福建省2015年本地登革热病例的病原学特征
2019-02-19,,,,,
,,,,,
登革热是由登革病毒感染引起的、经伊蚊传播的一种急性传染病,临床表现为突起发热,伴随头痛,全身肌肉、骨骼和关节痛,皮疹,白细胞减少,部分病人有出血倾向。重症病例可发生严重内出血、休克等[1-4]。登革病毒属于黄病毒科黄病毒属,有四种血清型(DENV1-DENV4),是全球分布最广的虫媒病毒之一,目前有100多个国家或地区报告过登革热病例,并且在一些热带、亚热带国家或地区呈地方性流行[1-4]。
登革病毒的传播循环分为城市循环和丛林循环,城市循环的主要传播媒介为埃及伊蚊和白纹伊蚊,病毒以蚊-人-蚊途径通过带病毒蚊虫叮咬人传播[1,4]。我国自1978年海南出现登革热疫情以来,南方的广东、云南、广西、福建、浙江等省均暴发过规模不等的本地登革热疫情[5]。福建省首次登革热本地暴发疫情出现于1999年,随后每年报告的登革热病例以输入性病例为主,偶尔会出现输入病例引起的本地流行[5-6]。近10年来福建省先后出现过几次规模较大的本地登革热疫情,分别是2004年福州台江、2007年莆田涵江、2014年南平建瓯疫情[5-8]。
2015年9月中旬福建省莆田市涵江区和福州市鼓楼区先后报告了本地聚集性登革热病例,分别为登革2型(DENV2)和登革1型病毒(DENV1)引起,随后10月初福州市晋安区也出现DENV2毒株引发的聚集性病例。为了追溯这些毒株可能的传播来源以及调查毒株之间的遗传联系,本文对2015年福建省部分本地登革热病例样品进行了病毒分离及基因组序列测定,分析分离株全长编码区序列与国内外流行毒株的种系发生关系,为福建省今后的登革热防控提供准确的病原学证据。
1 材料与方法
1.1病例资料及血清标本 病例资料来自“中国疾病预防控制信息系统”福建省2015年报告的本地登革热病例。本地感染病例是指发病前14 d内未离开过所在县(区),或未到过登革热疫情报告的地区,其感染地点位于报告地区。患者血清标本来自福州市内部分医院送检的疑似登革热标本,以及各设区市送检的登革热监测复核样品。实验室初筛结果显示,部分样品血清学检测登革病毒NS1抗原阳性或登革病毒特异IgM抗体阳性。
1.2登革病毒核酸检测及血清型鉴定 按QIAGEN公司试剂盒(QIAamp Viral RNA Mini Kit)操作手册提取患者血清样品总RNA,采用通用型实时荧光RT-PCR试剂盒检测登革病毒RNA(上海之江生物科技股份有限公司),随后用登革病毒1-4型实时荧光RT-PCR试剂盒对阳性样品进行血清型鉴定(上海之江生物科技股份有限公司)。
1.3病毒分离 登革病毒核酸阳性血清样品经过适当稀释,接种单层C6/36细胞,每日观察细胞,待出现细胞病变(CPE),收集培养上清,-80 ℃冰箱保存待检。
1.4基因组片段扩增 采用分段扩增的策略分别扩增登革病毒的基因组片段,扩增引物序列参考文献略做调整[9-10]。每个片段长度为1.0-1.5 kb,共10个片段,覆盖全长基因组。扩增试剂盒为大连TaKaRa公司PrimeScript One Step RT-PCR Kit Ver.2 (Dye Plus),扩增产物送铂尚生物技术(上海)有限公司测序。
1.5序列分析 对测序数据进行拼接、组装,得到毒株的全长基因组。分别从Virus Pathogen Resource(ViPR)数据库中选取基因型确定的登革病毒参比毒株、近几年福建省及国内外流行毒株约40株基因组序列(https://www.viprbrc.org),特别是近几年福建省输入病例来源地国家的毒株,以BioEdit软件(V7.0.5)进行序列比对和编辑。利用MEGA6.0软件,选择病毒全长编码区序列,以最大似然法(Maximum Likelihood)进行种系发生分析[11]。
2 结 果
2.1福建省2015年本地登革热疫情概况 2015年1-8月,福建省累计报告输入性登革热病例32例。9月中旬,莆田市和福州市先后出现本地登革热疫情,共报告41例。其中8例来自莆田市涵江区,其余33例来自福州市鼓楼区、晋安区和仓山区。初步流行病学调查显示,这些病例与当年已经确诊的输入性病例无关联性,发病时间集中在9月中旬至10月中旬(图1)。根据报告病例的流行病学信息及部分患者病毒血清型分型结果综合判断,大致分为4起聚集性病例(簇1-簇4),见表1。其中簇1为莆田涵江病例,共8例,由DENV2引起。簇2为福州鼓楼病例,来自同一社区,先后报告23例,由DENV1引起。簇3为福州晋安区病例,由DENV2感染,共报告病例7例。簇4的3例病例来自福州仓山区和鼓楼区,患者都供职于同一家公司,感染的病原体为DENV1。从时间顺序来看,簇1和簇2几乎同时报告,而簇3和簇2在时间和空间存在部分重叠(图1)。
2.2本地登革热疫情相关病毒分离株 患者血清样品接种C6/36细胞,部分培养瓶观察到细胞病变,经实时荧光RT-PCR分型鉴定,共有DENV1毒株9株,DENV2毒株8株,分别来自上述4起聚集性疫情,相关信息见表2。
表1 福建省2015年本地登革热病例汇总
Tab.1 Summary of indigenous dengue cases in Fujian province, 2015
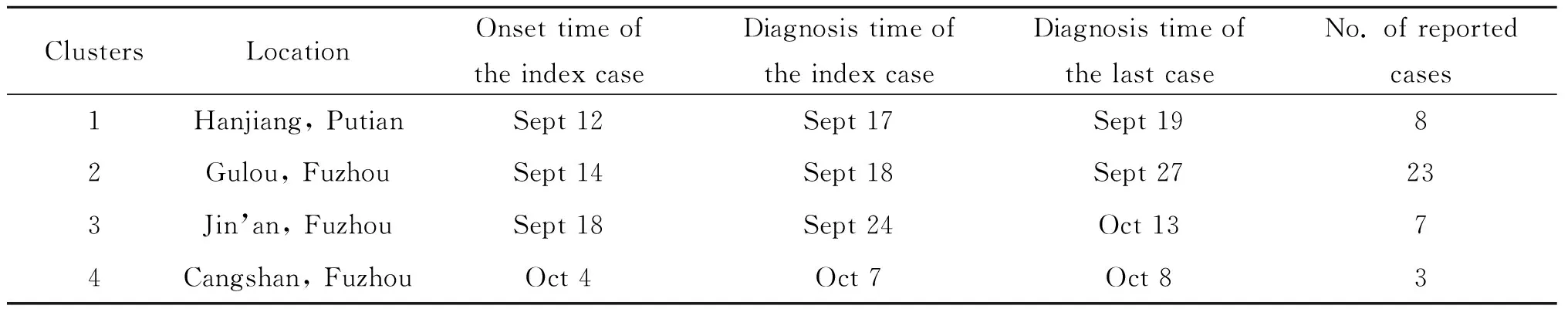
ClustersLocationOnset time of the index caseDiagnosis time of the index caseDiagnosis time of the last caseNo. of reported cases1Hanjiang, PutianSept 12Sept 17Sept 1982Gulou, FuzhouSept 14Sept 18Sept 27233Jin’an, FuzhouSept 18Sept 24Oct 1374Cangshan, FuzhouOct 4Oct 7Oct 83
表2 福建省2015年本地登革病毒分离株信息汇总
Tab.2 Summary of indigenous dengue viruses isolated in Fujian province, 2015

Strain IDSerotype-genotypeLocationOnset timeDiagnosis timeClusterFull-length(bp)DF150301D2-CosmopolitanHanjiang, PutianSept 16,2015Sept 17,2015110 668DF150303D2-CosmopolitanHanjiang, PutianSept 15,2015Sept 17,2015110 671DF150305D2-CosmopolitanHanjiang, PutianSept 14,2015Sept 17,2015110 657DF150308D2-CosmopolitanHanjiang, PutianSept 12,2015Sept 19,2015110 666DF150336D2-CosmopolitanJin’an, FuzhouOct 3,2015Oct 7,2015310 641DF150361D2-CosmopolitanJin’an, FuzhouOct 5,2015Oct 8,2015310 650DF150407D2-CosmopolitanJin’an, FuzhouOct 10,2015Oct 13,2015310 641DF150409D2-CosmopolitanJin’an, FuzhouOct 10,2015Oct 13,2015310 667DF150275D1-G1Gulou, FuzhouSept 14,2015Sept 18,2015210 654DF150278D1-G1Gulou, FuzhouSept 16,2015Sept 18,2015210 658DF150328D1-G1Gulou, FuzhouSept 25,2015Sept 30,2015210 659DF150340D1-G1Gulou, FuzhouSept 19,2015Sept 22,2015210 644DF150344D1-G1Gulou, FuzhouSept 21,2015Sept 24,2015210 658DF150348D1-G1Cangshan, FuzhouSept 23,2015Sept 29,2015210 655DF150364D1-G1Cangshan, FuzhouOct 4,2015Oct 7,2015410 658DF150365D1-G1Cangshan, FuzhouOct 6,2015Oct 8,2015410 653DF150366D1-G1Gulou, FuzhouOct 6,2015Oct 8,2015410 663
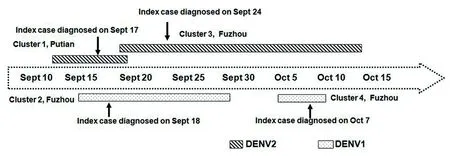
图1 福建省2015年本地登革热病例时间轴Fig.1 Timeline of indigenous dengue cases in Fujian province, 2015
提取病毒RNA,分别扩增病毒基因组节段,得到的序列数据经过拼接,获得病毒全长基因组序列,长度在10 641-10 671 bp之间。将病毒基因组序列在GenBank数据库进行BLAST分析,所有新分离DENV1毒株与G1基因型毒株的核苷酸相似度最高,所有DENV2与大都市基因型(D2-Cosmopolitan)毒株核苷酸相似度最高,见表2。
2.3新分离毒株与国内外流行毒株的种系发生分析 通过对9株新分离DENV1毒株与41株国内外流行株基因组序列进行比对,用全长编码区10 179 bp构建种系发生树。图2显示,这9株病毒之间具有较高的同源性,聚集在种系发生树的顶端,位于G1基因型的分支。遗传关系最近的参比毒株为2014年斯里兰卡分离株DK87(GenBank登录号:KP398852)。福建省2004年DENV1分离株FJ231/04(DQ193572)及2012年分离株DF1203(KC759167),与这9株病毒虽然同属于G1基因型,但分别位于G1基因型的另外两个分支,显示了它们之间的遗传差异性。而其它参考毒株,包括国内部分省市1991-2015年分离的DENV1毒株、2009-2015年福建省主要输入病例来源地的国外流行株,与这9株病毒则存在明显的差异,位于种系发生树的不同分支,分属于不同的基因型(图2)。
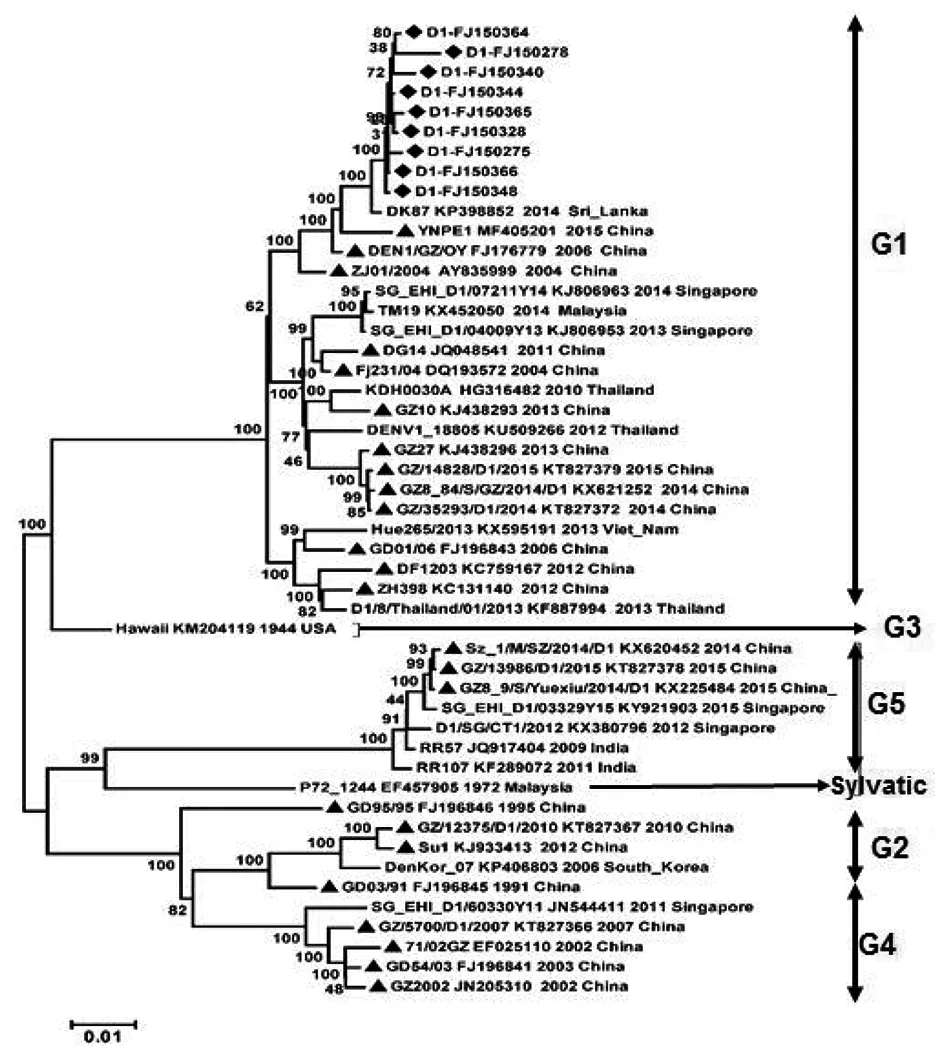
图2 福建省2015年部分本地登革1型病毒分离株与国内外流行株的遗传联系Fig.2 Phylogenetic relationship of global circulating DENV1 strains with indigenous isolates in Fujian province, 2015
新分离的DENV2共8株,将其基因组序列与40株国内外基因组序列比对之后,选取全长编码区10 176 bp,构建种系发生树(图3)。尽管这8株病毒都属于DENV2的大都市基因型,其中4株莆田毒株之间相似度高,聚集成簇,位于该基因型的顶端分支。而4株福州毒株也表现出高度相似性,聚集成另外一簇,但位于该基因型的底端分支。以上结果显示,莆田毒株与福州毒株之间存在明显的遗传异质性。从种系发生树上看,与莆田毒株遗传关系最近的参比毒株是2014年马来西亚毒株TM39 (KX452018)及2015年8月由菲律宾输入福建的毒株PH-CN77-15(KU517847)。而与福州毒株遗传关系最接近的参比毒株则是2011年印度分离株RGCB921/2011(KY427085)。唯一的福建省DENV2既往毒株,1999年福州本地分离株FJ11/99(AF359579)与这两簇病毒存在不同程度的差异。其它参比毒株,分属不同的基因型,位于种系发生树的不同分支,与这8株病毒存在不同程度的遗传差异(图3)。
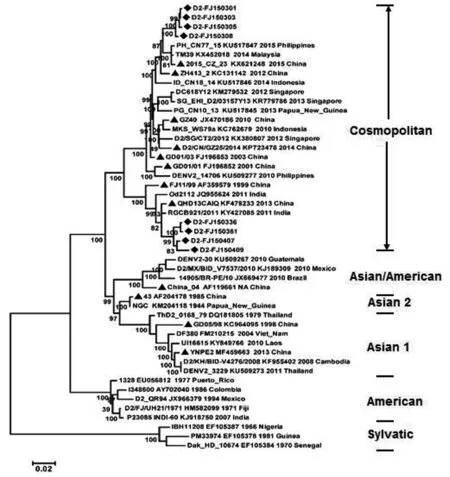
图3 福建省2015年部分本地登革2型病毒分离株与国内外流行株的遗传联系Fig.3 Phylogenetic relationship of global circulating DENV2 strains with indigenous isolates in Fujian province, 2015
3 讨 论
过去数十年里,登革热在全球发病率增长超过30倍,成为分布最广的虫媒病毒病之一。随着主要传播媒介伊蚊孳生地的地理范围不断扩大,日益增长的国际贸易、跨境旅游、森林破坏、城市化以及气候变化等因素,也促进了登革病毒的传播[1-4]。登革热主要发生在热带和亚热带国家和地区,近几年里,法国、葡萄牙及日本等温带国家也出现了本地登革热疫情,显示登革热流行地区正在由地方性区域逐渐向非地方性区域扩散[12-15]。我国属于登革热非地方性流行国家,全国报告病例以输入性病例为主,在蚊媒活跃季节也出现因输入病例引起的本地暴发流行[5]。在这样的国际环境和流行趋势下,福建省近10年来不仅存在来自登革热地方性流行地区的输入性登革热病例,同时由于地处亚热带区域,全省大部分地区都具备适合白纹伊蚊生长的温度和湿度,因此自1999年以来,还陆续出现了几起本地登革热暴发疫情,其中也包括2015年出现聚集性本地病例的莆田市和福州市[6-8]。
不同于以前的几次本地登革热疫情,福建省2015年的疫情表现为多疫点发病、多种血清型同时流行的特点[6-8]。在前后1 d之内,相隔近100 km的莆田涵江区和福州鼓楼区同时报告本地登革热病例,且分别为DENV2和DENV1感染。簇1中大部分病例来自同一个行政村,簇2中病例集中在一个社区,根据时间、空间和血清型综合判断,簇1和簇2病例应该是互相独立的事件。簇3病例感染的是DENV2,簇4的病原体为DENV1,如何揭示它们与簇1、簇2病例之间的联系,必须通过解析病原体基因组序列,获得病原学证据。因此,本研究测定了相关毒株的全长基因组序列,并进行种系发生分析。根据DENV1构建的种系发生树(图2),簇2和簇4病例中分离毒株的基因组序列显示出较高的同源性。虽然簇4的病例居住地不属于簇2病例所在社区,但几例病例就职于同一家公司,其工作场所距离簇2病例的社区不到3公里,因此尽管二者发病时间无交集,从基因组序列分析推测,簇4病例可能为簇2疫情的延续。而根据DENV2构建的种系发生树(图3),簇1和簇3病例的分离株各自聚集成簇,虽然同属于大都市基因型,仍然显示了两组基因组序列具有明显的差异,表明两起疫情的病原体之间无遗传关联性。
我国属于登革热非地方性流行地区,近10多年来,福建省输入性登革热病例来源地包括东南亚、南亚、非洲、大洋洲、南美洲等国外疫区,也出现过从国内流行区返回的输入性病例[5-8]。在出现本地登革热病例之后,由于流行病学调查结论不支持当年输入性病例与本地病例之间的关联性,为了追溯本地病例的可能来源,阐明新分离株与国内外流行毒株之间的遗传联系,我们测定了这些本地分离株的基因组序列,并分别选取了约40株参比毒株进行种系发生分析。在种系发生树上,新分离的9株DENV1毒株与2014年斯里兰卡分离株DK87(KP398852)同源性最高,显示引起簇2和簇4病例的DENV1毒株可能的共同输入来源为斯里兰卡。而簇1的4株DENV2毒株,与2014年马来西亚毒株TM39 (KX452018)同源性最高,簇3的4株DENV2毒株,则与2011年印度分离株RGCB921/2011(KY427085)最接近,因此它们的输入地可能分别为马来西亚和印度。福建省近几年登革热输入病例来源地名单中,也包括以上这些国家。
在登革病毒传播过程中,病毒要经历两个阶段,即在人体中扩增的内潜伏期和在蚊媒中复制的外潜伏期,分别为3~14 d和5~12 d,而患者发病之后还有5~6 d的病毒血症期[1,4]。处于病毒血症期的患者容易被蚊媒叮咬将病毒扩散,经过外潜伏期以后的蚊媒终生具有感染性,可以感染多人[1,4]。因此,一旦在登革热地方性流行区域之外,出现了本地登革热疫情,往往由于人群普遍无免疫力而导致短时间内涌现大量病例。例如,始于2012年10月的葡萄牙马德拉群岛本地登革热疫情,前后历时7个月,病例超过2 000例,有13个欧洲国家报告输入病例[12-13]。2014年日本东京的本地登革热疫情,历时两个月,报告病例160例,新卡里多利、英国、澳大利亚报告输入病例[14]。相对来说,2015年福建省的本地登革热疫情,经历的时间短、涉及病例少,可能原因是莆田和福州曾经发生过登革热疫情,相关部门具有处理类似疫情的经验,能够密切配合进行病例管理,积极开展蚊媒控制,从而有效防止了病毒的进一步扩散。
综上所述,福建省2015年本地登革热疫情涉及4起聚集性病例,分别为DENV2和DENV1感染引起。根据部分病毒分离株全长基因组序列分析,簇2和簇4的DENV1毒株高度同源,可能输入来源为斯里兰卡。簇1和簇3的DENV2毒株存在明显差异,可能输入来源分别为马来西亚和印度。因此,通过对2015年本地登革热毒株的病原学特征分析,提示福建省今后登革热防控的重点依然是加强对境内外输入病例的监测及管理,同时在蚊媒活跃季节积极开展本地蚊媒控制,提前切断登革病毒在本地的传播链,才能避免输入登革病毒的扩散蔓延。
利益冲突:无
