培技术提供理论参考。以‘济麦22’(JM22)、056852品系(056852)、‘新麦26’(XM26)和‘藁城8901’(GC8901)4个不同耐热性小麦品种(系)为材料, 通过灌浆初期(花后12~14 d)在田间搭建塑料棚模拟高温胁迫, 研究高温胁迫对小麦形态和籽粒产量的影响。高温胁迫处理3 d, 处理最高温度达43.13 ℃, 处理日均温较不搭棚的田间对照温度在胁迫3 d中分别高10.48 ℃、9.71 ℃、9.84 ℃。结果表明: 灌浆初期高温胁迫降低了小麦的植被覆盖指数和冠层叶绿素含量, JM22和056852高温胁迫处理与对照的NDVI值和冠层叶绿素含量在胁迫后差异不显著, 而XM26和GC8901分别显著下降9.66%、6.26%和12.10%、10.73%。高温胁迫后不同耐热性小麦品种(系)籽粒灌浆持续期显著缩短, 与对照相比, JM22、XM26、056852和GC8901籽粒灌浆持续期分别显著缩短1.4 d、2.4 d、0.8 d和3.0 d。千粒重和籽粒产量因高温胁迫显著降低, XM26和GC8901分别比对照产量降低11.43%和10.05%, JM22和056852产量分别降低6.41%和6.93%。综上, 灌浆初期高温胁迫不同程度地加速了耐热性不同小麦品种(系)冠层叶绿素的降解, 缩短了籽粒灌浆天数, 减少了灌浆物质的积累, 降低了籽粒产量。试验材料JM22耐热性和丰产性都较好; 056852品系耐热性较好, 产量一般; XM26和GC8901耐热性较差, 产量较低。
高温胁迫; 小麦; 耐热性; 植株形态; 产量
小麦()是我国主要的粮食作物, 其产量的丰欠直接影响人民生活水平和国家粮食安全。在冬小麦250 d左右的生育期里, 非生物胁迫是造成小麦产量降低的主要原因[1-2], 其中高温胁迫, 尤其是生育后期的高温严重影响小麦产量和品质[3-4]。小麦开花后经常遭遇32 ℃以上的高温, 此时正是小麦产量和品质形成的关键时期。高温胁迫使小麦光合能力下降, 植株老化加速, 灌浆时间缩短, 粒重大大降低[4-8]。高温条件也是诱发干热风的主要因素之一。在我国的黄淮海小麦主产区, 干热风天气在小麦灌浆的初期、中期以及后期均经常出现, 严重影响小麦的产量和品质[8-9]。有报道[10]指出, 到2100年全球平均温度较1980—2000年间的平均温度将增加1.8~4.0 ℃。在全球环境温度不断增高的背景下, 小麦灌浆期遭遇高温胁迫的频率也将呈上升趋势[11], 高温胁迫对小麦的直接危害也将变得日益明显与突出。
前人采取了很多研究方法研究小麦对高温胁迫的反应, 并进行耐热性筛选与评价。高温胁迫后小麦叶绿素荧光变化[12-14]、细胞膜透性变化[15-17]、冠层温度变化[18-19]等被用来作为耐热性筛选和评价的生理指标, 尤其是细胞膜透性的变化在耐热性筛选上被应用较多。依据千粒重和容重计算的热感指数或热害指数也被许多人用来进行耐热性评价[17,20]。仪小梅等[21]根据小麦旗叶功能期和千粒重, 计算出干热风抗逆指数和抗逆系数, 进行小麦抗干热风能力评价。虽然关于高温胁迫对小麦产量的影响研究较多, 但许多研究者在进行小麦耐热性筛选与评价时只重视耐热性而忽视产量, 或者由于试验方法仅限于实验室测定等原因, 导致目前尚没有统一的、简单易行的评价小麦耐热性的指标方法。小麦是群体作物, 本研究在考察小麦耐热性时, 将群体冠层叶绿素含量、群体NDVI值作为评价小麦耐热性的指标, 同时与灌浆特性和产量结果联系分析, 希望为耐热丰产稳产性小麦品种选育提供方法, 也为小麦丰产抗逆栽培技术提供理论参考。
1 材料与方法
1.1 试验设计
试验于2011—2013年、2017—2018年在山东省农业科学院济南试验田进行。供试材料为耐热性较好的‘济麦22’(JM22)、056852品系(056852)以及耐热性较差的‘新麦26’(XM26)和‘藁城8901’ (GC8901)。JM22和056852在2011年、2012年、2017年都是10月6日播种, 而XM26和GC8901都是10月9日播种, 花期一致。小区面积1.5 m×9 m, 基本苗165株∙m-2, 行距25 cm。完全随机排列, 3次重复。采取田间搭棚的方法进行高温处理(Str), 以不搭棚的大田小区作为对照(CK), 田间自动气象站(CM10l Campbell Scientific, Logan, UT, USA)记录对照温度。高温处理时期为开花后12~14 d, 即小麦灌浆的初期搭建塑料大棚, 大棚采用刷漆铁骨架作构架, 塑料薄膜的厚度为70 μm, 透光率为92.7%, 大棚高度1.7 m, 面积为18 m×9 m。在每个小区冠层上方50 cm处均匀悬挂3个温度计, 每日8:00—20:00每2 h调查记录小区温度。高温处理结束后, 移除覆盖大棚的薄膜。大棚两侧距顶部20 cm处每50 cm留若干30 cm×30 cm的小窗口, 以保证棚内良好的通风状态。
高温处理期间, 处理与对照田的温度如图1所示, 2012年、2013年以及2018年处理3 d期间棚内外的平均气温差分别是: 10.48 ℃、9.71 ℃、9.84 ℃, 10.65 ℃、8.99 ℃、9.96 ℃和2.00 ℃、2.37 ℃、1.55 ℃。3年高温处理期间均无降水。播前每公顷底施有机肥3 750 kg, 纯氮120 kg, P2O5105 kg和K2O 105 kg, 拔节期每公顷追施纯氮105 kg, 越冬期和拔节期各灌水60 mm。6月4日收获。每个小区分为两部分, 一部分材料用作数据测定和取样分析, 另一部分成熟时测产。
1.2 测定项目与方法
1.2.1 植株形态观察记录
高温处理3 d结束后, 移除覆盖的大棚薄膜, 即刻观察记录处理与对照叶片、穗部、穗下节间的形状与颜色变化, 并用相机拍摄植株冠层照片。
1.2.2 叶片和穗部的叶绿素含量测定
处理结束后取高温处理与对照植株, 迅速带回实验室, 称旗叶、穗下节间、基部第三小穗以及旗叶鞘各0.5 g左右, 放入1∶1的无水乙醇和丙酮混合提取液中, 常温、黑暗条件下充分提取超过24 h, 于663 nm、645 nm和470 nm处测定吸光值, 叶绿素含量按照Arnon[22]的公式计算。
1.2.3 归一化植被指数(NDVI)值测定
处理结束后, 在小麦灌浆的中期(5月18日和5月21日)以及后期(5月29日和6月2日), 即高温胁迫结束后第4 d、7 d、15 d和19 d, 采用GreenSeeker手持式光谱仪(美国NTech公司)测定NDVI值, 传感器与冬小麦冠层保持平行, 距离冠层高度60 cm, 顺麦垄方向采集NDVI值获取3组数据。每小区固定3组测定区域。
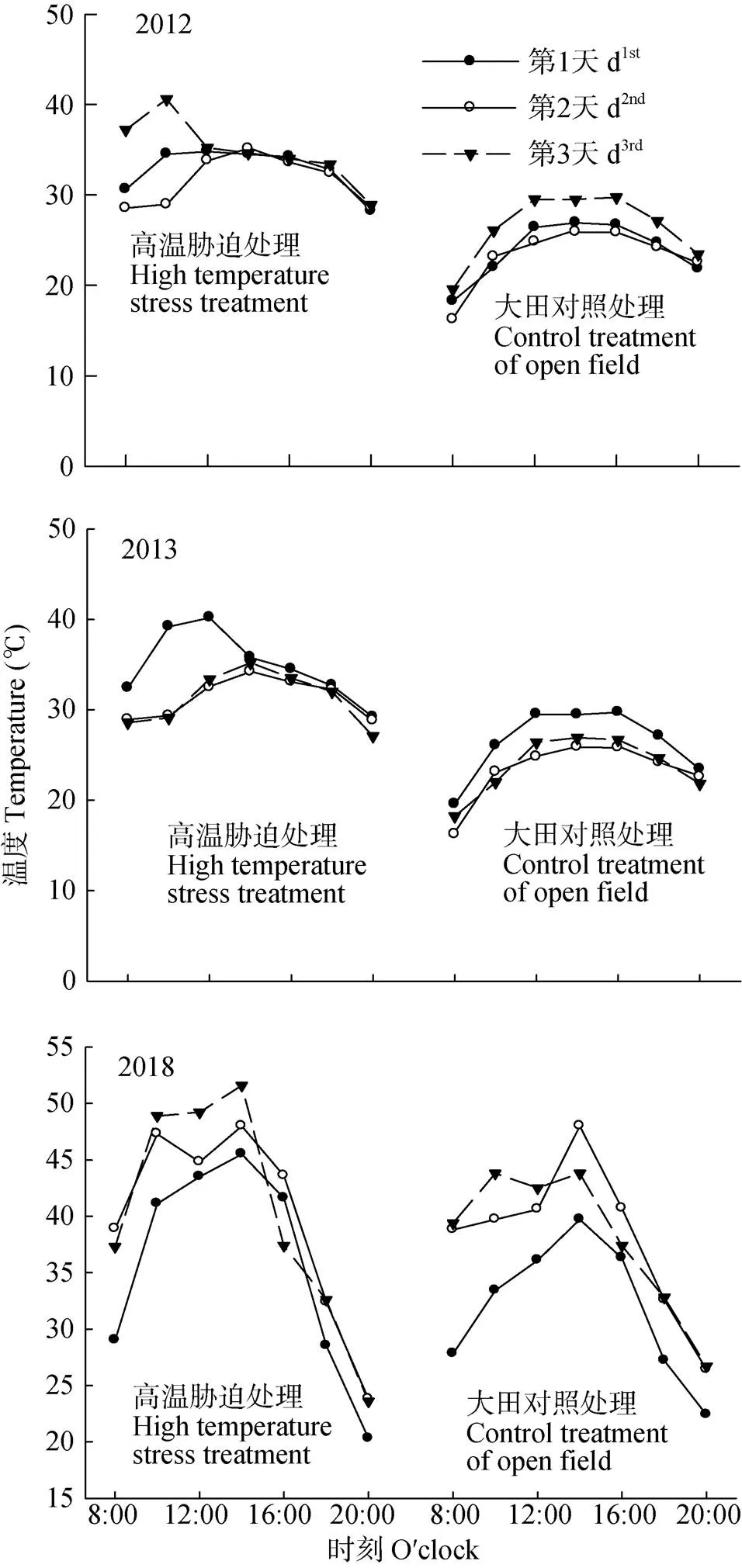
图1 2012年、2013年、2018年试验期间高温胁迫处理与对照处理的气温日变化
试验在2012年处理的第3天12:00时大棚内温度均超过45 ℃, 因此12:00—14:00揭开高温棚, 以解除超高温胁迫。因此, 2012年第3天高温胁迫处理12:00和14:00数据是揭开大棚后的温度数据。The proof of the artificial greenhouse was uncovered from 12:00 to 14:00 at the 3rdday in 2012 because of the extreme high temperature over 45 ℃inside, so as to alleviate the damage by the super high temperature stress. Therefore, the data at time 12:00 and time 14:00 of the 3rdday in 2012 were the temperature after the greenhouse proof was removed.
1.2.4 籽粒硬度和直径测试
成熟期收获后, 取高温处理与对照的籽粒进行籽粒硬度和直径测试, 采用4100型单籽粒硬度仪(瑞典PERTEN公司)测定。每个重复测定300个籽粒, 取平均值, 样品的籽粒硬度用平均硬度指数表示。籽粒直径用游标卡尺取最宽处测定, 每个重复测定50个籽粒, 取平均值。
1.2.5 小麦籽粒灌浆速率的测定
开花期对开花一致的小麦单茎标记, 开花至成熟期每隔5 d每个重复取标记穗5穗, 105 ℃杀青10 min, 70 ℃烘至恒重, 称籽粒重, 用于籽粒灌浆速率的测定。籽粒的增重动态参照李世清等[23]的方法, 用三次多项方程式=+1+22+33进行模拟, 其中1、2、3是根据品种的不同确定常数,代表粒重,代表花后天数。并推导出当=-2/33时最大灌浆速率max=1-22/33, 最大灌浆速率持续期dmax=-2(22-331)1/2/33。
1.3 数据分析
因2011—2013两年度的试验结果趋势基本一致, 而2017年高温处理与对照日均温差较低, 试验结果与2011—2013年重演性不好, 因此本文采用2011—2012年度的试验数据进行分析。采用DPS v17.10软件对试验数据进行方差分析和多重比较(LSD法), 用SigmaPlot 10.0软件作图。
2 结果与分析
2.1 高温胁迫对不同耐热性小麦植株形态的影响
不同耐热品种对高温胁迫的反应不同(图2)。高温胁迫对056852品系植株形态影响最小, 植株个体的差异很难分辨, 从群体上看较对照略微失绿变黄。高温对JM22的外部形态有较小的影响, 与对照相比, 高温胁迫的芒、穗部以及少量叶尖颜色略微失绿, 颜色发黄。高温对XM26的植物形态影响最大, 胁迫处理的芒几乎全部变黄白色, 穗部明显失绿变黄, 叶尖失绿干枯面积较大。高温对GC8901植株形态的影响也较大, 主要以芒变白和部分叶片叶尖失绿为特征, 穗部与对照相比也明显的失绿变黄。

图2 灌浆初期高温胁迫对不同耐热品种(系)冬小麦植株形态的影响
2.2 灌浆初期高温对不同耐热性小麦冠层叶绿素含量的影响
高温胁迫后不同耐热性品种小麦冠层叶绿素含量下降(图3)。XM26和GC8901冠层叶绿素含量下降程度显著大于JM22和056852。冠层各光合器官比较发现, 穗部的叶绿素含量受高温胁迫的影响变化最大, 较冠层其他绿色器官叶绿素含量下降显著。XM26和GC8901穗部叶绿素含量分别下降12.10%和10.73%, JM22和056852下降5.29%和1.95%; 穗下节间的叶绿素含量下降最小, 变化较大的XM26和GC8901下降5.65%和7.18%, JM22和056852变化较小, 分别下降0.61%和3.13%; 旗叶和旗叶鞘的叶绿素含量变化居于穗部和穗下节间之间。
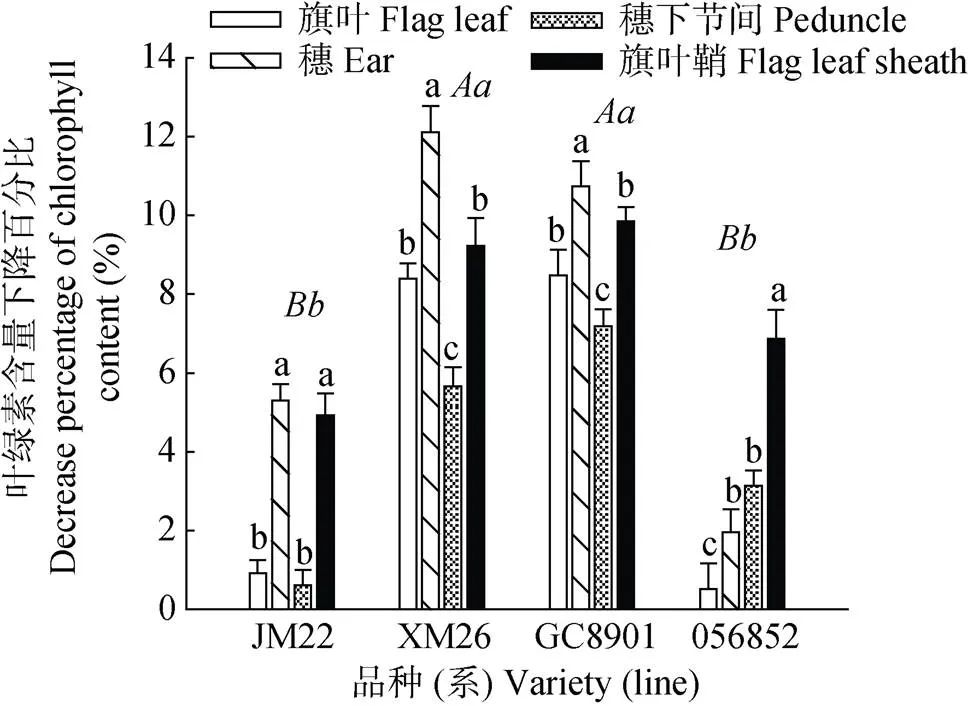
图3 灌浆初期高温胁迫后不同耐热性小麦品种(系)冠层不同器官叶绿素含量较对照下降百分比
正体不同小写字母表示同一品种不同器官间差异达5%显著水平。斜体不同大、小写字母分别表示不同品种间差异达1%或5%显著水平。Different normal lowercase letters mean significant differences among different organs for the same cultivar at 0.05 level. Different italic uppercase or lowercase letters mean significant differences among different varieties at 0.01 or 0.05 level.
2.3 灌浆初期高温胁迫对不同耐热性小麦植被覆盖指数的影响
灌浆初期连续3 d的高温胁迫降低了小麦的植被覆盖指数(NDVI, 图4), 但不同品种影响程度不同。高温胁迫处理结束后第4 d(5月18日), XM26和GC8901的NDVI显著低于各自对照, 分别较对照降低9.66%和10.28%, JM22和056852分别下降4.45%和2.53%, 高温处理与对照无显著差异。品种间差异不显著。胁迫结束后第7 d(5月21日), 高温处理与对照的差异更加显著, 其中XM26和GC8901高温处理与对照差异极显著(>0.01), JM22和056852高温处理与对照差异显著(<0.05)。随着生育进程推进, 至高温胁迫后15 d(5月29日), 各品种(系)NDVI降低,且参试品种(系)高温处理与对照差异均显著。XM26和GC8901两品种之间NDVI差异不显著, 但两品种均显著低于JM22和056852。至高温胁迫后19 d(6月2日), JM22和056852品系高温处理NDVI显著高于对照; 而XM26和GC8901高温处理与对照差异不显著, 并且该两个品种间无显著差异。这是由于自5月下旬以来环境温度越来越高, 加速了不耐热品种(系)的早衰, 耐热性较差的品种(系)的对照也由于外界环境温度升高而趋于成熟, 使得高温处理与对照差异不显著。这也进一步证实JM22和056852耐热性更好, XM26和GC8901耐热性不如前两者。
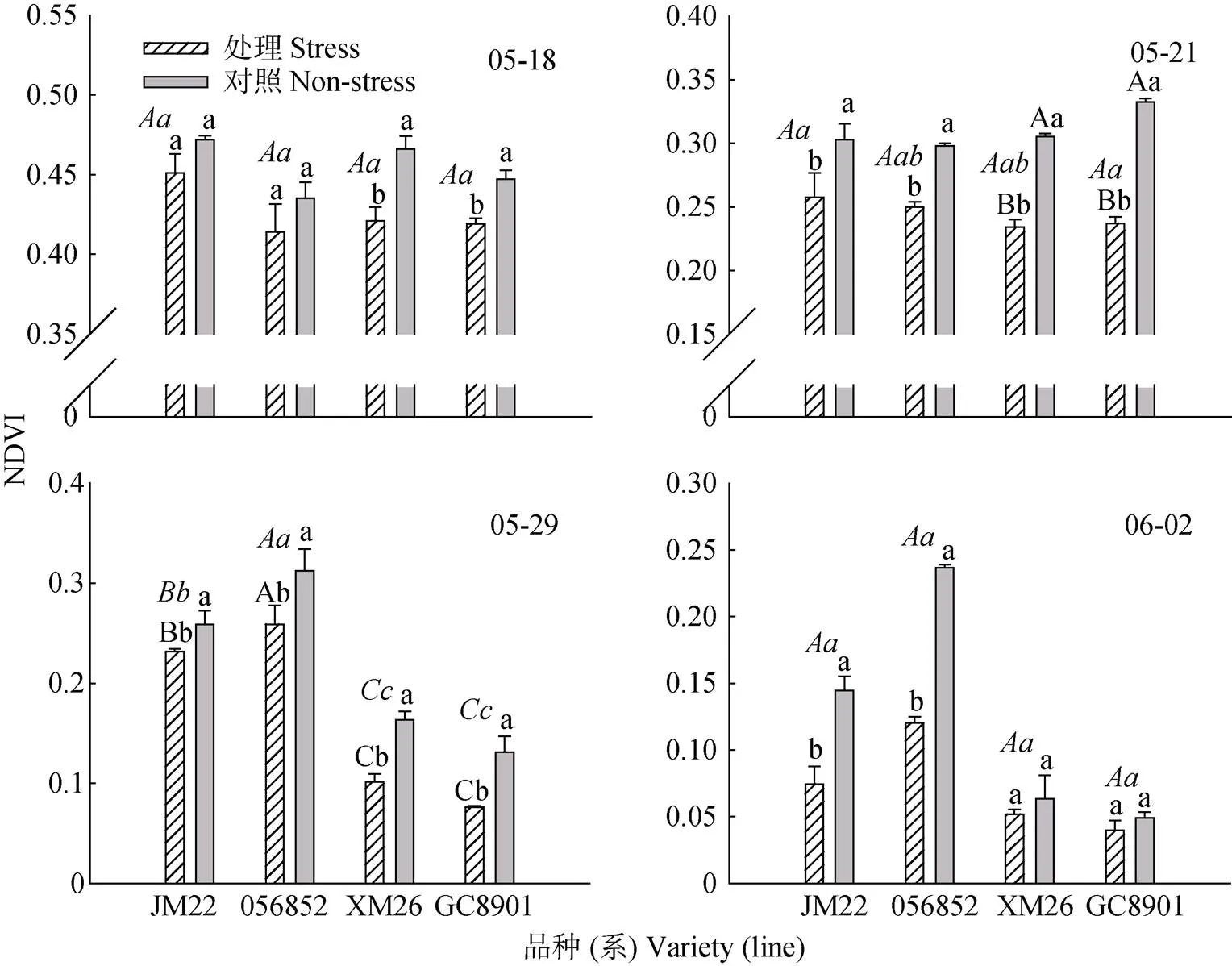
图4 灌浆初期高温胁迫对不同耐热小麦品种(系)不同时间植被覆盖指数(NDVI)的影响
05-18、05-21、05-29和06-02分别为高温胁迫后4 d、7 d、15 d和19 d。正体不同大、小写字母表示同一品种不同处理间差异达1%极显著或5%显著水平。斜体不同大小写字母分别表示不同品种间差异达1%或5%显著水平。05-18, 05-21, 05-29 and 06-02 are 4, 7, 15 and 19 days after high temperature stress. Different normal upper and lowercase letters above each bar mean significant differences between treatments for the same cultivar at 0.01 level and 0.05 level. Different italic uppercase and lowercase letters mean differences among different varieties at 0.01 or 0.05 level.
2.4 灌浆初期高温胁迫对不同耐热性小麦籽粒形态的影响
如图5所示, 受高温胁迫的影响, 不同耐热性小麦的籽粒形态也发生了程度不等的变化。变化最大的是XM26, 高温胁迫后籽粒变小, 籽粒饱满度下降, 籽粒颜色明显变深。其次是GC8901, 其籽粒大小、饱满度、颜色较对照变化程度仅次于XM26。JM22和056852的籽粒形态与对照相比变化程度较小。

图5 灌浆初期高温胁迫对不同耐热小麦品种(系)籽粒形态的影响
2.5 灌浆初期高温胁迫对不同耐热性小麦品种(系)籽粒硬度及直径的影响
高温胁迫增加了不同耐热性小麦品种(系)的籽粒硬度指数, 降低了小麦的籽粒直径(表1)。不同耐热性品种(系)间籽粒硬度的变化幅度不同, 056852籽粒硬度增加幅度最大, 达极显著水平, 且显著大于其他3个品种(系); XM26、GC8901和JM22分别增加8.54%、6.72%和6.04%,差异显著。JM22和GC8901间差异不显著。品种间直径大小差异极显著。高温胁迫后, XM26和GC8901高温胁迫处理直径较对照籽粒极显著降低; 056852和JM22高温胁迫处理较对照显著降低。
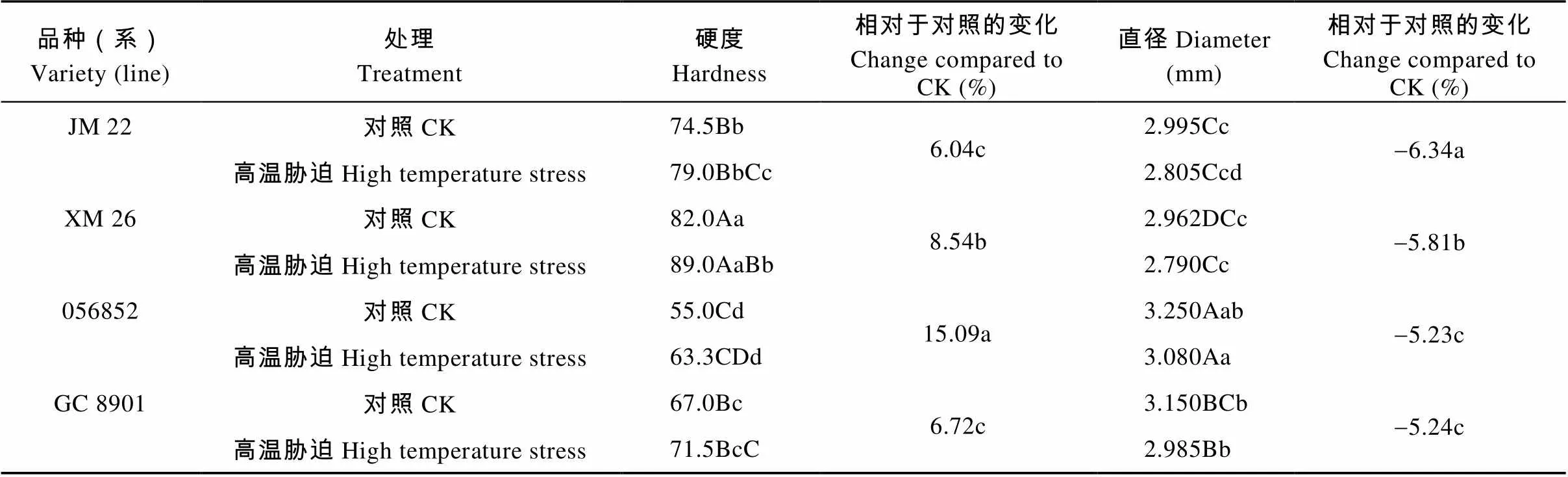
表1 灌浆初期高温胁迫对不同耐热小麦品种(系)籽粒硬度及直径的影响
不同大小写字母分别表示高温胁迫与对照间差异在<0.01水平和<0.05水平显著。Different lowercase and capital letters indicate significant differences between CK and high temperature stress treatments at< 0.05 and< 0.01 levels, respectively.
2.6 灌浆初期高温胁迫对小麦籽粒千粒重动态的影响
高温胁迫后不同耐热品种小麦籽粒灌浆速率无降低(图6)。花后15~20 d, JM22和056852处理与对照相比灌浆速率变化不大, XM26和GC8901灌浆速率下降较大。JM22、XM26、GC8901的千粒重都在花后30 d时达最大值, 056852花后35 d时籽粒增重曲线仍呈上升趋势, 灌浆持续时间较长。与各自对照(CK)相比, 灌浆初期的高温处理均显著降低了不同耐热品种小麦的千粒重。小麦成熟时期, JM22、056852、GC8901和XM26 4个品种高温胁迫处理的千粒重比对照分别下降4.39%、5.93%、7.64%和8.03%。
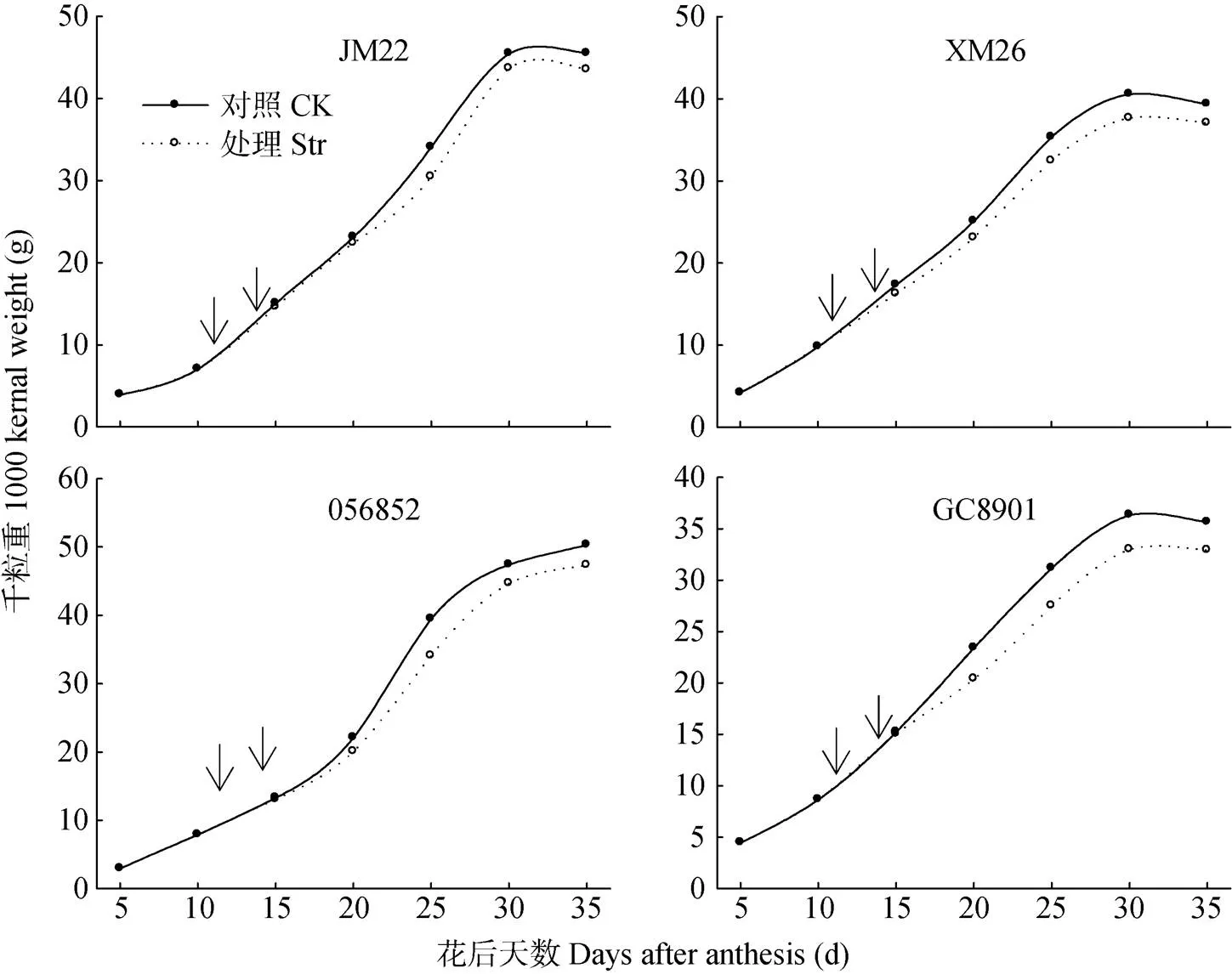
图6 灌浆初期高温胁迫(Str)对不同耐热性小麦品种(系)粒重增长动态的影响
箭头之间为高温胁迫处理时间: 花后12-14 d。Space between two narrows is the high temperature stress treatment time, from 12 to 14 days after anthesis.
2.7 灌浆初期高温胁迫对小麦籽粒增重动态及最大灌浆速率变化的影响
用方程式=+1+22+33模拟籽粒增长动态, 并计算灌浆持续期和最大灌浆速率及其出现时间。由表2可以看出, 灌浆初期高温胁迫显著缩短了不同耐热品种(系)的籽粒灌浆持续期, 与对照相比, JM22、XM26、056852和GC8901分别显著缩短1.4 d、2.4 d、0.8 d和3.0 d。056852灌浆持续期极显著大于JM22、XM26和GC8901, JM22显著大于XM26和GC8901, XM26和GC8901之间无差异显著。高温胁迫使不同耐热品种的最大灌浆速率时间提前, JM22、056852、XM26和GC8901分别极显著提前1.7 d、2.1 d、1.7 d和1.8 d。XM26和GC8901的最大灌浆速率出现时间差异不显著, 但其灌浆速率持续期极显著低于JM22和056852。JM22、056852、XM26和GC8901的最大灌浆速率降低, 与对照相比分别降低10.53%、5.56%、35.29%和29.41%, 但品种(系)间与处理间差异均不显著。
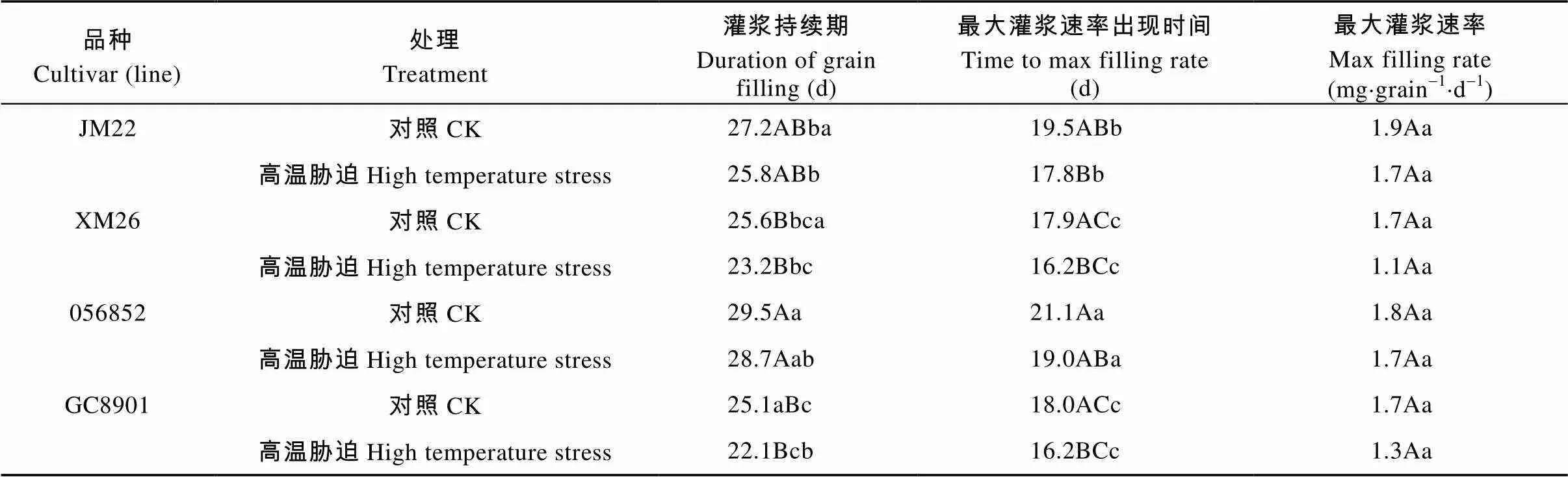
表2 灌浆初期高温胁迫对不同品种(系)小麦籽粒增重模型及灌浆参数的影响
不同大、小写字母分别表示高温胁迫与对照间差异在<0.01水平和<0.05水平显著。Different lowercase and capital letters indicate significant differences between CK and high temperature stress treatments at< 0.01 and< 0.05 levels, respectively.
2.8 高温胁迫对不同耐热性小麦籽粒产量的影响
灌浆初期高温胁迫显著降低了小麦千粒重, 从而造成了产量的降低(表3)。与对照相比, XM26、GC8901、JM22和056852籽粒产量受高温胁迫分别极显著减产11.43%、10.05%、6.41%和6.93%。受高温胁迫的影响千粒重降幅在品种(系)间差异极显著, XM26和GC8901籽粒产量降幅差异显著, 都极显著大于JM22和056852, 后两者之间差异也达显著水平。
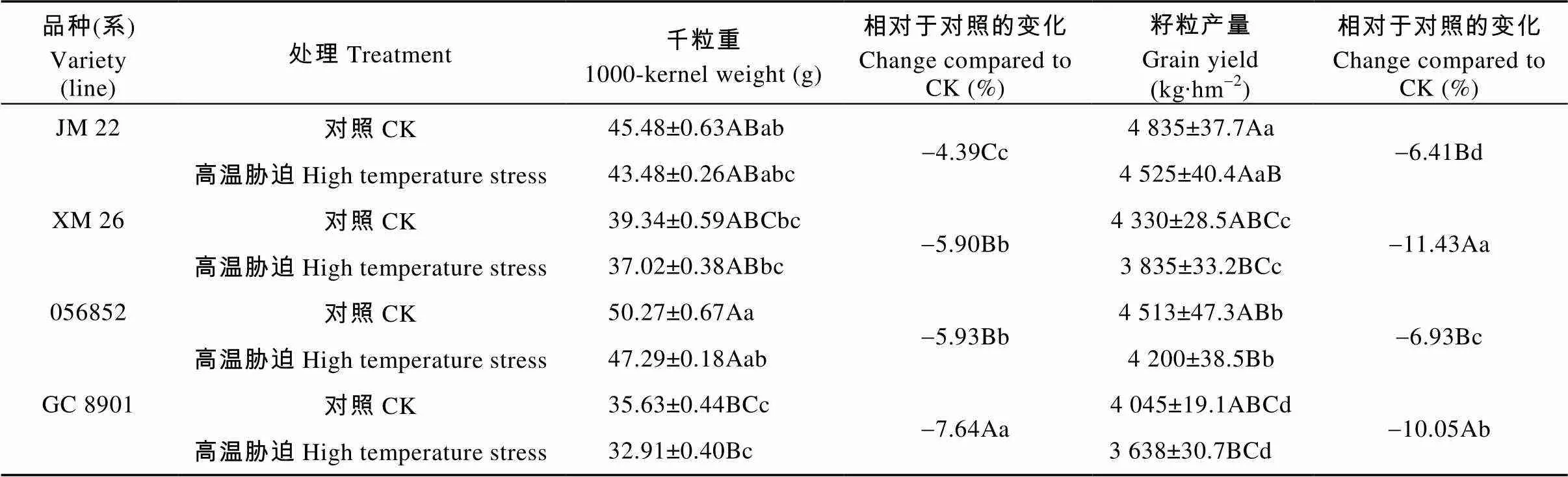
表3 灌浆初期高温胁迫对不同耐热品种(系)千粒重和产量的影响
不同大小写字母分别表示高温胁迫与对照间差异在<0.01水平和<0.05水平显著。Different lowercase and capital letters indicate significant differences between CK and high temperature stress treatments at< 0.05 and< 0.01 levels, respectively.
3 讨论与结论
在全球气温不断升高的背景下, 极端高温事件频发[24-26]。温度每升高1 ℃, 全球小麦减产6%[27]。IPCC(2007年)第4次报告已将高温胁迫作为威胁全球粮食安全的重要因子之一。高温胁迫对我国小麦的危害主要表现为干热风, 可使小麦灌浆期缩短、粒重降低, 品质下降。有前人研究表明, 灌浆中后期是小麦热胁迫的高发期, 对小麦生产影响较大[3]。然而, 也有少数研究表明灌浆期前期比后期对高温更敏感[28-29]。且全国农业气象月报显示, 2014— 2017年5月上、中旬小麦灌浆前期, 黄淮海大部分地区出现了干热风天气。因此研究灌浆前期高温胁迫对小麦的影响是当前的迫切需求。
许多研究表明, 环境条件对冬小麦生产具有显著的影响[3,7-9,28-31]。灌浆期高温胁迫主要通过影响籽粒灌浆特性来影响粒重[6,32-34]。李世清等[23]认为籽粒灌浆持续期、灌浆速率受环境因素(特别是温度)调控, 籽粒灌浆速率与粒重呈正相关, 并且不同基因型间粒重的差异主要与灌浆速率的差别有关。本试验发现, 灌浆初期连续3 d的高温处理, 使4个参试品种(系)JM22、056852、XM26和GC8901籽粒灌浆特性发生改变, 籽粒灌浆持续期的天数都减少, 这与前人[35-36]研究结果一致。不同小麦品种对高温胁迫耐性不同[35-37]。本试验结果表明, 与各自对照相比, JM22和056852灌浆持续期缩短时间较小, XM26和GC8901灌浆持续期缩短时间较多。根据冯素伟等[38]将小麦灌浆过程分为渐增期、快增期和缓增期的划分, 本试验在灌浆初期的高温胁迫使籽粒灌浆快增期的灌浆速率降低, 尤其是XM26和GC8901受热胁迫影响的时间早于JM22和056852。高温胁迫还使供试品种的最大灌浆速率降低。灌浆时间减少和灌浆速率下降是高温胁迫后小麦粒重显著降低的直接原因。粒重降低与籽粒形态变化相吻合。高温胁迫后灌浆物质充实不足导致籽粒饱满度下降, 籽粒直径下降, 硬度指数升高。
灌浆期高温胁迫对小麦籽粒灌浆产生负面影响主要是由于高温胁迫后冠层的呼吸速率加剧, 呼吸消耗的同化产物增加, 籽粒灌浆的同化产物减少。姜雨萌等[39]研究发现冬小麦灌浆期间的高温胁迫对冠层碳同化具有显著的抑制作用。穗、茎、叶是冠层碳同化的主体, 即籽粒灌浆最主要的光合器官。持绿型小麦产量较普通小麦产量高的重要原因就是生育后期冠层具有较多的绿叶面积和数目, 净光合速率高, 光合功能维持时间长, 干物质积累多, 并且茎秆和叶鞘的储备转运能力强[40-41]。耐热性好的小麦即使在高温下也能够保持较高的叶绿素含量和光合效率, 并且具有较长的灌浆时间[34,42]。本研究发现, 灌浆期高温胁迫后不同耐热小麦植株冠层黄化失绿, 冠层叶和非叶光合器官的叶绿素含量降低。穗部的叶绿素含量降低最多, 而穗下节间的叶绿素含量下降最小, 旗叶和旗叶鞘的变化居中。不同耐热性小麦品种冠层叶绿素含量变化受高温胁迫的影响不同。GC8901和XM26冠层叶叶绿素含量下降显著大于JM22和056852。冠层穗、茎鞘、叶的衰老影响同化物质的合成与转运。小麦干物质转运受生育后期温度影响较大, 花后高温胁迫使小麦籽粒干物质分配量显著降低。因此推测高温胁迫也将降低叶片、茎鞘以及颖壳中贮存同化物向籽粒的转运能力, 降低籽粒产量。植被覆盖指数可以无损快速地估算作物冠层叶绿素含量[43-45], 本试验中高温胁迫后小麦群体的植被覆盖指数降低, 证实了冠层叶绿素含量降低, 叶面积指数减少。
在品种选育时, 群体NDVI值、植株冠层叶绿素含量可以作为小麦耐热性评价的参考指标。生产上进行品种推广时, 应综合考虑品种(系)耐热性与丰产性, 本研究中XM26和GC8901耐热性较差, 产量也较低; 056852品系虽然耐热性较好, 但是产量一般; JM22耐热性较好, 产量表现也最好。
[1] KOÇ M, BARUTCULAR C, TIRYAKIOGLU M. Possible heat-tolerant wheat cultivar improvement through the use of flag leaf gas exchange traits in a Mediterranean environment[J]. Journal of the Science of Food and Agriculture, 2008, 88(9): 1638–1647
[2] DEL MORAL L F G, RHARRABTI Y, VILLEGAS D, et al. Evaluation of grain yield and its components in durum wheat under Mediterranean conditions[J]. Agronomy Journal, 2003, 95(2): 266–274
[3] 李永庚, 于振文, 张秀杰, 等. 小麦产量与品质对灌浆不同阶段高温胁迫的响应[J]. 植物生态学报, 2005, 29(3): 461–466 LI Y G, YU Z W, ZHANG X J, et al. Response of yield and quality of wheat to heat stress at different grain filling stages[J]. Acta Phytoecologica Sinica, 2005, 29(3): 461–466
[4] 苗建利, 王晨阳, 郭天财, 等. 高温与干旱互作对两种筋力小麦品种籽粒淀粉及其组分含量的影响[J]. 麦类作物学报, 2008, 28(2): 254–259 MIAO J L, WANG C Y, GUO T C, et al. Effects of post-anthesis interactions of high temperature and drought stresses on content and composition of grain starch in two wheat cultivars with different gluten strength[J]. Journal of Triticeae Crops, 2008, 28(2): 254–259
[5] WARDLAW I F. Interaction between drought and chronic high temperature during kernel filling in wheat in a controlled environment[J]. Annals of Botany, 2002, 90(4): 469–476
[6] 胡阳阳, 卢红芳, 刘卫星, 等. 灌浆期高温与干旱胁迫对小麦籽粒淀粉合成关键酶活性及淀粉积累的影响[J]. 作物学报, 2018, 44(4): 591–600 HU Y Y, LU H F, LIU W X, et al. Effects of high temperature and water deficiency during grain filling on activities of key starch synthesis enzymes and starch accumulation in wheat[J]. Acta Agronomica Sinica, 2018, 44(4): 591–600
[7] 肖登攀, 陶福禄, 沈彦俊, 等. 华北平原冬小麦对过去30年气候变化响应的敏感性研究[J]. 中国生态农业学报, 2014, 22(4): 430–438 XIAO D P, TAO F L, SHEN Y J, et al. Sensitivity of response of winter wheat to climate change in the North China Plain in the last three decades[J]. Chinese Journal of Eco-Agriculture, 2014, 22(4): 430–438
[8] 成林, 张志红, 方文松. 干热风对冬小麦灌浆速率和千粒重的影响[J]. 麦类作物学报, 2014, 34(2): 248–254 CHENG L, ZHANG Z H, FANG W S. Effects of dry-hot wind on grain filling speed and 1000-kernel weight of winter wheat[J]. Journal of Triticeae Crops, 2014, 34(2): 248–254
[9] AL-KHATIB K, PAULSEN G M. Photosynthesis and productivity during high-temperature stress of wheat genotypes from major world regions[J]. Crop Science,1990, 30(5): 1127–1132
[10] IPCC. Summary for policymakers[C]//Climate Change 2007: The Physical Science Basis. Contribution of Working Group Ⅰ to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. New York: IPCC, 2007
[11] 谭凯炎, 房世波, 任三学. 增温对华北冬小麦生产影响的试验研究[J]. 气象学报, 2012, 70(4): 902–908 TAN K Y, FANG S B, REN S X. Experiment study of winter wheat growth and yield response to climate warming[J]. Acta Meteorologica Sinica, 2012, 70(4): 902–908
[12] BAKER N R, ROSENQVIST E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities[J]. Journal of Experimental Botany, 2004, 55(403): 1607–1621
[13] ELHANI S, RHARRABTI Y, DEL MORAL L F G, et al. Evolution of chlorophyll fluorescence parameters in durum wheat as affected by air temperature[M]//ROYO C, NACHIT M, DI FONZO N, et al. Durum Wheat Improvement in the Mediterranean Region: New Challenges. Zaragoza: CIHEAM-Options Méditerranéennes, 2004: 275–277
[14] CUI L J, LI J L, FAN Y M, et al. High temperature effects on photosynthesis, PSⅡ functionality and antioxidant activity of twocultivars with different heat susceptibility[J]. Botanical Studies, 2006, 47(1): 61–69
[15] THIAW S, HALL A E. Comparison of selection for either leaf-electrolyte-leakage or pod set in enhancing heat tolerance and grain yield of cowpea[J]. Field Crops Research, 2004, 86(2/3): 239–253
[16] DHANDA S S, MUNJAL R. Inheritance of cellular thermotolerance in bread wheat[J]. Plant Breeding, 2006, 125(6): 557–564
[17] 耿晓丽, 张月伶, 臧新山, 等. 北方冬麦区与黄淮北片优良小麦品种(系)耐热性评价[J]. 麦类作物学报, 2016, 36(2): 172–181GENG X L, ZHANG Y L, ZANG X S, et al. Evaluation the thermos tolerance of the wheat (L.) cultivars and advanced lines collected from the northern China and north area of Huanghuai winter wheat regions[J]. Journal of Triticeae Crops, 2016, 36(2): 172–181
[18] MAHAN J R, BURKE J J. Active management of plant canopy temperature as a tool for modifying plant metabolic activity[J]. American Journal of Plant Sciences, 2015, 6(1): 53531
[19] REYNOLDS M P, BALOTA M, DELGADO M I B, et al. Physiological and morphological traits associated with spring wheat yield under hot, irrigated conditions[J]. Australian Journal of Plant Physiology, 1994, 21(6): 717–730
[20] 韩利明, 张勇, 彭惠茹, 等. 从产量和品质性状的变化分析北方冬麦区小麦品种抗热性[J]. 作物学报, 2010, 36(9): 1538−1546HAN L M, ZHANG Y, PENG H R, et al. Analysis of heat resistance for cultivars from North China Winter Wheat Region by yield and quality traits[J]. Acta Agronomica Sinica, 2010, 36(9): 1538−1546
[21] 仪小梅, 孙爱清, 韩晓玉, 等. 黄淮麦区小麦主推品种(系)干热风抗性鉴定[J]. 麦类作物学报, 2015, 35(2): 274–284YI X M, SUN A Q, HAN X Y, et al. Identification of dry-hot wind resistance of major wheat cultivars (lines) in Huanghuai wheat region[J]. Journal of Triticeae Crops, 2015, 35(2): 274–284
[22] ARNON D T. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in[J]. Plant Physiology,1949, 24(1): 1–15
[23] 李世清, 邵明安, 李紫燕, 等. 小麦籽粒灌浆特征及影响因素的研究进展[J]. 西北植物学报, 2003, 23(11): 2031–2039 LI S Q, SHAO M A, LI Z Y, et al. Review of characteristics of wheat grain filling and factors to influence it[J]. Acta Botanica Boreali-Occidentalia Sinica, 2003, 23(11): 2031–2039
[24] TEBALDI C, HAYHOE K, ARBLASTER J M, et al. Going to the extremes: An intercomparison of model-simulated historical and future changes in extreme events[J]. Climatic Change, 2006, 79(3/4): 185–211
[25] JIANG Z H, SONG J, LI L, et al. Extreme climate events in China: IPCC-AR4 model evaluation and projection[J]. Climatic Change, 2012, 110(1/2): 385–401
[26] 丁一汇, 任国玉, 石广玉, 等. 气候变化国家评估报告(Ⅰ): 中国气候变化的历史和未来趋势[J]. 气候变化研究进展, 2006, 2(1): 3–8 DING Y H, REN G Y, SHI G Y, et al. National assessment report of climate change (Ⅰ): Climate change in China and its future trend[J]. Advances in Climate Change Research, 2006, 2(1): 3–8
[27] ASSENG S, EWERT F, MARTRE P, et al. Rising temperatures reduce global wheat production[J]. Nature Climate Change, 2015, 5(2): 143–147
[28] 赵俊芳, 赵艳霞, 郭建平, 等. 基于干热风危害指数的黄淮海地区冬小麦干热风灾损评估[J]. 生态学报, 2015, 35(16): 5287–5293 ZHAO J F, ZHAO Y X, GUO J P, et al. Assessment of the yield loss of winter wheat caused by dry-hot wind in Huanghuaihai Plain based on the hazard index of dry-hot wind[J]. Acta Ecologica Sinica, 2015, 35(16): 5287–5293
[29] 杨绚, 汤绪, 陈葆德, 等. 气候变暖背景下高温胁迫对中国小麦产量的影响[J]. 地理科学进展, 2013, 32(12): 1772–1779 YANG X, TANG X, CHEN B D, et al. Impacts of heat stress on wheat yield due to climatic warming in China[J]. Progress in Geography, 2013, 32(12): 1772–1779
[30] ARAUS J L, SLAFER G A, REYNOLDS M P, et al. Plant breeding and drought in C3 cereals: What should we breed for?[J]. Annals of Botany, 2002, 89: 925–940
[31] 刘霞, 尹燕枰, 姜春明, 等. 花后不同时期弱光和高温胁迫对小麦旗叶荧光特性及籽粒灌浆进程的影响[J]. 应用生态学报, 2005, 16(11): 2117–2121 LIU X, YIN Y P, JIANG C M, et al. Effects of weak light and high temperature stress after anthesis on flag leaf chlorophyll fluorescence and grain fill of wheat[J]. Chinese Journal of Applied Ecology, 2005, 16(11): 2117–2121
[32] FAROOQ M, BRAMLEY H, PALTA J A, et al. Heat stress in wheat during reproductive and grain-filling phases[J]. Critical Reviews in Plant Sciences, 2011, 30(6): 491–507
[33] 韩利明, 张勇, 彭惠茹, 等. 从产量和品质性状的变化分析北方冬麦区小麦品种抗热性[J]. 作物学报, 2010, 36(9): 1538−1546 HAN L M, ZHANG Y, PENG H R, et al. Analysis of heat resistance for cultivars from north china winter wheat region by yield and quality traits[J]. Acta Agronomica Sinica, 2010, 36(9): 1538−1546
[34] HAYS D B, DO J H, MASON R E, et al. Heat stress induced ethylene production in developing wheat grains induces kernel abortion and increased maturation in a susceptible cultivar[J]. Plant Science, 2007, 172(6): 1113–1123
[35] VISWANATHAN C, KHANNA-CHOPRA R. Effect of heat stress on grain growth, starch synthesis and protein synthesis in grains of wheat (L.) varieties differing in grain weight stability[J]. Journal of Agronomy and Crop Science, 2001, 186(1): 1–7
[36] 赵彦坤, 王秀堂, 王静, 等. 热胁迫对不同小麦品种灌浆速率的影响[J]. 中国生态农业学报, 2016, 24(9): 1239–1245ZHAO Y K, WANG X T, WANG J, et al. Effects of heat stress on grain-filling rate of different wheat varieties[J]. Chinese Journal of Eco-Agriculture, 2016, 24(9): 1239–1245
[37] 胡吉帮, 王晨阳, 郭天财, 等. 灌浆期高温和干旱对小麦灌浆特性的影响[J]. 河南农业大学学报, 2008, 42(6): 597–601HU J B, WANG C Y, GUO T C, et al. Effects of high temperature and drought stress on grain filling characteristics in wheat during grain filling period[J]. Journal of Henan Agricultural University, 2008, 42(6): 597–601
[38] 冯素伟, 胡铁柱, 李淦, 等. 不同小麦品种籽粒灌浆特性分析[J]. 麦类作物学报, 2009, 29(4): 643–646 FENG S W, HU T Z, LI G, et al. Analysis on grain filling characteristics of different wheat varieties[J]. Journal of Triticeae Crops, 2009, 29(4): 643–646
[39] 姜雨萌, 赵风华, 刘金秋, 等. 极端高温对冬小麦冠层碳同化的影响[J]. 中国生态农业学报, 2015, 23(10): 1260–1267 JIANG Y M, ZHAO F H, LIU J Q, et al. Effect of extreme heat on winter wheat canopy carbon assimilation[J]. Chinese Journal of Eco-Agriculture, 2015, 23(10): 1260–1267
[40] 武永胜, 薛辉, 刘洋, 等. 持绿型小麦叶片衰老和叶绿素荧光特征的研究[J]. 干旱地区农业研究, 2010, 28(4): 117–122WU Y S, XUE H, LIU Y, et al. Study on senescence and chlorophyll fluorescence traits of stay-green leaf in wheat[J]. Agricultural Research in the Arid Areas, 2010, 28(4): 117–122
[41] 龚月桦, 林娜, 石慧清, 等. 持绿型小麦冠温特性及其对低氮和高温的适应性[J]. 西北农林科技大学学报: 自然科学版, 2016, 44(9): 49–55GONG Y H, LIN N, SHI H Q, et al. Canopy temperature characteristics and adaptability of stay-green wheat to low nitrogen and high temperature[J]. Journal of Northwest A&F University: Natural Science Edition, 2016, 44(9): 49–55
[42] 陈冬梅, 马永安, 苏玉环, 等. 不同落黄型小麦品种光合器官衰老及产量对花后高温的响应[J]. 麦类作物学报, 2017, 37(12): 1596–1603 CHEN D M, MA Y A, SU Y H, et al. Response of photosynthetic organ senescence and yield of wheat with different yellowing types to heat stress after anthesis[J]. Journal of Triticeae Crops, 2017, 37(12): 1596–1603
[43] 陈晓娜, 赵庚星, 周雪, 等. 基于高光谱的小麦冠层叶绿素(SPAD值)估测模型[J]. 天津农业科学, 2017, 24(2): 60–65 CHEN X N, ZHAO G X, ZHOU X, et al. Estimation model of wheat canopy chlorophyll content (SPAD value) based on hyperspectral technology[J]. Tianjin Agricultural Sciences, 2017, 24(2): 60–65
[44] 张俊华, 张佳宝, 贾科利. 不同施肥条件下夏玉米光谱特征与叶绿素含量和LAI的相关性[J]. 西北植物学报, 2008, 28(7): 1461–1467 ZHANG J H, ZHANG J B, JIA K L. Correlation between summer maize spectral reflectance and leaf chlorophyll, LAI under different fertilizations[J]. Acta Botanica Boreali-Occidentalia Sinica, 2008, 28(7): 1461–1467
[45] 王晓飞, 李志洪, 袁家萍, 等. 玉米品种冠层NDVI与叶绿素的关系[J]. 中国农学通报, 2010, 26(16): 175–179 WANG X F, LI Z H, YUAN J P, et al. The relationship between corn varieties canopy of NDVI and chlorophyll[J]. Chinese Agricultural Science Bulletin, 2010, 26(16): 175–179
Effect of high temperature stress at early grain-filling stage on plant morphology and grain yield of different heat-resistant varieties of wheat*
FENG Bo, LI Shengdong, LI Huawei, WANG Zongshuai, ZHANG Bin, WANG Fahong**, KONG Ling’an**
(Crop Research Institute, Shandong Academy of Agricultural Sciences, Jinan 250100, China)
As one of the major crops in China, wheat has direct relationship with living standard and national food security. Wheat suffers high temperature stress often at late growth period which negatively impacted wheat yield and quality. Under the steadily increasing global temperature, the frequency of high temperature stress in wheat has increased. There are many indicators used for heat resistance screening and evaluation of wheat, including chlorophyll fluorescence, cell membrane, canopy temperature, thermal index, etc, in previous researches. However, most researches were limited to laboratory analysis, and neglected yield investigation. Considering wheat as a group crop, the chlorophyll content of population canopy and NDVI were used for wheat heat-resistance evaluation in this study. The grain-filling characteristics and yield outputs were also investigated. The study aimed at providing reliable methods of breeding, and theoretical basis for cultivation of high-yielding and stress-resistant wheat varieties. In this study, the effect of high temperature stress for 3 days at early grain-filling stage on morphology and grain yield of different heat-resistant wheat varieties (lines), including heat-resistant varieties (lines) of JM22 and 056852, and heat-sensitive varieties (lines) of XM26 and GC8901, were investigated through erecting artificial greenhouse to increase on-field temperature. The highest temperature during the 3-day high temperature stress was 43.13 ℃ and the average temperatures in every day were respectively 10.48 ℃, 9.71 ℃ and 9.84 ℃ higher than that of the control. Different heat-resistance varieties (lines) varied in response of plant and grain morphologies to high temperature stress. NDVI and canopy chlorophyll content of four varieties (lines) decreased after high temperature stress. These changes of JM22 and 056852 were not significant, while NDVI values and chlorophyll contents of XM26, GC8901 significantly decreased by 9.66%, 12.10%, and 6.26%, 10.73%, respectively. High temperature stress accelerated the senescence process of wheat. The grain-filling duration were significantly shortened by 1.4 d, 0.8 d, 2.4 d and 3.0 d for JM22, 056852, XM26 and GC8901, respectively. High temperature stress significantly decreased 1000-kernel weight and grain yield of wheat. The yield reductions of heat-sensitive varieties (lines) of XM26 and GC8901 were 11.43% and 10.05%, those of heat-resistant varieties (lines) of JM22 and 056852 were 6.41% and 6.93%, respectively. In conclusion, high temperature stress at early grain-filling stage accelerated canopy chlorophyll degradation, shortened grain-filling duration, reduced grain yield of wheat. JM22 showed better heat resistant ability and yield performance. 056852 had better heat resistance ability but normal yield performance. XM26 and GC8901 were worse both in heat resistance and in grain yield.
High temperature stress; Wheat; Heat resistance; Plant morphology; Yield
WANG Fahong, E-mail:13001719601@163.com; KONG Ling’an, E-mail: kongling-an@163.com
Jun. 22, 2018;
Sep. 30, 2018
S512.1; S311
A
2096-6237(2019)03-0451-11
10.13930/j.cnki.cjea.180578
冯波, 李升东, 李华伟, 王宗帅, 张宾, 王法宏, 孔令安. 灌浆初期高温胁迫对不同耐热性小麦品种形态和产量的影响[J]. 中国生态农业学报(中英文), 2019, 27(3): 451-461
FENG B, LI S D, LI H W, WANG Z S, ZHANG B, WANG F H, KONG L A. Effect of high temperature stress at early grain-filling stage on plant morphology and grain yield of different heat-resistant varieties of wheat[J]. Chinese Journal of Eco-Agriculture, 2019, 27(3): 451-461
* 山东省农业科学院青年基金项目(2014QNM04)、公益性行业(农业)科研专项(201503130)、山东省重点研发计划项目(2017GNC11106)和山东省泰山产业领军人才高效生态农业创新类项目(LJNY201601)资助
王法宏, 主要研究方向为小麦高产高效栽培技术及生理生态, E-mail: 13001719601@163.com; 孔令安, 主要研究方向为小麦抗逆生理, E-mail: kongling-an@163.com
冯波, 主要研究方向为小麦丰产稳产节本高效栽培技术及其生理基础。E-mail: fengbo109@126.com
2018-06-22
2018-09-30
* This study was supported by the Fund for Youth Scholars of Shandong Academy of Agricultural Sciences (2014QNM04), the Special Fund for Agro-scientific Research in the Public Interest of China (201503130), the Key Science and Technology Projects of Shandong Province (2017GNC11106), the Mountain Tai Industry Leaders Innovation Projects for High Efficient and Ecological Agriculture of Shandong Province (LJNY201601).
