铁死亡研究现状*
2019-03-06葛金文余清平曾劲松吴万丰孙艺航
廖 君,葛金文,余清平,曾劲松,吴万丰,孙艺航
(湖南中医药大学医学院 湖南中医药大学中西结合学院 长沙 410208)
铁死亡是一种新的程序性细胞死亡方式[1],与铁依赖的脂质过氧化代谢有关,通过NADPH/H+、多不饱和脂肪酸代谢、谷氨酰胺分解代谢途径调节细胞死亡。铁死亡是导致神经系统退行性疾病及中风后神经元损伤的重要机制。随着铁死亡机制研究的深入,发现促进癌细胞铁死亡可以抑制癌细胞分化、迁移以及减少耐药性等功能。因此,抑制和诱导铁死亡分别是防治神经系统损伤和癌症治疗的新途径。本文主要从铁死亡机制研究进展及针对神经系统疾病和癌症中铁死亡干预研究进行文献综述。
1 铁死亡机制研究
铁是人体内重要的微量元素,参与血红素的合成,髓鞘的形成,线粒体呼吸链电子传递,DNA 复制、修复和表观遗传学控制等。细胞通过铁离子输入、输出和储存来维持铁代谢平衡,其中铁输入蛋白包括:转铁蛋白(transferrin:Tf)、转铁蛋白受体(transferrin receptor:TFR)、二价金 属离子 转运体1(DMT1,divalent metal transporter 1),铁输出蛋白包含膜铁转运蛋白1(ferroportin1:Fpn1),细胞内铁储存蛋白为铁蛋白(ferritin)[2]。人体内铁调节系统包括系统调节和细胞内调节:①系统调节是肝脏分泌产生的铁调素(hepcidin)的调节,hepcidin 能降低十二指肠铁吸收以及减少脾脏内的红细胞降解。hepcidin 同时可以诱导膜铁转运蛋白(ferroportin1:Fpn1)降解[3];②细胞调节指的是铁调节蛋白(Iron Regulatory Protein,IRP)/铁反应元素(Iron Responsive Element,IRE)系统的调节[4]。轻链铁蛋白(L-ferritin)、转铁蛋白受体(transferrin receptor:TfR)及膜铁转运蛋白(ferroportin1:Fpn1)mRNA 5’或3’端上的IRE在细胞内铁超载时与IRP结合调节细胞内铁代谢[5]。铁超载时,过量的二价铁离子通过fenton反应产生羟自由基诱导铁死亡[6]。
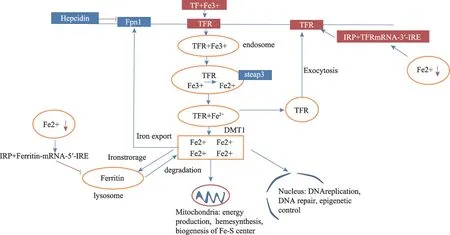
图1 细胞内铁代谢机制
铁死亡形态学上表现出细胞萎缩,线粒体高密度。半胱氨酸利用率、谷胱甘肽生物合成、多不饱和脂肪酸代谢以及磷脂的调节是铁死亡途径的关键因素。细胞膜上胱氨酸/谷氨酸逆转运体,即Xc-系统(the cystine/glutamate antiportersystem,system Xc-),介导谷胱甘肽(GSH)的生成,GSH 作为细胞内重要的抗氧化剂,是铁死亡的负调节器[7,8]。谷胱甘肽(GSH)/谷胱甘肽过氧化物酶4(GPX4)能抑制脂质氧化酶(lipoxygenase,LOX)的活性,清除铁聚集产生的脂质过氧化物,有效地抵消脂质双层脂质过氧化,防止细胞膜的损伤[9-13]。也有研究者提出铁死亡为一种细胞死亡的子程序,因缺乏GPX4 活性而被特异性激活,可通过铁螯合减少细胞内游离铁来抑制[14]。长链脂酰CoA 合成酶(ACSL)家族中的铁死亡敏感性ACSL4,可参与合成易被氧化的膜磷脂,因此提出ACSL4 参与铁死亡启动程序[15](图1)。
2 铁死亡与其他细胞死亡方式的相关性研究
铁死亡概念提出后,研究者们同时开始探讨铁死亡与其他细胞死亡方式的相关性。细胞焦亡又称细胞炎性坏死,是一种程序性细胞死亡,伴有大量促炎症因子的释放。研究者发现铁死亡早期,细胞释放炎性损伤相关因子,作用于人体免疫系统[16]。但不同于焦亡和凋亡,铁死亡不需要caspases 作为介质,也不需要作为程序性死亡中诱导线粒体通透性增加的介质Bax/Bak 的存在[17]。铁死亡与自噬相关性研究发现,铁死亡中氧化应激损伤会诱导溶酶体破裂[18,19],而自噬性死亡中吞噬泡形成后也与溶酶体结合并降解。进一步发现胰腺导管腺癌中STAT3介导组织蛋白酶B表达诱导细胞溶酶体死亡是铁死亡的重要途径[20]。肝纤维化的铁死亡过程中RNA 绑定的蛋白ELAVL1/HuR起重要作用,同时ELAVL1 上调可增加细胞内自噬体的生成及自噬的发生[21]。目前,更有研究者提出铁自噬(ferritinophagy)概念[22],铁自噬为调节细胞内铁代谢的选择性自噬。溶酶体自噬、铁释放是直接或间接参与神经退行性疾病的发病机制[23]。细胞内铁以三价铁结合形成铁蛋白储存,铁蛋白在溶酶体中降解并释放惰性的三价铁,三价铁还原为二价铁完成生理功能或促进ROS 的生成[24-26]。研究者提出核受体辅激活子4(Nuclear Receptor Coactivator 4,NCOA4)通过结合铁蛋白促进其自噬降解导致铁自噬的形成[27]。因此,有研究者们认为铁自噬是诱导铁死亡发生的关键[28]。
3 铁死亡相关疾病研究
铁死亡作为一种不同于凋亡的铁依赖脂质过氧化导致的细胞程序性死亡的方式,目前研究主要集中在铁死亡作为神经退行性变等病理机制及癌症药物治疗机制。
3.1 神经系统疾病与铁死亡相关性研究
阿尔茨海默病(Alzheimer disease,AD)是一种常见的神经退行性疾病,特征性病理改变为β淀粉样蛋白沉积及tau 蛋白过度磷酸化。临床MRI 检查发现AD 患者脑内铁含量增高,海马严重受损[29]。铁离子聚集能促进β铁淀粉样蛋白(A 粉)沉积与淀粉样斑块形成,tau 蛋白过度磷酸化与神经原纤维缠结[30]。AD 患者神经元,星型胶质细胞及小胶质细胞中铁聚集,铁调节蛋白异常,脂质过氧化物增多[31,32],AD中铁聚集诱导ROS 产物生成[33],而氧化应激参与神经毒性作用及淀粉样斑块形成和神经原纤维缠结的反应过程[34]。因此,研究者们提出AD 发病机制与铁聚集和神经元铁死亡有关。帕金森症(Parkinson's disease,PD)在神经系统退行性变中发病率排名第二,病理表现为黑质多巴胺神经元变性,目前治疗原则主要是多巴胺补充及症状控制。而针对PD 发病机制的进一步研究发现,黑质神经元内铁离子参与酪氨酸羟化酶依赖的多巴胺合成,α质突触核蛋白(α触syn)上的IRE 可以受铁离子状态调节,因此黑质中铁离子状态变化可直接干预多巴胺的表达,铁离子代谢与PD 发生具有相关性[35,36]。缺血性脑中风研究发现,神经元内铁离子聚集,细胞膜上铁输出蛋白表达减少,铁超载导致神经元损伤[87]。铁离子通过Fenton反应将超氧和过氧化氢(H2O2)转化为高活性有毒羟基自由基(OH)[37],铁螯合剂(如DFO)可改善哺乳动物缺血性卒中的预后[38]。小鼠缺血性中风动物模型中谷胱甘肽(GSH)水平显著降低,脂质过氧化增加[39],铁调蛋白抑制剂可起到保护脑缺血损伤的作用[40]。在神经系统代谢机制研究中,提出氧化还原敏感核相关因子2(Nuclear factor-E2 related factor 2,Nrf2)通过调节下游基因的表达对脑血管疾病和神经退行性疾病起到保护作用,近阶段研究发现Nrf2 能抑制铁介导的氧化应激损伤。兴奋性神经递质释放并通过调节下丘脑-垂体-肾上腺神经内分泌轴可激活星型胶质细胞生成Nrf2[41-43]。
综上,抑制神经退行性疾病及脑缺血后铁死亡可以起到神经元保护作用。实验证明ACSL4 可将长链多不饱和脂肪酸活化,参与铁死亡。而α6β4通过激活STAT3抑制ACSL4的表达,抑制铁死亡[44]。铁螯合剂,铁抑素,抗氧化剂治疗均可有效抑制铁死亡[45]。
3.2 癌症治疗的铁死亡机制研究
铁是有利于细胞增殖和生长的必要营养,同时参与氧化还原反应和自由基生成。铁参与与癌症的发生和生长,癌症微环境的调节及癌细胞转移[46]。铁死亡分子机制的研究将有助于打破抑制和诱导细胞死亡之间的平衡。近年来研究者们越来越多的关注诱导癌细胞铁死亡的研究[47]。药物诱导铁死亡主要通过:①抑制胱氨酸/谷氨酸逆转运体(Xc-系统)[48,49];②抑制或消耗GPX4 的表达[50];③高表达P53,诱导铁死亡。
3.2.1 抑制胱氨酸/谷氨酸逆转运体诱导铁死亡。
临床常用的铁死亡诱导剂erastin 是通过抑制胱氨酸/谷氨酸逆转运体,减少胱氨酸输入,进而GSH 合成减少,ROS 聚集[51]。erastin 和sorafenib 都是Xc-系统抑制剂,但erastin 可以激活STAT3,而sorafenib 抑制STAT3 活性。sorafenib 是诱导铁死亡治疗肝细胞癌的一线药物[52],针对胰腺导管腺癌、肝癌研究中,sorafenib通过抑制STAT3 活性促进癌细胞铁死亡增强抗癌活性[53,54]。研 究表明MAPK/ERK 是erastin 诱导STAT3 激活的上游信号通路[55]。Sulfasalazine 可以通过抑制Xc-系统治疗头颈部癌。Sulfasalazine 治疗中出现耐药的分子机制研究中发现CISD2 基因过表达,而抑制CISD2 表达后,Sulfasalazine 恢复诱导铁死亡[56]。肿瘤抑制剂BAP1 通过调节胱氨酸转运体SLC7A11 的表达来调控癌细胞的生长[57]。BAP1抑制SLC7A11的表达,减少胱氨酸吸收,导致脂质过氧化物聚集及癌细胞铁死亡[58,59]。针对铁死亡探讨消化系统其他癌症胃癌、结肠癌治疗机制的研究也正在展开[60-62]。
3.2.2 GPX4是铁死亡过程中的关键调节器。
恶性B 细胞淋巴瘤研究中抗氧化酶GPX4 负表达与对照组比较具有较高的总生存率,GPX4 基因敲除细胞易发生ROS 诱导的铁死亡。因此研究者提出诱导铁死亡可以作为癌症新的治疗方法,并建议将GPX4 作为恶性B 细胞淋巴瘤的评价预后的指标[63]。RSL3 可直接结合和抑制谷胱甘肽过氧化物酶glutathione peroxidase 4(GPX4),减少脂质过氧化物的产生,因此,RSL3可视为铁死亡诱导剂[64,65]。但也有实验提出消耗GPX4 诱导的细胞死亡时间为72 h,RSL3药物干预导致的铁死亡时间点是12 h。提出RSL3 诱导的铁死亡机制可能不是因为降低GPX4[66]。基质通过α质通过整合素维持GPX4 的表达,激活PVRL4/是降低是降Src 信号通路抑制脂质过氧化。而与基质分离的癌细胞α质分离的缺失,具有铁死亡易感性[67]。
3.2.3 p53 作为基因组卫士,参与了各种应激条件下的细胞生存和分裂的调节
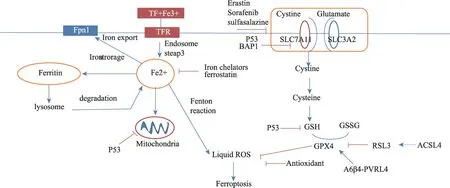
图2 铁死亡及药物干预机制
p53 基因是一种抑癌基因,是细胞生长周期中的负调节因子,与细胞周期的调控、DNA 修复、细胞分化、凋亡等重要的生物学功能有关。p53 通过泛素化、磷酸化、乙酰化等调节细胞功能[68,69]。低水平p53 可诱导细胞周期停滞,DNA 修复,通过下调活性氧抑制DNA 的氧化应激损伤[70]。高水平细胞质内p53 直接结合BCL-2,增加线粒体膜通透性,诱导细胞凋亡[71]。近年来,研究表明p53是铁死亡的调节因素。p53可通过干预单核苷酸多态性、长链非编码RNA 和SOCS1(细胞因子信号抑制物)在铁死亡中表现出多种生物化学特性,是铁死亡和细胞凋亡调节的重要途径[72,73,74,75,76]。p53 通过抑 制SLC7A11,增 加SAT1(spermidine/spermine N1-acetyltransferase 1)和GLS2(glutaminase 2)的表达,来促进铁死亡[77]。长链非编码RNA(Long noncoding RNAs,IncRNA)与很多癌症密切相关,研究发现细胞质中p53 相关的lncRNA 称p53RRA 可以作为癌症发展的抑制剂。胞浆P53RRA-G3BP1 相互作用从G3BP1复合物中替换p53,导致大量p53滞留在细胞核里,诱导凋亡和铁死亡[78]。土槿皮乙酸(PAB)是土槿皮科树根及干皮的提取物,PAB 上调转铁蛋白受体(transferrin receptor)的表达,通过p53 介导消耗GSH,增加H2O2和脂质过氧化物,通过铁死亡治疗胶质瘤[79](图2)。
4 中药干预铁死亡研究
中药干预铁死亡的探索还处于起步阶段,但展现出不凡的研究前景。小鼠间充质干细胞高铁离子环境下培养,出现细胞低存活和低增殖,及线粒体活性氧增多。黄芪多糖则可明显抑制线粒体活性氧的产生,保护间充质干细胞[80]。黄芪黄酮内的有效成分Formononetin 具有抗氧化应激的作用,并可抑制抗癌药物cisplatin 的细胞毒性等副作用[81]。右旋糖酐铁注射液注射7 周造模慢性铁超载肝细胞纤维化模型小鼠,注射铁剂前4 h 加丹参注射剂干预,可明显增加抗氧化剂谷胱甘肽(GSH)及超氧化物歧化酶(SOD)的表达[82]。蒿甲醚(ART)是青蒿素衍生物,是治疗疟疾的一线药。近段研究发现,蒿甲醚通过抑制肝星状细胞(HSC)活化从而抑制肝硬化铁死亡[83]。肝癌和胰腺癌研究中黄芩素通过(Keap1)-Nrf2 通路抑制铁死亡,青蒿素可调节铁相关基因,通过促进ROS 聚集诱导CCRF-CEM 白血病细胞铁死亡[84,85]。药物干预小鼠AD 模型,普鲁士蓝染色检测海马铁分布及水迷宫检测小鼠学习记忆指标,发现淫羊藿、黄芪、葛根三种中药具有抑制铁聚集,改善学习记忆能力的功效[86]。中药复方脑泰方通过促进脑缺血后铁输出蛋白Fpn的表达,抑制铁聚集,保护神经元[87]。
5 总结
随着铁死亡概念的提出,研究者们在这个领域做了逐步深入的机制探讨。提出了铁死亡是铁依赖的脂质过氧化物生成的细胞程序性死亡方式。抑制神经系统退行性疾病及脑缺血铁死亡起到神经元保护作用,大量铁离子聚集导致脂质过氧化物增加,诱导癌细胞铁死亡是目前癌症治疗新的有效方式。因此铁死亡的提出既促进了疾病发生的病理生理机制研究,同时为相关疾病治疗提供了新的靶点。目前铁死亡的研究集中提出Xc-系统、谷胱甘肽及谷胱甘肽过氧化物酶在调节铁死亡中起着重要作用。那么除脂质过氧化物产生是导致细胞铁死亡的主要原因外,氧自由基反应及细胞铁代谢调节(铁输入、铁输出、铁储存、铁离子细胞器内的代谢)的关键机制,及诱导铁死亡的相关性需要进一步探讨。
