庐山毛竹扩张及模拟氮沉降对土壤N2O和CO2排放的影响*
2019-02-26刘苑秋王翰琨邓邦良刘晓君董星宇令郑
李 超 刘苑秋 王翰琨 陈 琦 邓邦良 刘晓君 董星宇 张 令郑 翔 曹 雯
(江西农业大学林学院,鄱阳湖流域森林生态系统保护与修复国家林业局重点实验室(江西农业大学),江西省森林培育重点实验室,南昌 330045)
近一个世纪以来,全球气候变暖引发了各界对温室气体排放及其潜在风险的广泛关注。氧化亚氮(N2O)、二氧化碳(CO2)是与全球气候变化密切相关的2种重要的温室气体,探索其源汇及地域排放特征也成为目前全球气候变化研究领域的重要内容。森林土壤是与N2O及CO2排放相关的重要碳、氮库[1]。大量研究表明,森林土壤的C、N特征因植物种类组成的变化[2]和植被类型[3]的转变而改变[4-5],而土壤的C、N特征和环境条件的改变会进一步影响土壤微生物群落[6]。由于土壤温室气体排放受到C、N底物和微生物群落的控制,因此土壤C、N状态和植被的群落变化可能会改变土壤的温室气体排放。
目前,源于人类活动的外源氮输入大约比源于陆地生态系统的氮输入高30%~50%[7],人类活动加速了全球活性氮的排放和固定。1961—2010年的50年内,中国陆地大气氮沉降速率由0.29 g·m-2·a-1增长至 2.32 g·m-2·a-1,增加了近8倍[8],目前中国已成为氮沉降最严重的区域之一。研究表明,森林中外源氮的输入对森林生态系统的C、N循环过程、土壤微生物群落特征及生态系统的稳定性产生了重要影响[9]。
毛竹(Phyllostachys edulis)属禾本科竹亚科刚竹属,是一种高大散生乔木状克隆植物,广泛分布于我国亚热带地区。毛竹属于典型的无性系繁殖,其营养生长过程(即克隆生长)使其具有相当大的水平拓展能力,依靠强大的地下茎(竹鞭)不断向邻近的异质小生境蔓延,实现种群克隆扩张,形成混交林甚至毛竹纯林。毛竹强大的入侵性对周边植被构成了潜在的威胁。因此,近期的研究已将毛竹视为亚热带地区一种潜在的入侵物种[10-12]。
目前,关于毛竹扩张的研究主要集中于其扩张对阔叶林的根系[13]、土壤物理性质[14]、土壤碳[15]、氮[16]、磷[12]特征以及对生态系统生物多样性[17]、土壤动物[18]、土壤微生物[15]的影响等方面,而对于在氮沉降背景下其扩张及对土壤温室气体排放造成的影响研究甚少,且无原位观测数据。本研究采用静态箱-气相色谱法,在江西庐山自然保护区选择毛竹林扩张严重的日本柳杉(Cryptomeria japonica)林为研究对象,分析比较毛竹纯林、毛竹扩张形成的毛竹-日本柳杉混交林及日本柳杉纯林的土壤N2O和CO2排放速率及累积排放量,同时通过人工模拟氮沉降试验,探究3种林分土壤N2O和CO2排放速率及累积排放量对氮沉降的响应,为毛竹扩张的生态影响研究提供理论基础。
1 材料与方法
1.1 研究区概况
庐山自然保护区位于江西省九江市(115°51′—116°07′ E,29°30′—29°41′ N),总面积约30.2 km2。气候属于亚热带季风气候,年均降水量为2070 mm,年均温度11.6℃,年均雾日191 d,霜期150 d[19]。庐山的毛竹林以前只生长在600 m左右的低海拔地区,但是,目前已经扩张到海拔较高(高达950 m)的曾被阔叶林生长覆盖的地区。区内毛竹面积约4 142 hm2,占总面积的13.12%,近十几年来,毛竹的数量估计增加了230万余株[19]。
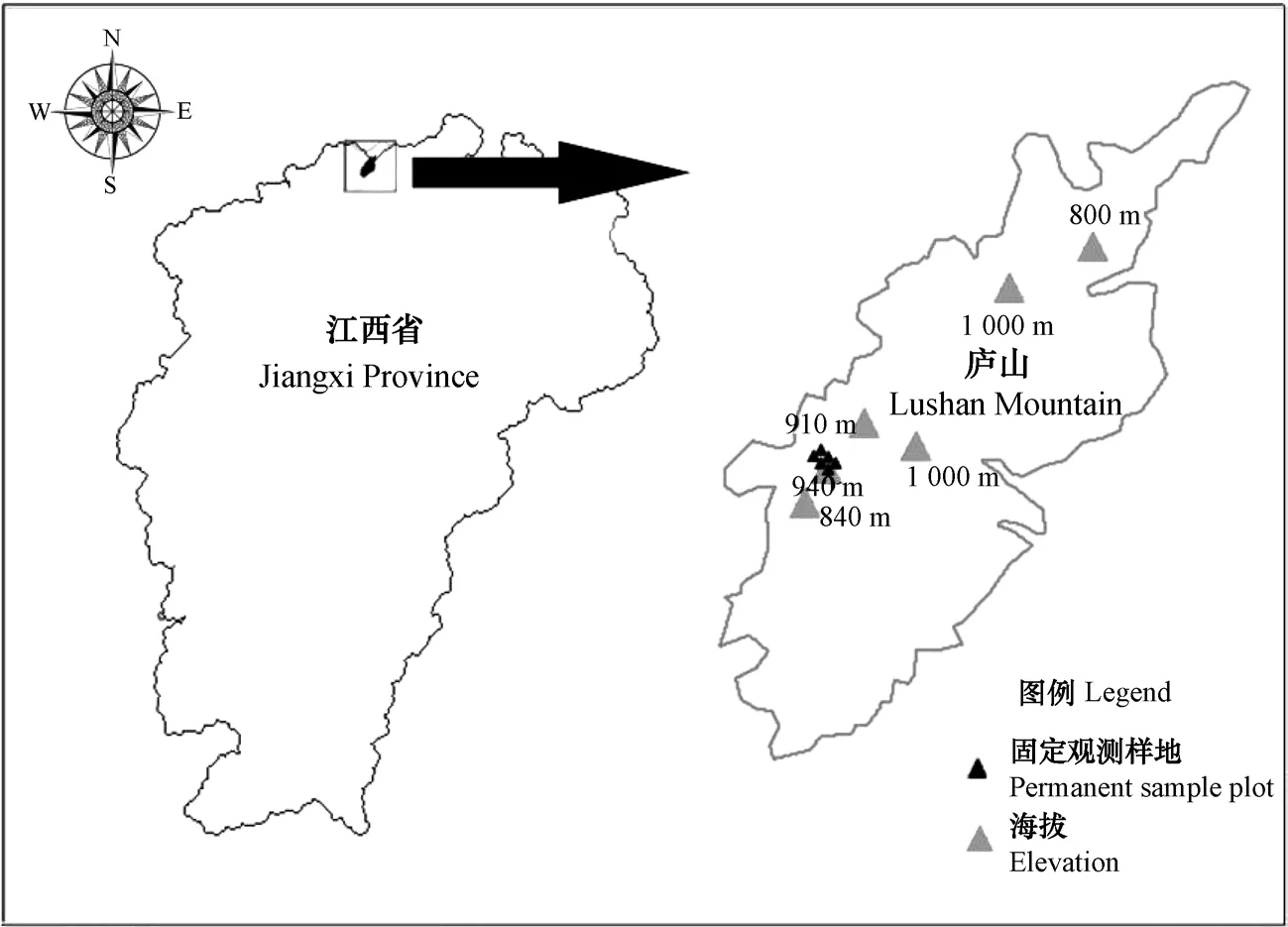
图1 试验样地区位图Fig.1 Distribution of the sample sites
本试验地点位于海拔940~960 m的金竹坪,试验设置3个林分样地:(1)毛竹纯林,林样地土壤为山地黄壤,成土母质主要为石英砂岩风化的残积物和坡积物,土层深度为40~60 cm。(2)毛竹-日本柳杉混交林(以下称“混交林”,形成于2010年前后),样地土壤为山地黄棕壤,土质较疏松,土层深度为60~90 cm。(3)日本柳杉纯林,样地土壤为山地黄棕壤,土质较疏松,土层深度为60~90 cm。三种样地均林下灌草较少,覆盖率低,零散分布的植物有中国绣球(Hydrangea chinensis)、华南毛蕨(Cyclosorus paresiticus)、山麦冬(Liriope spicata)、山橿(Lindera reflexa)、凹叶厚朴(Magnolia officinalis)、淡竹叶(Lophatherum gracile)、老鼠矢(Symplocos stellaris)、鸡爪槭(Acer palmatum)、斑叶兰(Goodyeraschlechtendaliana)、毛叶石楠(Photinia villosa)等。
1.2 研究方法
2017年4月,在选定的3种林分类型中,每种林分类型各随机设置3个面积为30 m×30 m的样地,共9个样地。每个样地设置空白对照和氮沉降两种处理方式。
采用静态箱-气相色谱法测定土壤N2O和CO2通量。静态箱体由箱体和底座两部分组成,箱体为内径20 cm,高80 cm的PVC管制成,底座为内径18.7 cm、外径23.5 cm、高10 cm、壁厚1 cm的PVC圆环凹槽。在选定的每个样地中分别随机布置2个PVC底座(模拟氮沉降和空白对照两种处理),底座埋入土壤6~8 cm深,埋好后去除环内的活体植物及凋落物,上方覆盖一层孔眼大小为1 mm× 1 mm的纱网,防止凋落物掉落其中。本研究未包含完整植物的自养呼吸,因此未分析生态系统净碳收支[20]。分别在5月和9月对各个样地中一个底座内及周围1 m2用手提式喷雾器均匀喷洒尿素(CO(NH2)2)溶液进行氮沉降处理,每次施氮水平为N 4 g·m-2,在另一个底座内及周围喷洒相同质量的蒸馏水作为空白对照。
2017年4—11月毛竹生长季进行气体采集,频率为每月一次。施氮后加密采样,平均2 d采集一次,持续两周,通量平稳后恢复每月一次。为避免天气因素影响,选择晴朗、无风天气采集气体。采气时先在底座内加入适量水保证采气时的密封性,后将采气箱扣放在底座凹槽中,并用60 mL塑料注射器在扣箱后0、5、10、15 min分别抽取40 mL气体。每次抽取气体时来回拉动注射器使箱内气体充分混合,每个点共采集4个气样,采集的气样储存在专用的气袋(德霖铝箔采气袋,大连)中,迅速带回实验室,使用气相色谱仪(Agilent 7890B,USA)分析,确定每个样品的N2O和CO2浓度。采气期间同时记录土壤温度(10 cm)、土壤湿度(10 cm)及大气温度。
气体样品采集同时,在静态箱周围多点采集0~10 cm深土壤样品后混合均匀,同时在样地中采集新鲜的毛竹凋落物、日本柳杉凋落物。土壤样品去除石砾、根系等杂物,用冰袋运回实验室用于测定其可溶性有机碳(DOC)、铵态氮()、硝态氮()、全氮(TN)含量和pH。凋落物风干后研磨过筛,测定有机碳(OC)、全氮(TN)含量。相应指标的样品处理及测定方法参照文献[21]。
1.3 数据分析
土壤N2O和CO2排放通量采用如下方程式计算:
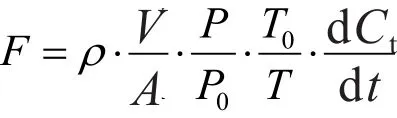
式中,F表示气体排放通量(单位为mg·m-2·h-1,通量为正则说明土壤排放该气体,为负则为吸收该气体;ρ为标准状态下被测气体的密度(mg·m-3);V和A分别为静态箱体体积(m3)和箱体底面积(m2);P和P0分别为观测时的大气压和标准大气压(KPa);T和T0分别为观测时的箱内温度和标准状态温度(℃);dCt/ dt为单位时间内箱内气体浓度随时间变化的速率(m3·m-3·h-1)。
土壤N2O和CO2累积排放量计算公式如下:
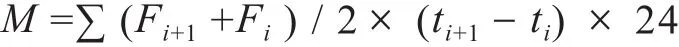
式中,M表示气体累积排放量(CO2气体累积排放量单位为mg·m-2,N2O为μg·m-2),F表示气体排放通量(CO2气体通量单位为mg·m-2·h-1,N2O为μg·m-2·h-1),(Fi+1+ Fi)表示为同一采样点连续两次气体排放通量之和,i表示第i次采集气体样品,(ti+1- ti)为连续两次采集样品期间所隔天数。
采用SPSS 19.0进行数据分析,Origin 8.1作图。
采用单因素方差分析(ANOVA)和多重比较(LSD Duncan)检验不同林分类型之间土壤DOC、TN、、、pH及凋落物OC、TN等理化性质的差异性。以林分类型和模拟氮沉降为固定效应,试验时间为随机效应,采用混合模型方差分析,分析林分类型、模拟氮沉降以及二者的交互作用对土壤N2O和CO2排放速率的影响。运用二因素方差分析(two-way ANOVAs)分析林分类型和模拟氮沉降对土壤N2O和CO2累积排放量的影响,所有差异性检验水平α = 0.05。
2 结 果
2.1 不同林分类型的土壤理化性质
从表1看出,不同的林分类型对土壤的化学性质产生了不同程度的影响。毛竹纯林与日本柳杉纯林的土壤DOC含量无显著差异(P>0.05),但混交林与其他两个林分类型间差异显著(P<0.05),且低于其他两个林分类型,说明在毛竹扩张进入日本柳杉纯林的过程中,土壤的DOC含量有降低的趋势。日本柳杉纯林与混交林土壤TN含量之间无显著差异(P>0.05),毛竹纯林土壤TN含量显著低于其他两种林分类型(P<0.05)。混交林与毛竹纯林土壤NH4+-N含量及pH均无显著差异(P>0.05),日本柳杉纯林土壤NH4+-N含量及pH显著低于其他两种林分类型(P<0.05)。毛竹纯林与日本柳杉纯林间土壤NO3--N含量无显著差异(P>0.05),混交林土壤NO3--N含量显著高于毛竹纯林与日本柳杉纯林(P<0.05),说明在毛竹扩张过程中,扩张区域土壤NO3--N含量有增大趋势。

表1 不同林分类型土壤基本理化性质方差分析比较Table 1 Analysis of variance(ANOVA)of soil chemical properties relative to type of forest
2.2 毛竹扩张及模拟氮沉降对土壤N2O排放的影响
由图2可知,林分类型、模拟氮沉降以及二者的交互作用对土壤N2O的排放速率有显著的影响,林分类型、模拟氮沉降对N2O累积排放量有显著影响,但是二者的交互作用对其无显著影响。3种林分类型的CK的N2O的排放速率无显著差异(P>0.05)。但是从累积排放量来看,混交林与其他两种林分类型的N2O累积排放量差异极显著(P<0.01)。虽然3种林分类型CK的土壤N2O的排放速率差异未达到显著水平,但混交林排放速率略高于其他林分类型,所以经过时间的累积效应,导致排放速率无显著差异但累积排放量差异却显著。
如图2,模拟氮沉降对各林分类型的土壤N2O排放影响不一致。日本柳杉纯林CK与氮沉降处理之间差异极显著(P<0.01),氮沉降促进了日本柳杉纯林土壤N2O排放速率和累积排放量。毛竹纯林CK与氮沉降处理之间无显著差异(P>0.05),氮沉降未对毛竹纯林土壤N2O排放产生影响。混交林CK的土壤N2O排放速率与氮沉降处理间无显著差异(P>0.05),氮沉降未对混交林的N2O排放速率产生显著影响,但是从累积排放量来看,CK与氮沉降处理的累积排放量间差异显著(P<0.05),氮沉降增大了混交林土壤N2O累积排放量。

图2 不同林分类型及模拟氮沉降条件下土壤N2O排放速率及累计排放量比较Fig. 2 Comparison between the forest soils in soil N2O emission rates and cumulative emission as affected by nitrogen deposition
2.3 毛竹扩张及模拟氮沉降对土壤CO2排放的影响
从图3看出,在本研究中,林分类型是影响土壤CO2排放的重要因素,而氮沉降及其与林分类型的交互作用对土壤CO2排放无显著影响。无论是CO2排放速率还是累积排放量,日本柳杉纯林CK与其他两种林分类型CK之间差异均极显著(P<0.01),毛竹纯林CK与混交林CK之间无显著差异(P>0.05)。三种林分的土壤CO2排放速率呈现出毛竹纯林(896 g·m-2·h-1)、混交林(887 g·m-2·h-1)大于日本柳杉纯林(578 g·m-2·h-1)的趋势,累积排放量呈现毛竹纯林(4 276 g·m-2)、混交林(4 655 g·m-2)大于日本柳杉纯林(2 815 g·m-2)的趋势,说明毛竹扩张增加了扩张区域土壤CO2的累积排放量。图3可以看出,氮沉降并未对3种林分类型的土壤CO2排放产生显著影响。
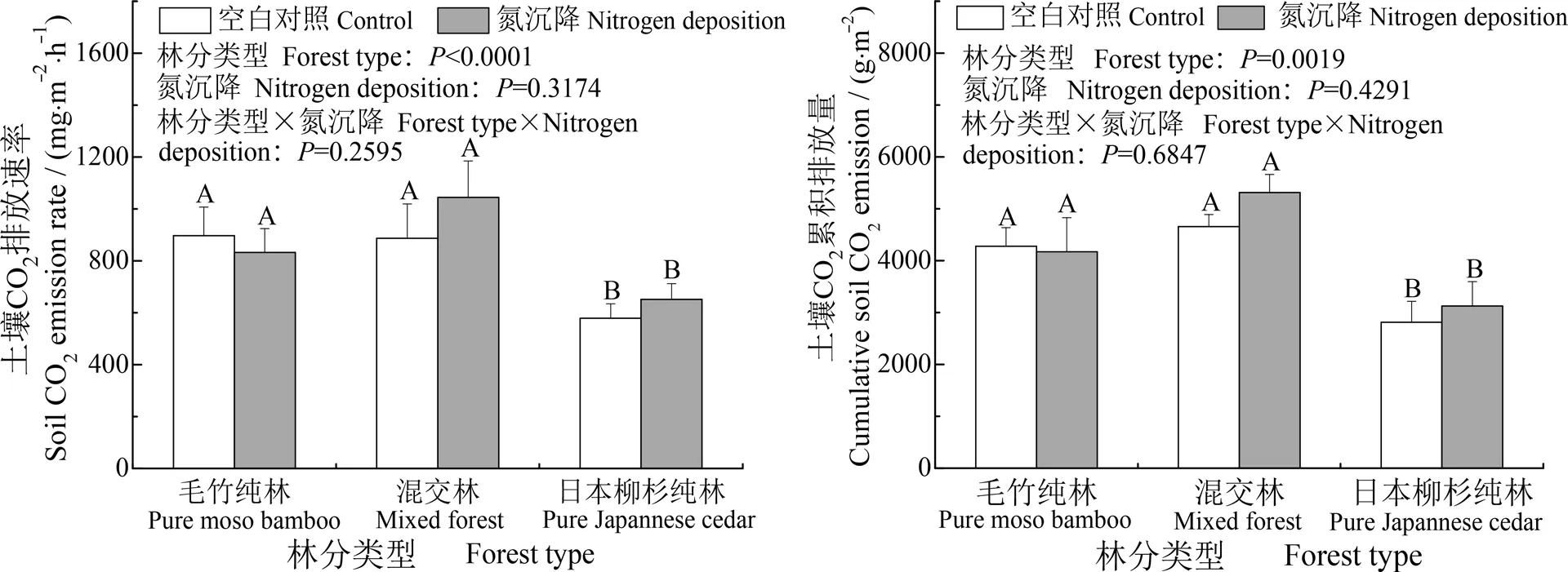
图3 不同林分类型及模拟氮沉降条件下土壤CO2排放速率及累计排放量比较Fig. 3 Comparison between the forest soils in soil CO2 emission rates and cumulative emission as affected by nitrogen deposition
3 讨 论
3.1 毛竹扩张对土壤理化性质的影响
植被类型转变会影响土壤基本理化性质和养分循环[22]。相对于日本柳杉纯林,混交林的土壤、含量增加,pH升高,土壤DOC含量降低,说明在毛竹扩张进入日本柳杉纯林的过程中,土壤的碳、氮循环过程发生了变化。与毛竹扩张进入的阔叶林相比,毛竹并未产生较多的凋落物输入量,较低的毛竹凋落物输入量导致其向土壤较低的氮输入量[10],从而导致毛竹林土壤较低的TN含量和N2O排放量(表1,图2)。但是,毛竹扩张形成的混交林的土壤TN含量和N2O排放量却均高于毛竹纯林和日本柳杉纯林(表1,图2),这可能是因为毛竹扩张进入日本柳杉林提高了土壤固氮菌群落多样性,但是随着年限延长,固氮菌群落结构差异、多样性及丰度降低减小,故在毛竹林扩张初期的混交林土壤TN含量高于生长多年的毛竹纯林[23]。同时,与日本柳杉纯林相比,混交林的土壤pH较大,说明毛竹扩张导致了土壤pH增加(表1),这可能会改变一些氨氧化细菌和与硝化作用相关的古生菌群落的改变[24],从而影响土壤氮转化过程。
3.2 毛竹扩张对土壤N2O和CO2排放的影响
虽然许多氮转化过程会导致土壤N2O排放,但是硝化和反硝化作用是形成N2O的主要过程,故凡是对这两个过程具有影响的因素如:土壤pH、碳氮底物可利用性、土壤温度、土壤水分、微生物群落结构和丰度等发生变化均会导致土壤中N2O排放通量的变化[25]。土壤中的和作为土壤硝化作用与反硝化作用的反应底物,与土壤N2O排放量密切相关。本研究中,混交林中的含量较高,含量较低,说明在毛竹扩张过程中,对土壤的氨化和硝化作用产生了影响,这可能也是毛竹林土壤N2O排放量较低的原因[16],这与Song[10]、Li[16]等的研究结果一致。毛竹纯林与日本柳杉纯林土壤的TN含量、含量、pH均差异显著,但含量无显著差异,而且土壤N2O排放速率和累积排放量无显著差异,说明反硝化作用可能是其产生N2O的主要过程[26],而混交林的含量高于其他两种林分类型,这可能也是混交林土壤N2O排放量高于其他两种林分类型的主要原因。此外,在毛竹扩张的过程中,土壤的pH升高,这可能缓解了pH对于土壤微生物活动的限制,改变了一些氨氧化细菌和与硝化作用相关的古生菌群落,促使土壤微生物的活性和多样性发生改变,影响土壤的硝化和反硝化细菌的活动,进而影响了土壤N2O的排放。
土壤呼吸主要由植物的根系呼吸、土壤微生物的异养呼吸以及土壤动物呼吸和部分化学氧化产生,其中植物根系呼吸和土壤微生物的异养呼吸是CO2产生的主要过程[27]。因为毛竹具有发达的地下茎(竹鞭),且繁殖能力强,当毛竹扩张进入日本柳杉纯林形成混交林,会显著增加林地土壤中的细根生物量[28],进而增大根系呼吸的CO2产生量。同时,毛竹根茎的快速繁殖生长通常伴随着较多的细根或根际沉积的碳输入,这可能会增加土壤的TOC和DOC含量,进而增加扩张区域土壤中的CO2排放[16]。通常情况下,日本柳杉纯林的土壤pH低于毛竹纯林,这可能限制了土壤微生物对土壤有机碳的分解。然而,随着毛竹不断扩张进入日本柳杉林,形成的混交林的土壤pH升高,缓解了pH对于土壤微生物活动的限制,导致更多的土壤有机碳被分解,从而促进混交林土壤CO2的排放通量,所以,这可能也是混交林DOC含量低于其他两种林分类型的原因。此外,凋落物的C/N是反映其分解速率快慢的重要因素,毛竹凋落物与日本柳杉凋落物的OC含量(443.4、550.4 g·kg-1)、TN含量(13.7、6.89 g·kg-1)及C/N(33.53、85.01)均差异显著。可以看出,与日本柳杉相比,毛竹凋落物的C/N更低,说明其更易分解。毛竹扩张进入日本柳杉纯林后,其凋落物分解较快,能较快地释放更多的养分到土壤中,缓解微生物活动的养分限制,促进微生物对凋落物的分解,增加土壤的碳输入,从而增加混交林土壤CO2的排放[29]。此外,研究发现,较低的凋落物C/N的森林土壤中微生物活性更高[29],输入的毛竹凋落物较低的C/N可能激发混交林土壤的微生物活性,从而影响土壤有机碳的分解,同时提高土壤微生物异养呼吸产生的CO2量,进而增大土壤的CO2排放量,这可能也是毛竹纯林和混交林土壤CO2排放量高于日本柳杉纯林的原因之一。
3.3 模拟氮沉降对土壤N2O和CO2排放的影响
模拟氮沉降作为一种外源氮输入方式,会改变土壤中氨氧化细菌(Ammonia-oxidizing bacteria,AOB)和氨氧化古菌(Ammoniaoxidizing archaea,AOA)的群落丰度和活性,而AOB或AOA会在氨单加氧酶和羟胺氧化还原酶的催化下,将NH3氧化成,该反应的中间产物NH2OH会分解释放N2O,是硝化作用的关键步骤,也是限速步骤[30]。但是目前关于氮沉降对土壤N2O排放的研究结果并不一致,一般分为促进、抑制及无影响三种结论[31]。与毛竹纯林和混交林相比,日本柳杉纯林的土壤N2O排放对于模拟氮沉降的响应最为剧烈,模拟氮沉降促进了日本柳杉纯林的土壤N2O的排放速率,增加了其累积排放量,这说明在日本柳杉纯林土壤中,氮源可能是其土壤排放N2O的限制因素。而且,日本柳杉纯林的土壤含量最低(表1),这是不利于土壤硝化作用的,但是在毛竹扩张进入后,提高了土壤、含量,潜在提高了土壤N2O产生的氮源水平,这与Li等[16]的研究结果是一致的,说明毛竹扩张改变了土壤N循环。但也有可能是因为施N后,日本柳杉纯林N吸收量低于毛竹纯林,更多的氮素以气体或淋溶形式流失而导致N2O排放量的增加。氮沉降对混交林土壤N2O排放速率无显著影响,却增加了其N2O累积排放量。这可能是因为随着毛竹的不断扩张进入日本柳杉林,混交林土壤的各项理化性质趋近于毛竹纯林,所以氮沉降未显著促进其排放速率,但是经过时间累积,其排放量显著增大,说明毛竹扩张可能进一步增加了氮沉降对日本柳杉纯林土壤N2O排放的促进作用。
氮沉降一般通过改变植物根系生物量、凋落物分解速率、土壤C/N、及土壤微生物数量和活性而影响土壤CO2的排放速率[32]。但是目前关于氮沉降对土壤CO2排放的研究结果并不一致,一般分为促进、抑制及无影响三种结论。在本研究中,模拟氮沉降并未对庐山3种不同林分类型土壤的CO2排放产生显著影响(图3),这与Micks[33]和莫江明[34]等的研究结果一致。主要原因可能如下:(1)土壤中含量较高(表1),对木质素分解的酶活性产生了抑制作用,降低了其分解速率,从而减少了木质素分解所产生的CO2量,但是可能促进了微生物的呼吸作用,当二者减少和增大的CO2量相当时对土壤CO2排放不产生影响[35]。(2)外源氮激发了土壤微生物活性,从而增加土壤微生物呼吸产生的CO2量,但是因为氮添加增加了土壤中的有效氮含量,导致根系吸收N素所需消耗的能量减小,当二者呼吸作用量相当,可能相互抵消而对土壤的CO2排放不产生影响[36]。
3.4 林分类型与模拟氮沉降的交互作用对土壤N2O和CO2排放的影响
在本研究中,氮沉降与林分类型的交互作用对土壤N2O累积排放量、CO2排放速率及CO2累积排放量无显著影响,但是对N2O的排放速率具有显著影响。氮沉降显著促进了日本柳杉纯林的N2O排放速率,但对毛竹纯林和混交林的N2O排放速率没有产生显著影响。在本试验中,氮沉降处理使用尿素(CO(NH2)2)溶液,因为尿素溶液易水解形成,而毛竹纯林与混交林土壤的显著高于日本柳杉纯林(表1),所以外界的输入增大了日本柳杉纯林的N反应底物,促进土壤硝化作用,进而导致其N2O排放速率的增大。此外,日本柳杉纯林土壤pH显著低于毛竹纯林和混交林(表1),这可能不利于土壤微生物的活动。然而,尿素水解产生的同时也会产生OH-,这对于日本柳杉纯林的酸性土壤有一定的中和作用,能在一定程度上增大土壤pH,从而增强反硝化酶活性、促进反硝化作用的进行,进而促进土壤N2O的产生[37]。
4 结 论
毛竹扩张过程中一定程度上增大了土壤N2O和CO2的排放量,但是完全扩张后N2O排放出现明显下降趋势,而CO2的排放未发生显著变化。氮沉降显著促进了柳杉纯林和混交林的N2O排放,但是对毛竹纯林N2O及3种林型土壤的CO2排放无显著影响。
