外源Ca2+提高百合耐热性的生理机制初探
2019-03-06,,,,,
,,, ,,
(1.聊城大学 农学院,山东 聊城 252059; 2.南京农业大学 园艺学院,江苏 南京 210095;3.中国农业大学 园艺学院/花卉发育与品质调控北京市重点实验室,北京 100193)
百合(Liliumspp.)为百合科百合属球根花卉,具有重要的观赏、食药用价值。百合性喜冷凉、湿润气候,耐热性较差,高温造成百合生长停滞、植株矮小、少花盲花、茎秆软、病虫害严重等现象,严重影响切花的产量和质量,并造成种球退化。我国大部分地区夏季炎热,严重影响百合商品化周年生产[1]。所以,提高百合耐热性是解决这一问题的关键。
作为植物生长发育所必需的大量元素,Ca2+是细胞壁的组成成分,是细胞膜的稳定剂,也是细胞内多种酶的活化剂,同时作为第二信使,Ca2+与其受体蛋白如钙调蛋白(CaM)、类钙调磷酸酶B亚基蛋白(CBL)、钙离子结合蛋白激酶(CDPK)等结合,参与了植物对内环境(发育)和外环境的感知和传导,涉及植物对病原菌、光、高低温、机械损伤、干旱、盐和渗透胁迫等的响应[2]。施用外源Ca2+能提高植物的耐热性。10 mmol/L Ca2+处理能够有效缓解高温强光对西葫芦光合机构的伤害[3]。10 mmol/L CaCl2处理能提高滨梅幼苗对高温的耐受性,Ca2+信号系统参与了胁迫过程中渗透物质和Ca2+-ATPase活性等的调节[4]。
利用Ca2+、Ca2+载体A23187,钙离子螯合剂EGTA,钙离子通道阻抑剂LaCl3、异搏定(Verapamil),钙调蛋白抑制剂氯丙嗪(Chlorpromazine, CPZ)、三氟拉嗪(Trifluoperazine, TFP)、N-(6-氨基己烷基)-5-氯-1-萘-黄胺(W7)等增强或阻碍Ca2+-CaM信号传导是目前研究植物Ca2+-CaM信号功能的重要手段[5]。以东方百合杂种系(Liliumoriental hybrids)品种西伯利亚(Siberia)为试验材料,研究CaCl2、EGTA和TFP对高温胁迫下百合表型、相关生理指标及基因表达的影响,探讨Ca2+在百合耐热性调节方面的功能及作用机制,以期为生产上应用Ca2+提高百合耐热性奠定理论与技术基础。
1 材料和方法
1.1 试验材料
试验材料为东方百合杂种系西伯利亚,每盆1株。栽培基质配比为V(草炭)∶V(细河砂)=2∶1。人工气候室培养温度为22 ℃/16 ℃(昼/夜),光照时间为14 h/d。CaCl2和EGTA购于中国国药集团,TFP购于Sigma公司,荧光定量试剂盒SYBR®Premix ExTaq购自TaKaRa公司。
1.2 试验方法
1.2.1 热害指数的测定 待百合地上部分长至约30 cm时,选取健壮、生长一致的植株,分别用5、10、15 mmol/L CaCl2,5、10、15 mmol/L EGTA和100、200、300 μmol/L TFP进行灌根处理,以去离子水处理为对照,每5 d处理1次,共处理3次。每个处理6株苗,重复3次。第3次药剂处理后立即将植株置于光照培养箱内,40 ℃胁迫处理72 h,光照时间为14 h/d。热激处理后,取植株中部叶片(株间相近叶位),利用Photoshop CS5软件参照于守超等[6]的方法测定叶面积,统计伤害面积率,计算热害指数。热害分级标准见表1。
伤害面积率=叶片明显失绿部分面积/叶片总面积×100%
热害指数= ∑(各级热害叶数×相应级数值)/(调查总叶数×最高级别数值)×100。

表1 热害分级标准
1.2.2 相关生理指标的测定 人工气候室培养条件下,分别用15 mmol/L CaCl2、10 mmol/L EGTA和200 μmol/L TFP溶液灌根,以去离子水处理为对照,每5 d处理1次,共处理3次。每个处理15株苗,重复3次。第3次药剂处理后立即将植株置于光照培养箱内,40 ℃胁迫处理72 h,光照时间为14 h/d,分别在热激处理0、12、24、48、72 h取植株中部叶片(株间相近叶位),测定相关生理指标。膜透性测定采用电导仪法,叶绿素含量测定采用浸提法,可溶性蛋白含量测定采用考马斯亮蓝G-250法,脯氨酸含量测定采用酸性茚三酮法,丙二醛(MDA)含量测定采用硫代巴比妥酸法,超氧化物歧化酶(SOD)活性测定采用氮蓝四唑显色法。以上指标的测定均参照李合生[7]的方法。
1.2.3 相关热响应基因的表达分析 人工气候室培养条件下,分别用15 mmol/L CaCl2、10 mmol/L EGTA和200 μmol/L TFP溶液灌根,以去离子水处理为对照,每5 d处理1次,共处理3次。第3次药剂处理后立即将植株置于光照培养箱内,40 ℃热激1 h和3 h,取植株中部叶片(株间相近叶位),用液氮速冻后放入-80 ℃超低温冰箱中保存。采用实时荧光定量PCR(Real-time quantitative RT-PCR,qPCR)方法测定样本中钙调蛋白基因LoCaM3和热激转录因子基因LoHsfA3a的表达量,以百合18SrRNA作内参。试验参照SYBR®Premix ExTaq试剂盒说明书进行,仪器为ABI StepOnePlusTM。LoCaM3的扩增引物为qF1(5′-CATGTCATGACTAACCTAGGCGAG-3′)和qR1(5′-CCATTAGAAATCAGCCAGCACC-3′),LoHsfA3a的扩增引物为qF2(5′-CTTGGTTTAAGTACGCCAGTGGAAG-3′)和qR2(5′-GTAAAATATTGTAAAAGAACATGAAGCCTATGG-3′),18SrRNA的扩增引物为18SqF(5′-AGTTGGTGGAGCGATTTGTCT-3′)和18SqR(5′-CCTGTTATTGCCTCAAACTTCC-3′)。反应程序为:95 ℃ 30 s;95 ℃ 5 s,58 ℃ 30 s,72 ℃ 20 s,40个循环。采用2-ΔΔCT方法[8]分析基因的相对表达量,3次生物学重复。
1.3 数据处理
用Excel 2007和SPSS 18处理分析数据。
2 结果与分析
2.1 Ca2+处理对高温胁迫下百合叶片热害指数的影响
热激处理对叶片造成了不同程度的伤害,CaCl2、EGTA和TFP处理对百合叶片热害指数的影响见图1。结果表明,与对照相比,CaCl2处理显著降低了叶片的热害指数(P<0.05),随着CaCl2浓度的增加,叶片的热害指数逐渐降低,15 mmol/L CaCl2处理的叶片热害指数比对照降低了58.53%。EGTA和TFP处理则显著提高了叶片的热害指数(P<0.05),热害指数随着处理浓度的增加而升高。与对照相比,10 mmol/L EGTA处理的热害指数增加了56.11%,但与15 mmol/L EGTA处理差异不显著。200 μmol/L TFP 处理的热害指数比对照增加了69.94%,但与300 μmol/L TFP处理差异不显著。根据表型结果,选择15 mmol/L CaCl2、10 mmol/L EGTA 和200 μmol/L TFP处理用于后续的生理试验。

不同小写字母表示在0.05水平上差异显著,下同
2.2 Ca2+处理对高温胁迫下百合叶片抗性相关生理指标的影响
2.2.1 细胞膜透性 高温破坏细胞膜的结构,导致细胞膜的透性增加,电解质外渗。因此,细胞膜电解质渗透率能反映出高温对植物伤害的程度。由图2A可以看出,随着热激时间的延长,百合叶片的相对电导率逐渐增大。Ca2+处理的相对电导率始终低于对照,表明Ca2+处理能够缓解高温下细胞膜透性的增大。EGTA和TFP处理的相对电导率始终高于对照,表明EGTA和TFP处理加剧了高温下细胞膜透性的增大。
2.2.2 叶绿素含量 高温胁迫过程中,百合叶片的叶绿素含量逐渐降低(图2B),表明高温抑制了叶绿素的合成,促进了叶绿素的降解。与对照相比,Ca2+处理的叶绿素含量下降趋势较缓,热激72 h后,降幅为对照的69.88%,叶绿素含量是对照的1.56倍;而EGTA和TFP处理则加剧了叶绿素含量的下降,热激72 h后,降幅分别为对照的1.22倍和1.31倍,叶绿素含量分别是对照的58.15%和42.55%。
2.2.3 脯氨酸和可溶性蛋白含量 植物细胞可以通过渗透调节的方式抵御高温胁迫,脯氨酸是最有效的渗透调节物质之一,可以防止植物水分散失和提高原生质胶体的稳定性。图2C表明,热胁迫前期(0~24 h),EGTA和TFP处理的脯氨酸含量升高,但绝对含量和增幅均显著低于对照(P<0.05)。随后,EGTA处理的脯氨酸含量变化不大,而TFP处理的脯氨酸含量降低后又迅速升高,72 h的含量接近对照水平。高温胁迫过程中Ca2+处理和对照的脯氨酸含量呈现先升高后降低的趋势,在48 h达到峰值,Ca2+处理的脯氨酸含量分别比对照、EGTA和TFP处理增加了27.24%、117.37%和165.80%。
可溶性蛋白是重要的渗透调节物质和营养物质,其增加和积累能提高细胞的保水能力,也对细胞的生命物质及生物膜起到保护作用。高温胁迫过程中,各处理的可溶性蛋白含量先升高后降低(图2D),前期升高可能是由于高温诱导合成了热激蛋白(HSP),后期降低可能是持续高温导致蛋白质合成受阻和降解加剧。热胁迫期间,Ca2+处理的可溶性蛋白含量始终高于对照,EGTA和TFP处理的可溶性蛋白含量始终低于对照。各处理的可溶性蛋白含量差异在48 h达到最大,Ca2+处理是对照的1.37倍,EGTA和TFP处理分别为对照的73.81%和52.86%。
2.2.4 MDA含量和SOD活性 高温胁迫诱导细胞积累过多活性氧,引起或加剧膜脂过氧化作用,MDA是产物之一,其含量的高低可以反映膜的损伤程度。高温胁迫期间,百合叶片MDA含量的变化趋势如图2E所示,各处理的MDA含量持续增加。热激12 h,各处理的MDA含量无显著差异(P>0.05)。热激24、48、72 h,EGTA处理的MDA含量分别是对照的1.64、1.34、1.21倍,TFP处理的MDA含量分别是对照的1.88、1.61、1.34倍,Ca2+处理的MDA含量分别是对照的67.85%、62.21%和77.83%。
虽然高温下产生的自由基会伤害细胞,然而植物体也有防御系统,SOD 是植物抗氧化防御系统中的保护酶之一,能清除高温胁迫下细胞内产生的过多活性氧,以减轻活性氧对细胞的伤害。Ca2+、EGTA和TFP处理对高温胁迫下百合叶片SOD活性的影响见图2F。高温胁迫期间,各处理的SOD活性呈先升高后降低的趋势。与对照相比,Ca2+处理显著提高了SOD活性(P<0.05),差异随胁迫时间的延长逐渐变大,至72 h,SOD活性为对照的2.15倍。EGTA和TFP处理的SOD活性在热胁迫初期(0~12 h)小幅增强后持续降低,至72 h,SOD活性分别为对照的36.14%和26.25%,表明EGTA和TFP处理显著抑制了SOD活性(P<0.05)。
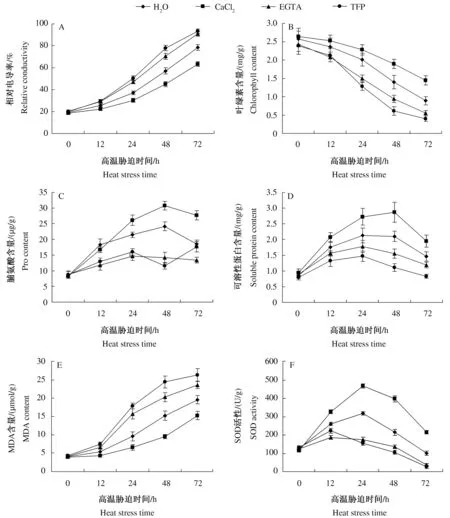
图2 高温胁迫下不同药剂处理对百合叶片抗性相关生理指标的影响Fig.2 Effect of different chemicals on correlative physiological indexes of lily leaves under heat stress
2.3 Ca2+处理对高温胁迫下百合LoCaM3和LoHsfA3a表达的影响
CaM3与HsfA3a参与了百合的热激信号转导[9-10]。40 ℃热胁迫条件下,检测了CaCl2、EGTA、TFP处理1 h后叶片LoCaM3的表达量和处理3 h后叶片LoHsfA3a的表达量(图3)。结果表明,与对照相比,CaCl2处理显著促进了LoCaM3和LoHsfA3a的表达(P<0.05),而EGTA或TFP处理则显著抑制了LoCaM3和LoHsfA3a的表达(P<0.05),表明外源Ca2+可以通过Ca2+/CaM-HSF信号途径参与百合的热激反应。
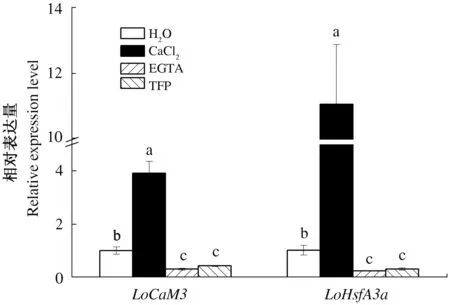
图3 高温胁迫下不同药剂处理的百合叶片LoCaM3和LoHsfA3a的表达量Fig.3 Expression of LoCaM3 and LoHsfA3a of leaves treatment with different chemicals under heat stress
3 结论与讨论
目前,有关百合的耐热研究主要集中在以下2个方面:第一,百合对高温胁迫的形态和生理响应。东方百合Sorbonne、岷江百合等在高温环境下叶片逐渐萎蔫、变黄,随着温度的升高和处理时间的延长,植株的耐热指数不断下降[11-12]。本研究中,西伯利亚百合在高温胁迫下的表型变化与上述研究基本一致,40 ℃热激72 h后,叶片基本都萎蔫下垂,因此,根据叶片的失绿程度进行热害分级,能够客观反映植株的受害程度。与对照相比,5~15 mmol/L CaCl2处理显著降低了热害指数,5~15 mmol/L EGTA和100~300 μmol/L TFP处理则显著提高了热害指数。高温处理的百合叶片脯氨酸、MDA含量和相对电导率显著升高,这3个指标可作为耐热性鉴定的生理指标[13]。高温胁迫下,百合的过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸过氧化物酶(APX)、SOD等抗氧化酶的活性和谷胱甘肽(GSH)、抗坏血酸(AsA)等抗氧化剂的含量处于较高水平,可以有效清除高温诱导产生的活性氧,提高百合的耐热性[14]。本试验中,热胁迫导致相对电导率和MDA含量升高,脯氨酸、可溶性蛋白含量和SOD活性先升高后降低,叶绿素含量下降,这与杨炜茹等[12]对岷江百合的研究结果基本一致。第二,提高百合耐热性的途径和方法。利用物理或化学的方法可以提高百合的耐热性。热激锻炼百合种球可以提高植株SOD活性、可溶性糖含量和根系活力[15],可以引起植株体内SOD与光合色素的协同作用并提高系统的热耗散[16],能抑制由高温胁迫引起的相对电导率的升高[17],从而提高百合的耐热性。外源水杨酸可以通过提高抗氧化系统酶的活性来增强百合的耐热性[18]。外施Ca2+可以提高小麦[19]、半夏[20]、藤本月季[21]、高山杜鹃[22]等植物的耐热性,之前的研究表明,外源Ca2+处理可以显著提高麝香百合品种白天堂组培苗的耐热性[9],但未深入研究其生理机制。本研究结果表明,与单纯高温处理相比,15 mmol/L CaCl2处理能够降低电解质渗透率和MDA含量,提高叶绿素、脯氨酸、可溶性蛋白含量和SOD活性,即通过降低细胞膜透性和膜脂过氧化作用,提高光合色素、渗透调节物质含量和抗氧化酶活性,增强植株的耐热性;10 mmol/L EGTA和200 μmol/L TFP处理则加剧了电解质的外渗和MDA的积累,降低了叶绿素、脯氨酸、可溶性蛋白含量以及SOD活性,从而降低植株的耐热性。在对茄子[23]、滨梅[4]、棉花[24]等的研究中也得到了类似的结果,表明作为植物生长发育所必需的大量元素,Ca2+也可能通过Ca2+-CaM信号途径介导百合对高温胁迫的响应。筛选耐热种质是培育耐热品种的基础[25-26],杂交育种是提高百合耐热性的主要途径[27-28]。近年来,百合耐热转基因育种也取得了初步进展。尚爱芹等[29]利用农杆菌介导的方法将拟南芥干旱应答元件结合蛋白AtDREB2A基因转化百合,高温胁迫下,转基因植株的POD、SOD活性等及脯氨酸含量高于野生型植株,耐热性明显强于野生型植株。从百合中鉴定了一些耐热功能基因,如LlHsfA1[30]、LlHsfA2a[31]、LlHsfA2b[32]、LlHsfA3a[10]、LlHsfA3b[10]、LlHSP70[33]、LimHSP16.45[34]等。前期对麝香百合的研究中发现,LlCaM3为响应Ca2+和热胁迫的重要信号分子,Ca2+/LlCaM3可能在A类HSF的上游发挥作用[9];LlHsfA3a的表达受热胁迫诱导,转基因拟南芥中一些HSP基因的表达量显著增加[10]。在本研究中,检测了CaCl2、EGTA、TFP处理对高温胁迫下东方百合LoCaM3和LoHsfA3a表达的影响,结果表明,CaCl2处理显著促进了LoCaM3和LoHsfA3a的表达,而EGTA或TFP处理则显著抑制了LoCaM3和LoHsfA3a的表达,说明外源Ca2+可能通过Ca2+/CaM-HSF信号途径诱导HSP的合成,利用HSP的分子伴侣作用[35],提高百合的耐热性。
15 mmol/L CaCl2处理能够缓解由高温胁迫造成的膜透性的加大和MDA的积累,提高叶绿素、脯氨酸、可溶性蛋白含量和SOD活性,促进热响应基因LoCaM3和LoHsfA3a的表达,表明Ca2+作为植物生长所必需的大量元素和偶联胞外信号与胞内生理反应的第二信使,能通过提高细胞的光合能力、渗透调节能力、抗氧化能力和蛋白质活性来维持细胞壁、细胞膜及蛋白质的稳定性,从而增强百合对高温胁迫的抗性。利用Ca2+提高栽培作物的抗逆性具有成本低、对环境影响小的突出优势,本研究初步探讨了Ca2+对百合耐热性的影响及其生理机制,为生产上应用Ca2+提高百合耐热性奠定了理论与技术基础。
