水分和养分添加对扭黄茅刈割后补偿作用的影响
2019-02-19闫帮国何光熊张梦寅和润莲史亮涛
孙 毅,闫帮国,何光熊,张梦寅,和润莲,史亮涛
(1. 云南省农业科学院热区生态农业研究所,云南 元谋 651300;2. 元谋干热河谷植物园,云南 元谋 651300)
萨瓦纳草地(savannah grassland)在云南元谋干热河谷(dry-hot valley)热坝已有较长的利用历史,但现在仅在坝周低山区成片分布(坝区已被灌溉条件良好的农田生态系统取代)。由于滥牧、缺乏管理、开发利用不充分等原因,造成水土流失、生物多样性下降等问题。刈割是常见的草地管理和利用方式之一。刈割或动物取食后牧草的补偿性生长模式包括超补偿生长(over-compensatory growth,刈割或取食后牧草的总产量增加)、等补偿生长(equalcompensatory growth,刈割或动物取食后牧草的总产量不变)和欠补偿生长(under-compensatory growth,刈割或动物 取食后牧草的总产量减小)[1-2]。刈割的观测包括频次、时期、间隔和方式等几个维度,适宜的刈割一方面能最大限度地利用植物的补偿性生长和均衡性生长特性,另一方面可改变植物形态和生理指标及其体内营养物质的沉积和分配方向,从而提高牧草的生产性能、品质,并延长牧草的利用时期[3-4]。刈割强度随刈割方式的不同影响着植物的补偿生长。例如,有对禾本科多年生耐牧牧草的研究[5]显示,羊草(Leymus chinensis)当年总生物量在中度刈割后显著增加40%,而在重度刈割后下降5%,其原因是重度刈割常导致牧草丢失过多的光合器官和营养物质,以致牧草的光合能力下降,影响牧草以后各年的总产量;而低强度刈割则会将营养价值较高的叶片、嫩茎等器官遗留较长时间,导致牧草产量低、营养利用不充分[3]。
除刈割强度外,刈割后植物的补偿性生长还与其生境中资源的可获取性密切相关,而水分或养分添加是草地管理过程中常采取的资源补充方式。牧草的高产优质只有在生境中水分和养分供应充足的情况下才能实现,适当地添加水分和养分可以提高羊草草地的产草量[6]。适宜地进行水分、养分添加后,禾草生物量的增加主要是由于其营养枝上叶片面积增大,禾草的光合作用同化效率提升,从而得以生产和积累更多营养物质[6]。此外,刈割处理造成了植物地上组织的损失及其引起的根冠比失调,这可以缓解邻近植株对生境中光照、水分、养分等资源的激烈竞争,但对于相邻的禾本科植株从幼苗期开始形成的不对称竞争格局没有影响[7]。
扭黄茅(Heteropogon contortus)是金沙江流域干热河谷坝区,特别是其坝周低山丘陵区萨瓦纳草地群落中的优势种群[8-9],嫩时牲畜喜食,但至花果期(4-12月)小穗的芒及基盘常穿透牲畜皮肤,为害牲畜。目前已有的扭黄茅相关研究主要包括其土壤种子库[10-11]、种子萌发[12-13]、耐旱机制[14-15]、群落生物多样性[16-17](包括昆虫[18-19]、根际AM真菌[20-21])、种间关系[22-23]、生态位[24-25]和水土保持[26-27]等方面,而对以扭黄茅为材料进行刈割或采食处理后的补偿生长及施肥处理后幼苗生长的研究比较缺乏。因此,本研究以盆栽的扭黄茅幼苗为材料,经过不同的水分和养分添加处理后,研究了刈割对扭黄茅补偿生长的影响,为扭黄茅的苗期管理、草地管理及利用等提供依据。
1 材料与方法
1.1 幼苗的培育
试验所采用的植物为云南省元谋县乡土草本扭黄茅,植物种子于2011年10-12月采自金沙江中游 元谋段干热河谷区 (25.4°-26.1° N,101.6°-102.1° E,海拔 1 100 m)。该区地处滇中高原北部,南亚热带干热季风气候,年平均气温21.9 ℃,年均降水量613.8 mm,雨季(5-10月)降水量约占年降水量的91.2%[28],年平均霜日2 d,年平均日照时数2 664.5 d,土壤类型主要有燥红土、变性土和紫色土[29]。该区既是云南省最大的冬早蔬菜主产区之一,又是长江中下游泥沙的主要产区之一。区域内主要的自然植被类型近似于热带稀树草原,以禾本科为群落优势种和建群种,植被退化和水土流失严重。
2012年6月7日,在云南省农业科学院热区生态农业研究所的大棚内开始育苗。将24只花盆(直径33 cm,高度25 cm,底部有孔并附有托盘)洗净晾干后,每盆装入已混合均匀的燥红土10 kg,在各盆内分别埋入种子 5 g(埋深约 0.2 mm)。播种 5 d后种子开始发芽。播种12 d后间苗,每盆保留大小基本一致的壮苗4株进行后续试验。
1.2 幼苗的水和养分添加以及刈割处理
于2012年6月28日开始进行水和养分添加处理。先将24盆分为2组,每组12盆用于水分处理,再把各组分为2个亚组,每亚组6盆进行养分添加处理。2个组的水分处理分为H组(高频次降水,模拟中度干旱,每30 d浇12次水,每次300 mL)和L组(低频次降水,模拟重度干旱,每30 d浇8次水,每次300 mL)[29-31]。2个亚组的养分处理分为F 组(每 15 d 添加1 次,每 次添加 1/6 Hoagland溶液300 mL)和I组(不添加)。另外,及时拔除杂草,并将所有花盆每隔1个月进行一次透彻浇水,以促使多余的养分流失,避免毒害。
于2012年8月30日进行刈割处理。先将各亚组的6盆分为2个小组,每小组3盆,以进行刈割处理,即为3次重复。刈割处理分为处理C(留茬1 cm)和处理O(不进行刈割)。将刈割时收获后烘干的生物量记为刈割干物质重量。
1.3 植株的收获和生长参数测定
2012年9月19日开始收获植株并及时测定生长参数。收获时,先收集凋落物,再将植物地上部分分割并挖出根系,带回实验室洗净后,将植物根、茎(含叶鞘[32])、叶、花果及凋落物分开后烘干(75 ℃,48 h),用于测定生物量,即凋落物干物质重量(DF)、根干物质重量(DR)、茎干物质重量(DS)、叶干物质重量(DL)、花果干物质重量(DI)均采用烘干法测定,并根据上述数据计算单株干物质重量 (DT),公式为 DT= DR + DS + DL + DI;植株及其各器官、凋落物的生物量补偿指数(CIB)按王海洋等[33]的定义计算。
1.4 数据的处理
数据的录入和整理均在WPS表格内完成,使用IBM SPSS Statistics 21 对数据进行统计和分析,使用SigmaPlot 12.5完成试验结果的绘图。所有进行方差分析的试验数据先经过对数转换,以使其更符合正态分布,用于多重比较的数据进行方差齐性检验(Homogeneity of Variance Test)。采用二因素方差分析(Two-Way ANOVA)分析水分、养分添加及二者的交互作用对生物量补偿指数的影响,采用单因素方差分析(One-way ANOVA)的R-E-G-W-Q法对各因素不同水平间各参数影响的差异进行多重比较。使用配对样本T 检验法分析各处理不同水平对参数影响的差异性。

图1 不同水分、养分添加条件下刈割扭黄茅植株时收获的冠部干物质重量Figure 1 Dry matter weight of harvested crown when H. contortus was mowed after different water and nutrient treatments不同小写字母表示同一水分处理下不同养分处理间差异显著(P<0.05),不同大写字母表示同一养分处理下不同水分处理间差异显著 (P<0.05)。Different lowercase letters indicate significant difference between the different nutrient treatments with the same water treatment at the 0.05 level, different capital letters indicate significant difference between the different water treatments with the same nutrient treatment at the 0.05 level.
2 结果与分析
2.1 水分、养分和刈割对扭黄茅植株干物质积累的影响
截至进行刈割处理时,无论降水频次如何,养分添加均能增加扭黄茅冠部的干物质重量,在高频次降水时具有显著影响(P<0.05),此时刈割冠部可收获的干物质重量高达0.85 g(图1)。在添加养分的情况下,冠部干物质重量随着降水频次增加而增加;而在未添加养分的情况下,冠部干物质重量随着降水频次增加略有降低。
在低频次降水、未进行刈割,以及高频次降水、进行刈割时,添加养分后凋落物干物质重量显著提高(P<0.05)(图2);在进行低频次降水并添加养分时,刈割后的凋落物干物质重量显著降低(P<0.05)。在各水分和养分处理条件下,刈割均降低了凋落物量。在低频次水分处理下,刈割后草地上凋落物干物质重量降低至0。提高降水频次后,植株凋落物干物质重量增加,此时添加养分可导致凋落物干物质重量积累增多,在未进行刈割时最多,达 0.12 g。
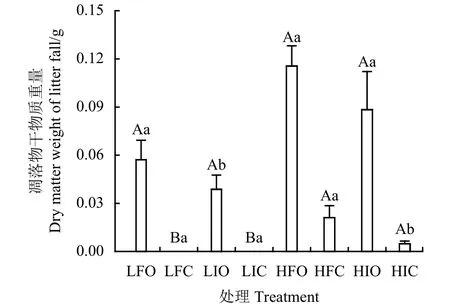
图2 不同水分、养分、刈割处理扭黄茅植株的凋落物干物质重量Figure 2 Dry matter weight of litterfall from H. contortus after different water, nutrient,and mowing treatmentsL和H分别表示低、高频次降水,F和I分别表示添加和未添加养分,O和C分别表示未刈割和刈割,下同。不同小写字母表示同一刈割处理下不同养分处理间差异显著(P<0.05),不同大写字母表示同一养分处理下不同刈割处理间差异显著(P<0.05)。L, Low-frequency precipitation; H, High-frequency precipitation; F,Nutrients added; I, Nutrients not added; O, Unmowed; C, Mowed;similarly for the following figures. Different lowercase letters indicate significant difference between different nutrient treatments under the same mowing treatment at the 0.05 level. Different capital letters indicate significant difference between different mowing treatments under the same nutrient treatment at the 0.05 level.
降水频次高低与否,扭黄茅单株干物质重量均表现为添加养分植株 > 未添加养分植株,未被刈割植株植株 > 被刈割植株;与其他处理相反,未添加养分而被刈割的扭黄茅单株干物质重量随着降水频次的增加而降低。高频次降水、添加养分条件下未被刈割的扭黄茅单株干物质重量最高,达3.88 g。与其他各处理相比,高频次降水、添加养分条件下未被刈割的扭黄茅根、茎、叶各器官干物质重量亦最高,分别为1.13、1.69、0.87 g,其花果干物质重量较高,但与其他各处理无较大差异。在各降水频次下,未添加养分而被刈割的扭黄茅单株根、茎、叶、花果各器官干物质重量均为最低,但各器官干物质重量在高、低频次降水处理间无较大差异(图3)。

图3 扭黄茅植株及其根、茎、叶、花果生物量对水分、养分及刈割处理的响应Figure 3 Response of biomass, roots, stems, leaves,inflorescences, and fruits of Heteropogon contortus to the water, nutrient, and mowing treatment
2.2 水分、养分和刈割对扭黄茅植株生物量分配的影响
未添加养分而被刈割的扭黄茅植株根冠比均显著高于其他处理 (P<0.05),分别为 1.08、1.06,但二者之间无显著性差异(P > 0.05)。无论降水频次高低及养分添加如何,刈割后植株的根冠比均升高 (图 4)。
低频次降水、添加养分、未被刈割处理,低频次降水、未添加养分、未被刈割处理,低频次降水、添加养分、被刈割处理,以及高频次降水、添加养分、未被刈割处理4个处理下植株的生殖分配间差异不显著(P > 0.05),但均显著小于高频次降水、未添加养分、未被刈割处理植株(P <0.05),均显著大于高频次降水、未添加养分、被刈割处理植株以及低频次降水、未添加养分、被刈割处理植株(P<0.05)。高频次降水条件下,添加养分、被刈割植株生殖分配率(5.03%)小于未添加养分亦未被刈割植株(6.91%),但二者间差异不显著(P > 0.05)。未添加养分而被刈割处理植株生殖分配率在降水频次高低间差异显著(P<0.05),分别为1.2%、0。养分添加与否,刈割后植株的生殖分配率在低频次降水均降低;而在高频次降水条件下,经刈割处理后,添加养分植株的生殖分配率增加(P > 0.05),未添加养分植株的生殖分配率显著降低 (P<0.05)(图 4)。
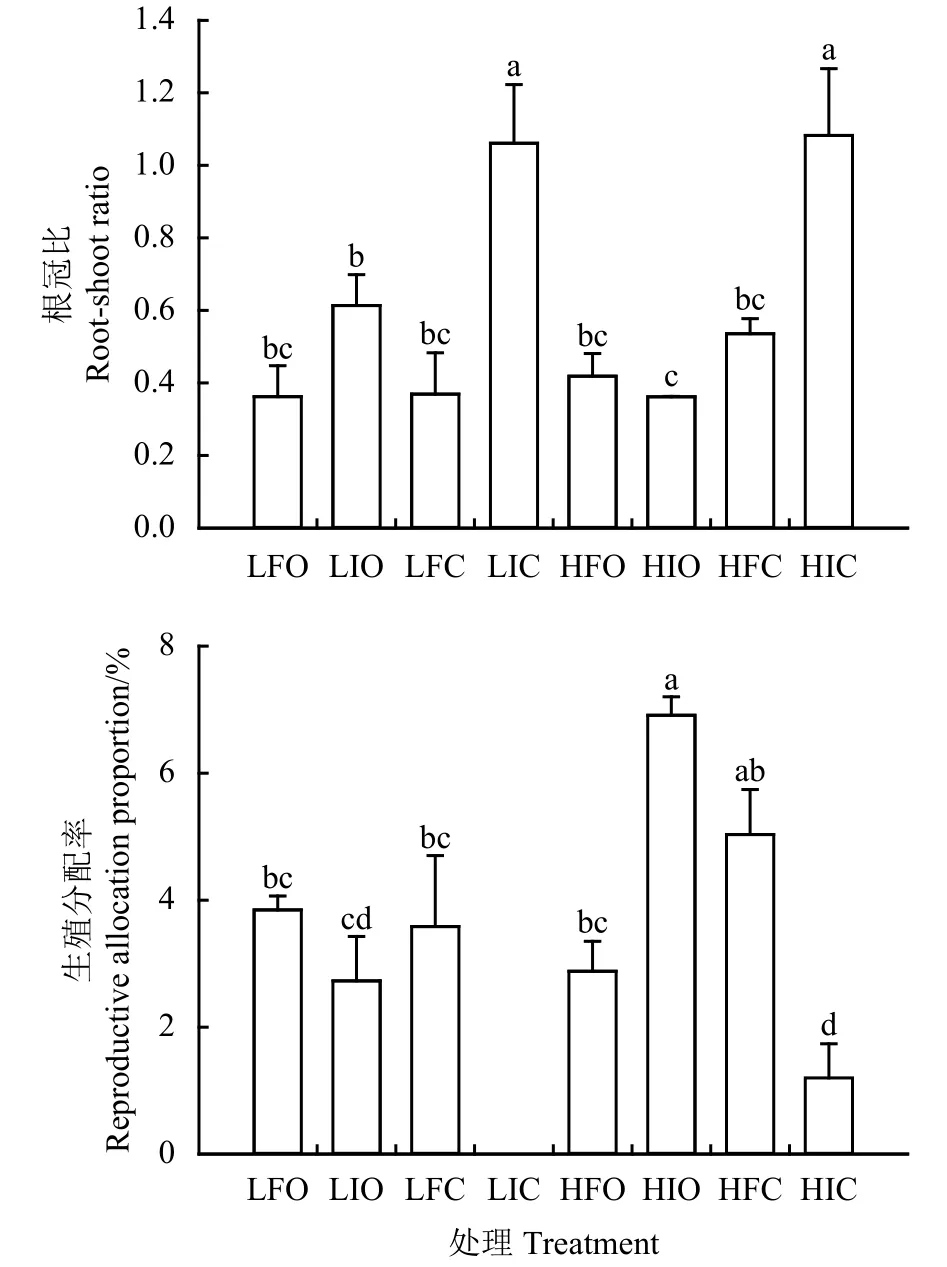

表1 水分、养分对扭黄茅植株凋落物、根、茎、叶、花果生物量及总生物量补偿指数影响的方差分析Table 1 Two-way ANOVA of water, nutrients, and their interactions on the compensation index of the biomass of litter, roots, stems, leaves, inflorescences and fruits, and total plant
2.3 不同水分、养分处理扭黄茅植株刈割后总生物量和各器官生物量的补偿生长
本研究中水分和养分处理的主效应对刈割后扭黄茅及其根、茎、叶、花果等各器官生物量的补偿生长均具有显著影响(P<0.05),而二者的交互效应仅对根、花果的生物量补偿生长有显著影响(P<0.05),对单株扭黄茅的总生物量补偿指数影响不显著(P > 0.05)。水分和养分处理的主效应及交互效应均对刈割与否的凋落物积累量变化有极显著影响 (P<0.01)(表 1)。
养分添加与否,刈割后,高频次降水时对根生物量补偿生长影响差异不显著(P > 0.05),低频次降水时对总生物量补偿生长的影响差异亦不显著(P > 0.05),其他处理下,植株及其各器官其生物量补偿指数均差异显著 (P<0.05)(表 2)。
同一养分处理下,不同水分处理间扭黄茅植株及其根、茎、叶、花果各器官的生物量补偿指数均无显著性差异(P > 0.05)。养分添加与否,在低频次降水条件下植株凋落物生物量补偿指数为0,在高频次降水条件下添加养分后显著提高(P<0.05),达0.19。凋落物生物量补偿指数在同一养分处理下不同水分处理间均差异不显著(P > 0.05)。扭黄茅植株总生物量及其根、茎、叶、花果各器官生物量的补偿指数均在低频次降水且进行养分添加条件下最高,分别为0.64、0.98、0.55、0.68、0.41,均小于1,为欠补偿生长。
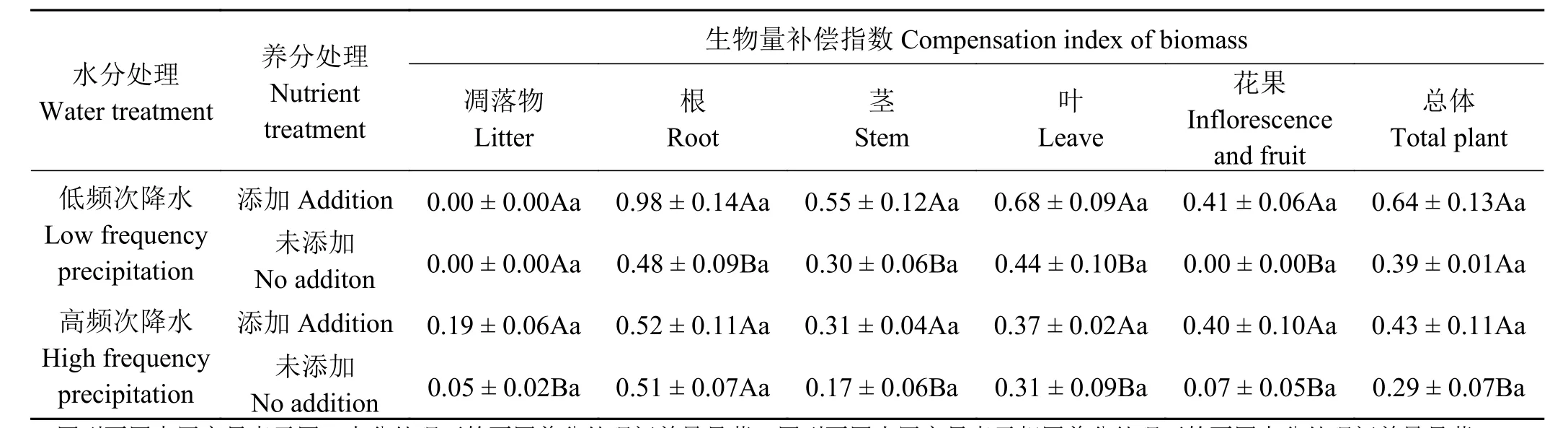
表2 不同水分、养分添加下扭黄茅刈割后凋落物、各器官生物量及总生物量的补偿指数Table 2 Compensation index of the biomass of litter, roots, stems, leaves, inflorescences and fruits,and total plant for different water and nutrient treatments
3 讨论与结论
3.1 水肥添加和刈割对扭黄茅植株凋落物形成的影响
本研究发现,刈割极不利于扭黄茅植株凋落物的形成,这与王仁忠[34]、吴东辉等[35]的研究结果,以及本研究在野外实地踏查过程中观测到被刈割或者放牧过的扭黄茅优势草地凋落物累积状况一致。这是由于刈割处理在直接去除植株成熟和衰老器官的同时,使植株丧失了大部分光合器官导致光合净同化减少,而且还改变了植物生理指标及其体内营养物质的沉积和分配方向用于补偿生长,从而抑制了凋落物的形成[36-37]。研究中水分或养分添加量增大后,植株凋落物积累量的增加证实了这一点。
3.2 水肥添加和刈割植株的补偿生长和生物量积累
本研究结果表明,生境中降水频次升高有利于扭黄茅植株生物量的积累。而在自然条件下,元谋干热河谷5-10月即为雨季,期间降水量和降水频次大幅增加,可达年降水量的91.2%[28],因此,进入雨季后,雨热同期的气候条件是扭黄茅积累生物量的有利条件。研究结果还显示,扭黄茅积累生物量时对雨热同期的气候条件的利用还受到生境中养分含量的限制,这与植株生长发育期间扭黄茅生境中资源的可利用性密切相关[6]。
对植株进行刈割处理后,无论水分、养分添加处理为何,扭黄茅植株的生物量均呈下降趋势,即刈割处理不利于植株生物量的积累。其原因应是由于重度刈割后植株同化器官大量减少,而刈割后补偿生长建成的新生器官同化产物不足造成的[6]。本研究中,在水分和养分状况一致的背景下,刈割后扭黄茅植株及其根、茎、叶、花果各器官的生物量补偿指数均小于1,均为欠补偿生长模式,这是由于刈割处理强度过大造成的。有研究表明,在干热河谷内生长的成熟扭黄茅植株营养物质常被大量储存于植株茎秆以增加光竞争能力[29],本研究对茎秆的刈割可能导致了植株在生长前期积累的营养物质大量丢失。
3.3 水肥添加和刈割植株的生物量分配
刈割会改变植物体内营养物质的沉积和分配方向,从而影响牧草的生产性能、品质和利用时期[3-4]。在本研究中,未添加养分、未刈割处理下的植株随着降水频次增加,植株根冠比呈降低趋势。这是由于水分增加后,未添加养分亦未刈割处理植株其生长受到地下生态因子(土壤水分和有效养分等)的限制相对缓解,而相对的受到地上生态因子(光照、CO2、O2等)限制后,将更多营养物质用于冠部生长并减少了对地下根系的投入[38]。在未添加养分条件下,对植株进行刈割后,临近植株对土壤养分竞争的相对减弱格局[7]与添加养分(无论是否刈割)后的临近植株竞争情况一致,因此在添加养分(无论是否刈割)处理下,以及刈割并且未添加养分处理下,植株根冠比的增加应当是对土壤水分相对亏缺现状的适应性变化。
在刈割情况下,养分添加(无论水分添加频率高低)降低了根冠比。根冠比降低后意味着植物可以将更多的生物量分配至地上,从而增加了植物光合能力和生长速度[39],提高植物补偿生长指数。从研究结果可以看出,养分添加确实增加了植物的补偿生长指数。
但是,在非刈割情况下,这种差异不显著。这表明养分与刈割的交互作用影响了根冠比。以往的研究认为养分增加后植物根系的生物量分配会下降,从而降低了根冠比[40]。但与此同时,水分也可以影响植物对根系的生物量分配[41]。水分是干热河谷植物生长的重要限制因素[42],在没有刈割的情况下植物具有较高的地上生物量,对水分需求较大,也就需要较高的根系生物量以吸收充足的水分。这可能是在未刈割处理下植物根系分配不受养分添加影响的原因。
植株生殖分配率在低频次降水、未添加养分、刈割处理下为0,并随着水分添加量、养分添加量、以及植株完整性的增加而增加,但均维持在较低水平。这说明扭黄茅植株的生殖分配依赖于一定的水分、养分条件和植株完整性。已有的研究发现干旱[43]、养分限制[44]和刈割[45]都会抑制植物的生殖生长和生殖分配。本研究结果表明3种因素共同作用时,植物生殖生长将受到严重抑制。
综上所述,刈割不利于扭黄茅草地凋落物的形成和生物量的积累,雨季可能是扭黄茅植株生物量和凋落物产生的高峰期,雨季适量施肥有助于缓解刈割对扭黄茅植株生物量积累的不利作用,但不改变刈割后扭黄茅欠补偿生长的格局,这可能是由于扭黄茅的遗传和生长特征导致植株将大部分营养物质积累到茎秆,而茎秆刈割强度较大导致植株营养物质过度流失造成的。在水分和养分添加及刈割各处理下,扭黄茅可以通过对植株各功能器官(根、茎、叶、花果)之间生长和生物量分配的权衡,因地制宜地实现植株对环境中资源的摄取,从而顺利地完成生活史过程。
