冰川退缩地15种丛藓科植物茎的比较结构学观察
2019-02-14艾菲热阿布都艾尼陈秋艳耿若楠吴玉环
艾菲热·阿布都艾尼 陈秋艳 王 虹* 耿若楠 吴玉环
(1.新疆大学生命科学与技术学院,乌鲁木齐 830046; 2.杭州师范大学生命与环境科学学院,杭州 310036)
新疆天山一号冰川退缩地是典型的高山冻原植被带[1],该地区海拔高,一般在3 100~3 900 m,紫外辐射强烈、风大、寒冷[2],气候和环境非常严酷,生长的植物大多特别矮小,而藓类植物[3~5]以其特有的非常矮小和垫状丛生的特点成为这里的最主要的优势植物类群[6~7]。
丛藓科(Pottiaceae)属于真藓亚纲(Bryidae)顶蒴单齿亚类(Acrocarpi-Haplolepideae)丛藓目(Pottiales),该科植物是全世界已知藓类植物中属的数量最多的一个科,有76属,2 000多种,在我国共报道有39属,179种和变种[8]。丛藓科植物主要分布在温带地区,多属于旱生性藓类植物,是沙漠中固定沙丘结皮层的主要植物类群,具有非常重要的生态学价值[9]。
而Zander对丛藓科植物的分类系统方法[10]也为国内外大多数学者所普遍接受[8]。目前对于国内不同地区的丛藓科植物的研究有王晓蕊等通过扫描电镜对16种国产丛藓科(Pottiaceae)植物蒴齿的形态特征进行了观察,发现丛藓科植物在齿条形状等方面有差异,表现出了不同属、种间的遗传分化[11];黄士良等系统地研究了18种丛藓植物孢子萌发和原丝体发育过程,分析了其发育过程中的不同特征,将这18种原丝体发育类型划分成真藓型、光藓型、虎尾藓型和蓑藓型4种孢子萌发型[12];寇瑾等利用光学显微镜对13种丛藓科植物叶细胞的疣和乳突进行观察,认为疣和乳突是丛藓科植物叶细胞表面的稳定结构,可借此来进行丛藓科属间和种间的划分[13];随着分子技术的发展,有学者开始利用叶绿体基因rps4等分子研究的方法[14~15],探索丛藓科疑难属种的问题。
王虹等对新疆几种丛藓科植物的结构做过一些解剖学研究[16~18],探讨了茎、叶形态结构与旱生环境的关系,并且对其他生长在一号冰川地区的藓类植物的茎也做了微结构的研究[19~20],詹琪芳、刘倩、Isawo Kawai等人也对不同属种的藓类植物的茎做了一系列的结构比较研究[21~24],均指出这些藓类植物茎的显微结构反映了其对环境的适应性,并且具有分类学意义。然而专门对高寒冰缘区丛藓科植物茎结构的研究还未见报道,本文将对乌鲁木齐河源区产的15种丛藓科植物茎的解剖结构进行研究,应用石蜡切片和电镜扫描技术,比较不同种的丛藓植物茎的微结构差异,并探讨其微结构对环境的适应性,以期为丛藓科植物分类及生态适应性的研究提供新的依据。
1 实验材料与方法
本实验中的实验材料采自新疆天山一号冰川地区,凭证标本存放于新疆大学生命科学与技术学院植物标本馆(XJU)和杭州师范大学标本馆(HTC),标本详细信息见表1。
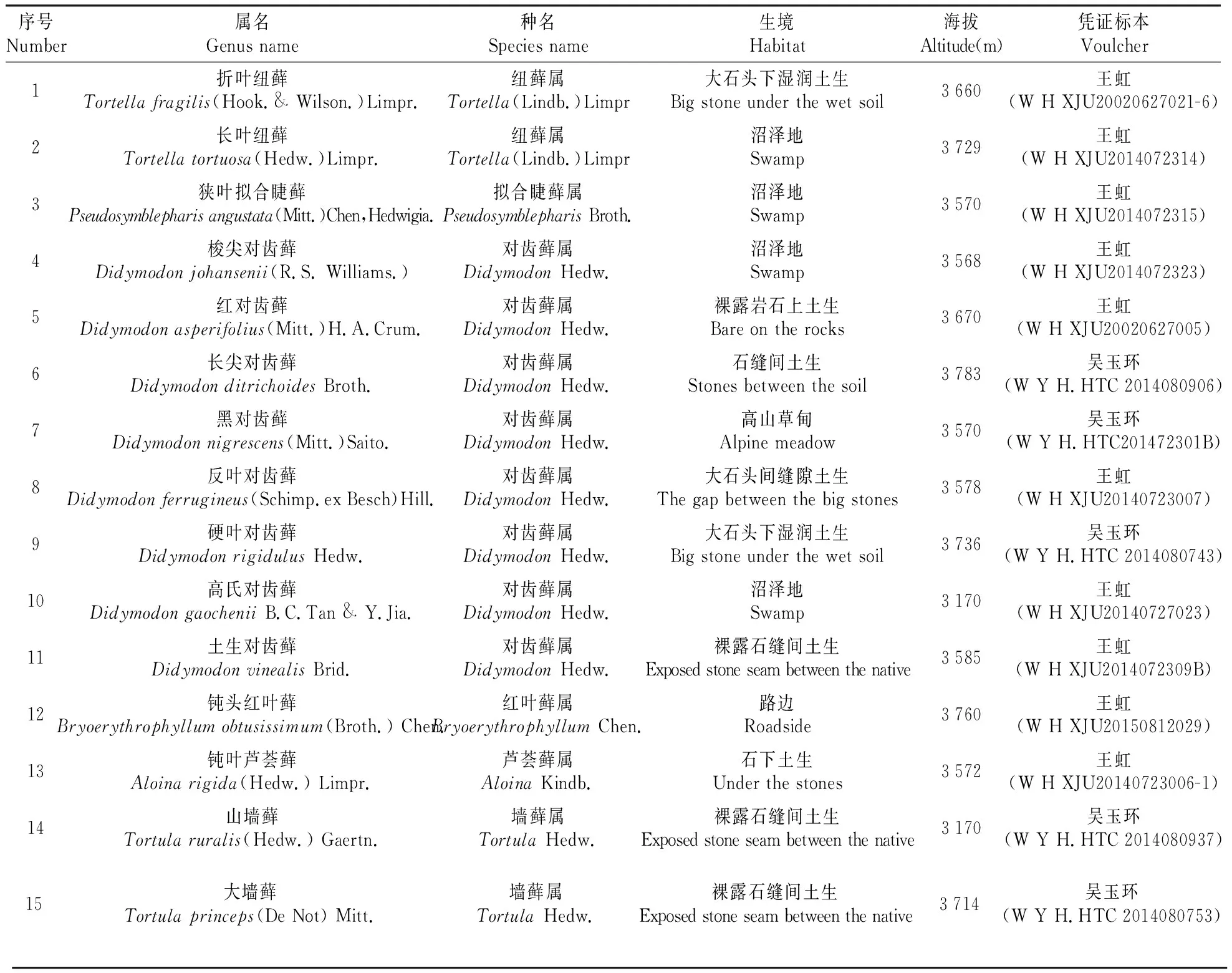
表1 实验材料及来源
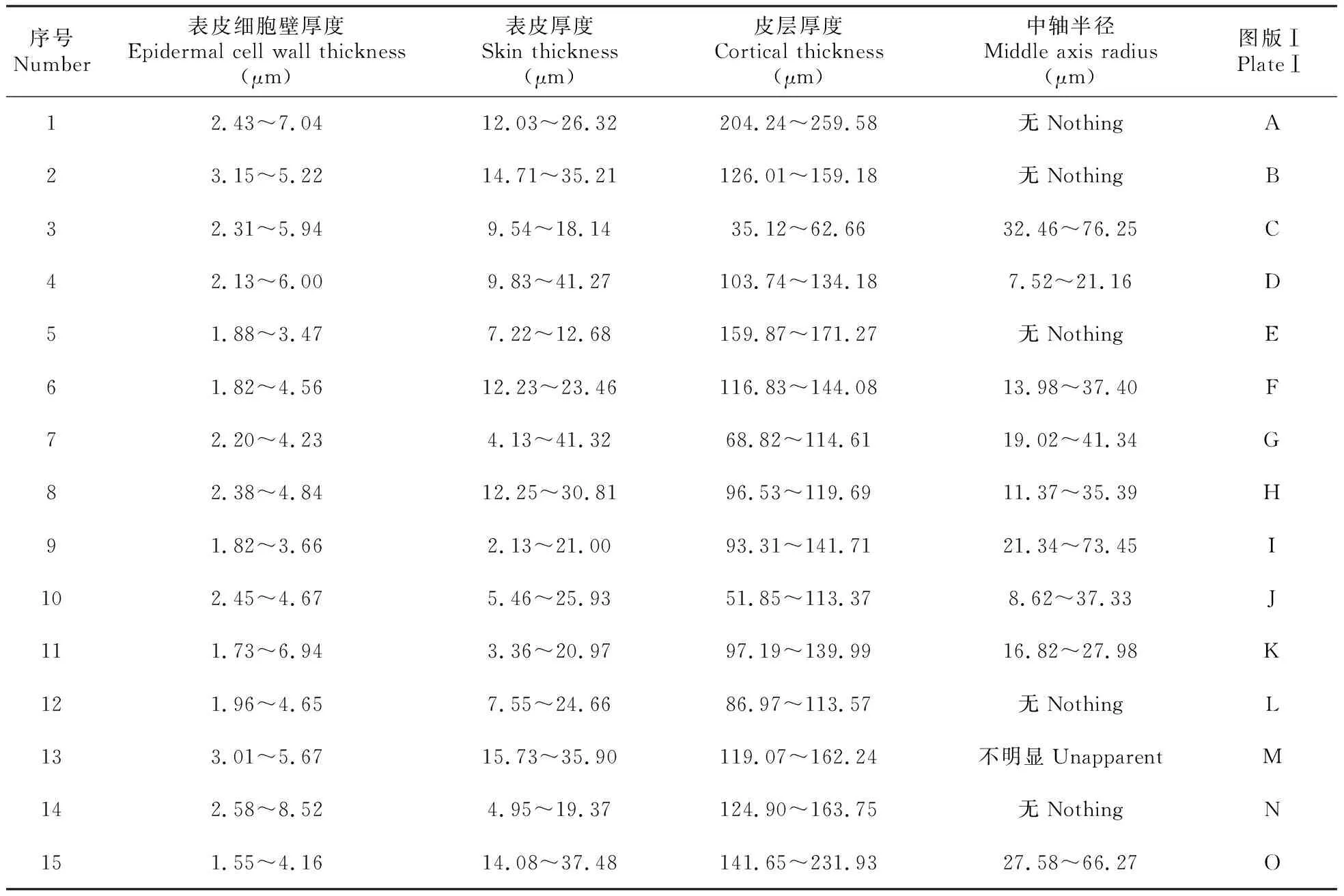
表2 15种丛藓科植物茎横切面结构比较
将标本进行表面泥土的简单清洗后,用POLARON CA7615型超声波洗净,用FAA进行固定。分别取一部分实验材料进行酒精梯度脱水、包埋、切片(厚度7 μm),番红染色后封片,Nikon E200显微镜下进行显微照相。每种标本随机选取12个横切面,测量茎的表皮细胞壁厚度、细胞壁厚度、皮部厚度、中轴半径,记录每种标本的茎横切面特征(表2)。将剩余的另一部分标本用95%酒精脱水、制片,用CARBON ACCESSORY喷镀仪镀金,在德国产LEO1430VP型扫描电镜下扫描观察并照相。
2 观察结果
2.1 折叶纽藓T.fragilis(Hook.& Wilson.) Limpr.
茎横切面近圆形,表皮细胞壁加厚并向外凸起;外皮部细胞1~3层,细胞壁厚薄不均;内皮部细胞3~4层,细胞壁薄,角隅加厚明显;无中轴分化。茎干燥时表皮细胞较长,收缩强烈,呈细条状排列,细胞壁表面凹陷较深,其上具细小颗粒及不规则的细小孔状窝点(图版Ⅰ.A;图版Ⅱ.A)。
2.2 长叶纽藓T.tortuosa(Hedw.) Limpr.
茎横切面近卵圆形,细胞中内含物较多;表皮细胞壁厚,向外凸起;外皮部细胞1~2层,细胞壁增厚明显;内皮部细胞3~5层,细胞壁稍薄;无中轴分化。茎表皮细胞较短,细胞壁表面粗糙,纹饰不规则(图版Ⅰ.B;图版Ⅱ.B)。
2.3 狭叶拟合睫藓P.angustata(Mitt.) Chen,Hedwigia.
茎横切面近不规则五角形,细胞均具浓厚的内含物;表皮细胞壁强烈加厚,外皮部细胞1~2层,细胞壁强烈加厚;内皮部细胞2~3层,细胞壁厚;中轴分化明显,细胞小,所占比例较小。茎干燥时表皮细胞呈粗条状整齐排列,细胞壁表面凹陷深浅不一,其上具大量鳞片状纹饰及小窝点(图版Ⅰ.C;图版Ⅱ.C)。
2.4 梭尖对齿藓D.johansenii(R.S.Williams.)
茎横切面近圆形,细胞质浓厚;表皮细胞壁厚,呈扁平状排列,细胞质明显少于皮部细胞的;外皮部细胞1~2层,细胞壁稍厚,细胞质浓;内皮部细胞3~4层,角隅加厚,细胞壁厚薄不均,细胞质浓厚;具中轴分化,只有几个极小的细胞,所占面积也极少。茎干燥时表皮细胞呈宽竹节状,其表面向内不同程度的凹陷,具较多不规则纹饰及小窝点(图版Ⅰ.D;图版Ⅱ.D)。
2.5 红对齿藓D.asperifolius(Mitt.) H.A.Crum.
茎横切面近圆形,表皮细胞壁强烈增厚,外凸明显,细胞排列整齐,细胞腔小;外皮部细胞1~3层,细胞壁强烈加厚,细胞小;内皮部细胞3~5层,细胞壁厚,呈网状排列;无中轴分化。茎干燥时表皮细胞呈条状紧密排列,表面粗糙,凹陷深浅不一,其上具细小颗粒及小窝点(图版Ⅰ.E;图版Ⅱ.E)。
2.6 长尖对齿藓D.ditrichoides Broth.
茎横切面椭圆形,表皮细胞壁明显增厚,且向内凹陷,细胞呈扁平状排列;外皮部细胞1~2层,细胞壁厚;内皮部细胞3~5层,细胞壁薄;具中轴分化,中轴细胞小,所占比例也少。茎干燥时表皮细胞相对较长,整齐地紧密排列,细胞壁表面粗糙,其上具大量细小颗粒、细纵纹及小状窝点分布(图版Ⅰ.F;图版Ⅱ.F)。
2.7 黑对齿藓D.nigrescens(Mitt.) Saito.
茎横切面近不规则多边形,表皮细胞壁明显增厚并内凹,细胞大小不一,排列不整齐;外皮部细胞1~2层,厚薄不均;内皮部细胞1~2层;中轴分化明显,细胞相对大一些,所占比例也稍多,细胞壁明显角隅加厚。茎干燥时表皮细胞短,呈网状排列,细胞壁表面明显向内凹陷,表面粗糙,有较多的细小颗粒及小窝点分布(图版Ⅰ.G;图版Ⅱ.G)。
2.8 反叶对齿藓D.ferrugineus(Schimp.ex Besch) Hill.
茎横切面卵圆形,细胞质较淡;表皮细胞壁加厚;外皮部细胞2~3层,细胞壁厚薄不均;内皮部细胞为3~4层,细胞壁薄;中轴分化,细胞小,所占比例也少。茎干燥时表皮细胞规则排列,细胞壁表面较光滑,少量纹饰多分布在凸起的细胞壁上(图版Ⅰ.H;图版Ⅱ.H)。
2.9 硬叶对齿藓D.rigidulus Hedw.
茎横切面近长椭圆形,细胞排列整齐;表皮细胞壁加厚,细胞呈扁平状排列;外皮部细胞1~2层,细胞壁厚;内皮部细胞2~3层,细胞壁薄,角隅加厚;中轴分化明显,细胞相对其他种植物的大些,所占比例也较大,细胞壁角隅加厚明显。茎干燥时表皮细胞壁凹陷相对较浅,其上有片状蜡质(图版Ⅰ.I;图版Ⅱ.I)。
2.10 高氏对齿藓D.gaochenii B.C.Tan & Y.Jia.
茎横切面不规则多边形,细胞镶嵌排列,内有一些内含物;表皮细胞壁加厚并向内凹陷;外皮部细胞1~2层,细胞壁厚薄不均;内皮部细胞2~4层,镶嵌排列,细胞壁薄,角隅加厚;中轴分化,所占比例少。茎干燥时表皮细胞收缩不规则,细胞壁表面很粗糙,其上有大量的鳞片状蜡质堆积及小孔窝点分布(图版Ⅰ.J;图版Ⅱ.J)。
2.11 土生对齿藓D.vinealis Brid.
茎横切面近圆形,细胞中有内含物;表皮细胞壁厚薄不匀,多向内凹陷;外皮部细胞1~2层,细胞壁厚;内皮部细胞2~3层,细胞呈镶嵌排列;具中轴分化,细胞很少,所占比例也少,角隅加厚明显。茎干燥时表皮细胞较短,细胞壁稍粗糙,厚薄不匀,细胞表面有不规则的小孔或小窝点分布(图版Ⅰ.K;图版Ⅱ.K)。
2.12 钝头红叶藓B.obtusissimum(Broth.) Chen.
茎横切面近卵圆形,细胞排列整齐;表皮细胞壁外凸;外皮部细胞1层,细胞小,细胞壁厚;内皮部细胞4~6层,细胞壁角隅加厚;无中轴分化。茎干燥时表皮细胞呈不规则锁链状,细胞短小,细胞壁不均匀加厚,细胞壁表面较粗糙,其上有大量细小颗粒及小窝点分布(图版Ⅰ.L;图版Ⅱ.L)。
2.13 钝叶芦荟藓A.rigida(Hedw.)Limpr.
茎横切面椭圆形,表皮细胞壁厚,排列不整齐,细胞腔大且有内含物;外皮部细胞为1层,细胞壁厚薄不均;内皮部细胞为3~6层,细胞壁薄,角隅加厚;中轴分化不明显。茎干燥时表皮细胞呈长条状排列,凹陷的细胞壁上有较多的细纵纹及一些细小的颗粒分布(图版Ⅰ.M;图版Ⅱ.M)。
2.14 山墙藓T.ruralis(Hedw.) Gaertn.
茎横切面近卵圆形;表皮细胞壁加厚;外皮部细胞为1~2层,细胞壁厚;内皮部细胞为4~6层,细胞壁角隅加厚明显,细胞大小不规则;无中轴分化。茎干燥时表皮细胞呈短竹节状,细胞壁不均匀加厚,表面略向内凹陷,其上表面具细小颗粒及小窝点分布(图版Ⅰ.N;图版Ⅱ.N)。
2.15 大墙藓T.princeps(De Not) Mitt.
茎横切面近圆形;表皮细胞壁加厚;外皮部细胞为1~2层,细胞壁厚薄不均;内皮部细胞为3~5层,细胞排列紧密;中轴分化明显,细胞细小,细胞壁角隅加厚。茎干燥时表皮细胞呈规则长方形网状整齐排列,细胞壁表面凹陷程度较为均一,其上具颗粒状附属物(图版Ⅰ.O;图版Ⅱ.O)。
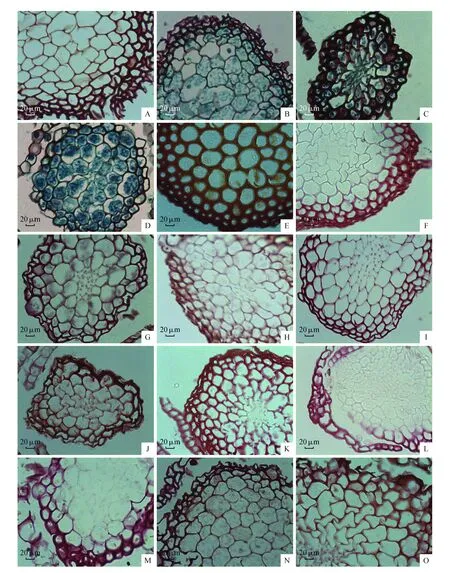
图版Ⅰ A.折叶纽藓;B.长叶纽藓;C.狭叶拟合睫藓;D.梭尖对齿藓;E.红对齿藓;F.长尖对齿藓;G.黑对齿藓;H.反叶对齿藓;I.硬叶对齿藓;J.高氏对齿藓;K.土生对齿藓;L.钝头红叶藓;M.钝叶芦荟藓;N.山墙藓;O.大墙藓Plant Ⅰ A.T.fragilis(Hook.& Wilson.)Limpr.; B.T.tortuosa(Hedw.)Limpr.; C.P.angustata(Mitt.)Chen,Hedwigia.; D.D.johansenii(R.S. Williams.); E.D.asperifolius(Mitt.)H.A.Crum.; F.D.ditrichoides Broth.; G.D.nigrescens(Mitt.)Saito.; H.D.ferrugineus(Schimp.ex Besch) Hill.; I.D.rigidulus Hedw.; J.D.gaochenii B.C.Tan & Y.Jia.; K.D.vinealis Brid.; L.B.obtusissimum(Broth.) Chen.; M.A.rigida(Hedw.) Limpr.; N.T.ruralis(Hedw.) Gaertn.; O.T.princeps(De Not) Mitt.
3 讨论与分析
3.1 冰川退缩地的15种丛藓科植物茎结构的生态适应性
藓类植物茎的结构会因为生长的环境不同而表现出一定的差异[22],而其茎的表皮、皮层及中轴的细胞层数、大小、形状、内含物以及微形态特征都能反映出与环境的适应性[19]。在这15种丛藓科植物茎的细胞中都有较浓厚的细胞质或内含物,如长叶纽藓、狭叶拟合睫藓等,可能其细胞具有分泌功能和储存营养物质的功能[16],而这几个属植物茎的表皮细胞壁大多数都有不同程度的加厚,并且向内凹陷或者凸起,皮部和中轴细胞壁都具有明显的角隅加厚现象,细胞多为镶嵌排列;具中轴的细胞均呈不规则的五边至多边形等,这些特征都是各属植物适应高寒环境的共同特征[16~20]。使植物能灵活应对外界环境的变化;这些丛藓科植物茎的表皮细胞大部分都较短,呈网格状排列,能增强其支撑能力。生长在裸露岩石上土生的红对齿藓,虽然无中轴的分化,但其茎细胞的细胞壁均为强烈加厚,细胞排列紧密,这样既可以起到很好的支撑作用,又可以使植物细胞充分储存水分并且能够防止水分蒸发;而狭叶拟合睫藓茎的细胞壁较薄,皮部细胞大,中轴非常小,这可能是其生长在沼泽的湿土中,水分充足,也不需要很强的支撑力度就可以很好的应对不利的环境而生存;而生长在岩石窝中的硬叶对齿藓茎的细胞壁虽然加厚不是很强烈,但是细胞壁基本上都有角隅加厚现象,表皮细胞呈扁平状排列,中轴细胞是这15种丛藓科植物中最大的,所占比例也较大,其表皮细胞壁上有片状蜡质,这些结构可以使植物维持细胞的形态及一定的保水作用,帮助植物体在恶劣的环境下生存,水分的散失和风雪、强光对其的伤害也相对小些[25~26]。通过研究丛藓科植物茎的微结构特征,可帮助我们较全面合理的解释藓类植物体在适应环境而产生某些特征差异,而植物茎结构对环境的适应能力,也使藓类植物能更好的应对干旱、高寒等恶劣环境对其造成的伤害[18~20]。
3.2 冰川退缩地的15种丛藓科植物茎结构的分类学意义
同为高寒地区生长的15种丛藓科植物,其茎的横切面形状及大小存在一定的相似之处,如横切面形状在圆形和椭圆形之间;皮部均有内、外皮部之分,外皮部细胞大多数为1~2层,细胞小,细胞壁厚,而内皮部细胞大,细胞壁由外向内逐渐变薄;有的同属植物的茎均有中轴,有的大多数无中轴,说明了它们具有作为一个自然类群的共性。胡人亮总结了苔藓植物的茎解剖结构认为,在不同种间,细胞的大小、形状及细胞各层之间的比例不完全相同,并将苔藓植物茎的结构归纳为:(1)泥炭藓型;(2)水藓属型;(3)葫芦藓型;(4)拟油藓属型;(5)提灯藓属型;(6)毛枝藓属型;(7)金发藓属型这7种类型[26]。由此,本文中的15种丛藓科植物茎的结构可细分为7种类型:(1)水藓属型,如折叶纽藓、长叶纽藓、红对齿藓、山墙藓;(2)水藓属变型Ⅰ,如钝头红叶藓(皮部细胞纵向中心拉长呈放射状排列);(3)水藓属变型Ⅱ,如钝叶芦荟藓(皮部细胞在同一个细胞中,细胞壁厚度明显不同,向外侧细胞壁明显增厚,而向内侧细胞壁则非常薄);(4)葫芦藓型,如梭尖对齿藓、反叶对齿藓、土生对齿藓、大墙藓;(5)葫芦藓变型Ⅰ,如长尖对齿藓(皮部细胞具2~3层厚壁细胞);(6)葫芦藓变型Ⅱ,如黑对齿藓、硬叶对齿藓、高氏对齿藓(茎表皮细胞多为扁平状排列,细胞外壁凹凸不规则);(7)提灯藓属型,如狭叶拟合睫藓。在本文的实验中红对齿藓茎无中轴分化,而Zander报道该种具中轴[9],因此,藓类植物茎的横切形状及中轴有无的性状特征不是很稳定。但是这些丛藓科植物茎的微结构都有不同的特征差异,在茎表面的纹饰、表皮细胞壁厚度、表皮厚度、皮层厚度等方面均有一些明显及细微上的差别(图版Ⅱ,表2),具有潜在的分类学意义[19~23]。例如,同为墙藓属的山墙藓及大墙藓,它们的亲缘关系和很多特点均接近,一般不易区分,但对其茎做结构的解剖学观察则较容易区分:前者无中轴分化、表皮细胞呈短竹节状排列,而后者有中轴分化、表皮细胞呈网格状排列;同样,折叶纽藓和长叶纽藓均为纽藓属,茎均无中轴分化,其他特征也很接近,但前者茎细胞的细胞壁角隅加厚明显,表皮细胞较长,干燥时收缩强烈,呈细条状排列,细胞壁表面具细小颗粒及不规则的细小孔状窝点;后者茎细胞中内含物较多,茎表皮细胞较短,细胞壁表面粗糙,纹饰不规则。而对不同属的丛藓科植物,同样也可以用光镜和扫描电镜相结合的方法加以区分。如中轴的有无、皮层的厚度(细胞层数、所占比例等)、表皮细胞的形态(细胞大小、形状、细胞壁厚薄、表面纹饰)等,均可为藓类植物的分类及亲缘关系分析提供一些依据[16]。
致谢衷心感谢中国科学院植物研究所的吴鹏程研究员、内蒙古大学的白学良教授鉴定本文实验标本。
