甜菜夜蛾几丁质脱乙酰酶SeCDA2a的外源表达及酶活力测定
2019-02-22陈祎,张伟,赵丹,郭巍*
陈 祎,张 伟,赵 丹,郭 巍*
(1.北京农学院植物科学技术学院,北京102206;2.河北农业大学植物保护学院,河北保定071001)
甜菜夜蛾SpodopteraexiguaHübner属鳞翅目Lepidoptera、夜蛾科Noctuidae,是一种全球性害虫,寄主繁多,中国20多个省、市、自治区均有甜菜夜蛾的为害记录[1]。由于甜菜夜蛾迁飞性惊人,世代重叠严重,发生规律复杂,寄主植物种植面积不断扩大,并且长期以来,单一使用化学药剂防治、盲目增加药量,使得该害虫抗药性不断增加,防治效果不理想[2]。如何安全、有效防治甜菜夜蛾是一个亟待解决的问题。几丁质是昆虫表皮的主要结构成分,参与昆虫生长发育的全过程[3]。昆虫发育需要经历周期性的蜕皮和形成新表皮的过程,伴随着几丁质代谢[4]。几丁质脱乙酰酶(chitin deacetylase, CDA)的主要功能是催化几丁质N-乙酰氨基-D-葡萄糖胺脱去乙酰基变成脱乙酰几丁质[5],是几丁质降解过程中极为重要的一类酶。
自2005年Guo等首次从鳞翅目夜蛾科昆虫粉蚊夜蛾(Trichoplusiani)中分离出CDA蛋白——TnPM-P42,并验证其具有几丁质结合活性后,研究者相继从其他昆虫中克隆得到编码CDA的基因并研究其重组CDA的催化活性[6]。2008年,Toprak等从蓓带夜蛾(Mamestraconfigurata)中肠分离得到McCDA1,其原核表达的McCDA1蛋白具有几丁质脱乙酰活性[7]。2012年,Zhong等在毕赤酵母中表达家蚕(Bombyxmori)BmCDA7,测得其几丁质脱乙酰酶活力为1.85 U/mL[8]。2015年,孙晓彤等在毕赤酵母GS115表达甜菜夜蛾SeCDA1,其几丁质脱乙酰酶活力为1.35 U/mL[9]。闫晓平等测得由甲醇诱导在毕赤酵母中表达的美国白蛾(Hyphantriacunea)HcCDA1重组蛋白的酶活力为1.685 U/mL[10]。2017年,赵盼等克隆得到飞蝗(Locustamigratoria)3个几丁质脱乙酰酶基因,测得由Ni-NTA亲和层析柱纯化后于Sf9细胞中表达的重组蛋白LmCDA1、LmCDA2a和LmCDA2b的酶活力分别为0.268、0.354和0.228 U/μL[11]。近年来研究表明,几丁质脱乙酰酶可以作为杀虫靶标,通过干扰几丁质代谢过程来实现对害虫的防治,具有广泛的应用前景。
本研究对编码甜菜夜蛾几丁质脱乙酰酶的基因Secda2a进行外源表达,并测定重组蛋白SeCDA2a的酶活力,从分子水平上研究甜菜夜蛾几丁质脱乙酰酶,为明确该酶在甜菜夜蛾幼虫体内的作用机制奠定理论基础,为研发新的生防措施提供理论依据。
1 材料与方法
1.1 材 料
1.1.1 供试昆虫和昆虫细胞培养 甜菜夜蛾为本实验室人工饲养种群,幼虫以人工饲料饲养,成虫喂食10%白糖水。人工气候箱设置成以下参数:温度(26.5±1)℃,湿度为50%±5%,光照L∶D=14 h∶10 h。草地贪夜蛾细胞系(Sf9),于27℃人工气候箱中培养,所用的Grace培养基加入10%的胎牛血清。
1.1.2 主要试剂及菌株 pMD19-T-Secda2a质粒(含SacI &NotI酶切位点及目的基因)、pET-28a质粒及pFastBac HT A质粒由本实验室保存。限制性内切酶、蛋白Marker购自Thermo公司;T4连接酶购自Promega公司;LA Taq酶、DNA Marker、DH5α感受态细胞购自Takara公司;质粒提取试剂盒及DNA凝胶回收试剂盒购自Axygen公司;BL21(DE3)感受态细胞、His抗体购自康为试剂公司;DH10Bac感受态细胞购自北京华越洋生物科技有限公司;Bacmid纯化试剂盒及Cellfectin Ⅱ reagent购自Invitrogen公司;胎牛血清购自四季青公司。
1.2 方 法
1.2.1 原核表达载体的构建与鉴定 用SacI &NotI酶切pMD19-T-Secda2a质粒和pET-28a质粒,使用凝胶回收试剂盒纯化酶切产物。使用T4连接酶将目的基因Secda2a和载体于4℃连接过夜。次日,产物转化E.coliBL21。挑取阳性重组子,提取质粒后进行PCR和酶切验证,验证正确后送至北京六合华大基因科技有限公司测序。
1.2.2 原核表达 将验证正确的阳性重组子接种于含有卡纳霉素的LB液体培养基中,37℃,220 r/min振荡培养过夜,次日,向含有卡纳霉素的新鲜LB液体培养基中接入1%体积的过夜培养的菌液,培养3~3.5 h,至OD600为0.5时,添加诱导物IPTG使其终浓度为0.50 mmol/L,于37℃对其进行诱导表达,并于诱导2、4、6、8 h分别取样1.5 mL,用于分析SeCDA2a重组蛋白的表达。
1.2.3 引物设计 使用DNAMAN 7.0软件分析Secda2a全长序列酶切位点,结合pFastBac HT A载体上的多克隆位点,设计基因特异引物,两端分别添加NcoⅠ &NotⅠ的酶切位点及保护碱基(F: 5′-CATGCCATGGATGGCTCGACACGCGTGTC TGTGTCTT-3′;R:5′-ATTTGCGGCCGCTCA CTTCTTAACATTGAAGCCTTCTCCC-3′),引物序列中的酶切位点用下划线标出。
1.2.4 重组Bacmid的构建及鉴定 以pMD19-T-Secda2a质粒为模板,使用1.2.3中引物PCR扩增获得Secda2a全长基因,经凝胶回收试剂盒纯化回收,PCR产物和pFastBac HT A载体分别用NcoⅠ &NotⅠ酶切,酶切产物纯化后使用T4连接酶以5∶1的比例连接过夜,产物转化E.coliDH5α,挑取阳性重组子进行PCR和酶切验证。鉴定正确后将重组转座载体pFastBac HT A-Secda2a转化DH10BacTM感受态细胞,获得重组杆状病毒Bacmid-Secda2a。利用通用引物PUC/M13 F&R 进行PCR鉴定。
1.2.5 重组蛋白SeCDA2a在昆虫细胞Sf9中的表达 选取鉴定正确的重组Bacmid,纯化后利用Cellfectin Ⅱ reagent转染昆虫细胞Sf9,获得重组病毒P1,继续侵染Sf9细胞分别得到P2和P3病毒,收集P3病毒上清得到重组蛋白,具体步骤参照Invitrogen Bac-to-Bac昆虫杆状病毒表达系统说明书进行。以转染空载体的Sf9细胞培养液上清为对照,进行SDS-PAGE电泳,Western blot分析重组蛋白SeCDA2a的表达。
1.2.6 重组蛋白SeCDA2a酶活力测定 对硝基乙酰苯胺是一种无色晶体,易水解生成黄色针状结晶体对硝基苯胺,这种黄色晶体的水溶液在400 nm处有特异性吸光值。参考刘丽萍等的方法绘制对硝基苯胺标准曲线,参考Zhong等的方法用硫酸铵对真核重组蛋白SeCDA2a进行纯化作为酶液参与酶促反应[8,12]。
2 结果与分析
2.1 原核表达载体的构建与鉴定
目的基因Secda2a与pET-28a载体连接后转化大肠杆菌,挑取阳性重组子,提取质粒经PCR鉴定后,获得约1.6 kb的目的片段(图1)。重组质粒pET-28a-Secda2a经SacI &NotI酶切得到1.6 kb的Secda2a基因片段和5.4 kb的载体片段,与预期大小一致(图2)。表明原核表达载体pET-28a-Secda2a构建成功。

图1 重组质粒PCR鉴定Fig.1 PCR identification of recombinant plasmid

图2 重组质粒pET-28a-Secda2a酶切鉴定Fig.2 Restriction analysis of of pET-28a-Secda2a
2.2 SeCDA2a重组蛋白的原核表达
用终浓度为0.50 mmol/L IPTG在37℃对重组蛋白进行诱导,分别在诱导2、4、6、8 h取样,收集菌体沉淀进行检测。结果显示,E.coliBL21成功表达出约61 kDa的目的蛋白,且于4 h之后表达量达到最大(图3)。

注:0 h是pET-28a-Secda2a未经IPTG诱导;2、4、6、8 h是pET-28a-Secda2a经IPTG诱导2、4、6、8 h。Note: 0 h represents pET-28a-Secda2a not induced by IPTG; 2, 4, 6, 8 h represent pET-28a-Secda2a induced by IPTG for 2, 4, 6, 8 h.图3 SDS-PAGE与Western blot检测IPTG诱导表达的SeCDA2a蛋白Fig.3 SDS-PAGE and Western blot analysis of SeCDA2a expression induced by IPTG
2.3 重组Bacmid的构建及鉴定
以重组质粒pMD19-T-Secda2a为模板,利用特异性引物PCR扩增得到约1.6 kb的片段(图4)。重组质粒pFastBac HT A-Secda2a经NcoⅠ &NotⅠ酶切验证正确,表明重组转座载体成功构建(图5)。利用通用引物对重组Bacmid进行PCR验证,获得约3.9 kb的目的条带,表明重组Bacmid-Secda2a构建成功(图6)。
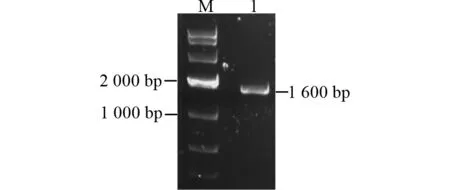
图4 甜菜夜蛾Secda2aPCR扩增产物Fig.4 PCR amplication product of Secda2a
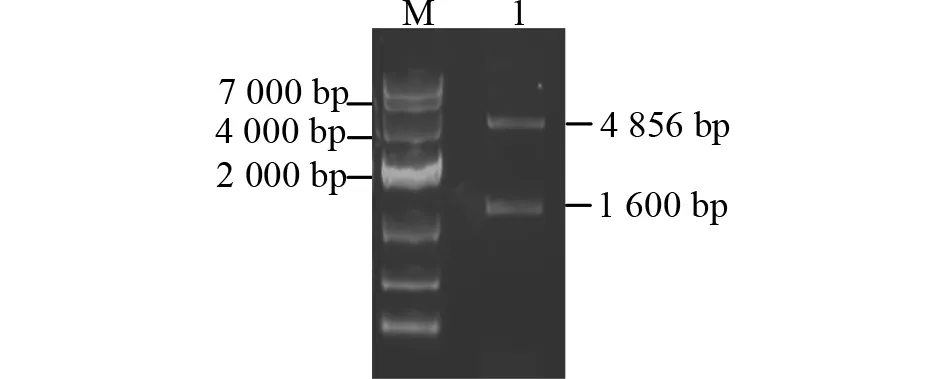
图5 重组质粒pFastBac HT A-Secda2a酶切鉴定Fig.5 Restriction analysis of pFastBac HT A-Secda2a
2.4 重组蛋白SeCDA2a在昆虫细胞Sf9中的表达
Bacmid转染生长状态良好的Sf9细胞后,获得原代病毒,传代培养至第3代,Western blot检测SeCDA2a在昆虫细胞中的表达,得到约61 kDa的重组蛋白(图7)。

图6 重组Bacmid验证Fig.6 Dection of recombinant Bacmid

注:CK是未感染重组病毒的Sf9细胞培养液上清;1是感染重组病毒的Sf9细胞培养液上清。Note: 1 represents supernatant of Sf9 cell culture infected by Bacmid; CK represents supernatant of Sf9 cell culture not infected by Bacmid.图7 重组蛋白SeCDA2a在昆虫细胞Sf9中的表达Fig.7 Recombinant protein of SeCDA2a expression in Sf9 cells
2.5 重组蛋白SeCDA2a酶活力测定
本研究中对硝基苯胺的标准曲线的线性回归方程是y=0.1362x+0.0252,R2为0.9988,符合朗伯-比尔定律(图8)。呈浓度梯度的对硝基苯胺溶液在400 nm处的吸光值具有良好的线性关系,可以通过分光光度法定量测定对硝基苯胺。本研究测得Sf9细胞表达的重组蛋白SeCDA2a的酶活力是1.57 U/mL。

图8 对硝基苯胺标准曲线Fig.8 Standard curve of p-nitroaniline
3 讨 论
CDA是一种几丁质修饰酶,在昆虫体内的主要功能是脱去表皮几丁质中的乙酰基,生成脱乙酰几丁质,参与昆虫体内几丁质的代谢[13,14]。该酶可用于扰乱昆虫正常生长机制,从而达到对害虫生物防治的目的。
到目前为止,大量昆虫几丁质脱乙酰酶基因的功能通过转录水平分析和RNA干扰技术得以研究。根据序列相似度及所含保守区域多样性的不同,几丁质脱乙酰酶被分为I-V,共5类。其中,第I类和第Ⅱ类均包含三个保守区域,几丁质结合区、多聚糖乙酰基转移酶催化区和低密度脂蛋白结合区,第V类仅包含一个多聚糖乙酰基转移酶催化区[15]。近年来,对昆虫CDA蛋白的功能研究成果主要集中在两方面:一方面,第I类CDA蛋白参与调控昆虫的蜕皮过程[15-19],另一方面,第V类CDA蛋白参与中肠免疫[20]。笔者在NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)上分析SeCDA2a蛋白保守结构域,发现SeCDA2a包含三个功能域,属于第I类CDA蛋白,推测其在甜菜夜蛾的蜕皮过程发挥作用。
体外表达有催化活性的蛋白对于更好研究SeCDA2a的功能意义重大。本课题组孙晓彤在E.coli中表达61.5 kDa的重组蛋白SeCDA1(属于第I类CDA蛋白),在毕赤酵母细胞和昆虫细胞High Five中均表达80 kDa的重组蛋白SeCDA1,并测出两种真核细胞表达的重组蛋白SeCDA1的酶活力分别为1.88 U/mL和1.35 U/mL[21]。本研究首次在大肠杆菌BL21和昆虫Sf9细胞中成功表达重组蛋白SeCDA2a,检测真核重组表达蛋白的几丁质脱乙酰酶活性;测得酶活力与孙晓彤等[21]、闫晓平等[22]的结果接近。为了进一步明确SeCDA2a蛋白的功能,仍需从转录水平和蛋白水平上对Secda2a基因进行组织定位分析和不同发育时期表达量分析;利用RNA干扰技术将Secda2a基因沉默,通过观察甜菜夜蛾的蜕皮情况,利用实时荧光定量PCR技术检测基因的沉默率。本研究实现甜菜夜蛾几丁质脱乙酰酶基因Secda2a的外源表达,测定重组蛋白SeCDA2a的酶活力。
