格氏栲自然保护区22种常见木本植物的生态策略
2019-02-10徐鹏程游章湉纪宇皝周嘉聪张秋芳郑蔚刘小飞林伟盛杨智杰杨玉盛
徐鹏程,游章湉,纪宇皝,周嘉聪,张秋芳,郑蔚,刘小飞,林伟盛,杨智杰* ,杨玉盛
(1.福建师范大学 a.地理科学学院,b.湿润亚热带山地生态国家重点实验室培育基地, 福州 350007;2.中国科学院 华南植物园 a.退化生态系统植被恢复与管理重点实验室, b.广东省应用植物学重点实验室,广州 510650; 3.中国科学院大学,北京 100049;4.华东师范大学 生态与环境科学学院,上海 200062)
生态策略(Ecological strategy)指的是物种维持生存的方法[1],是生态学领域经久未衰的研究热点之一。对于植物而言,生存的方法则是调整自身性状(属性)适应环境以维持[2]。作为森林生态系统中主要的生产者,植物的生态策略直接影响种群及群落尺度内物种的聚集规律,进而对生态系统尺度下的植被演变过程、系统功能变化等方面产生影响[3-4]。因此,明确植物生态策略及其对群落构建机制的影响力,在解释群落演替过程、生态系统变化,甚至是气候变化下的植被响应等方面有重要意义[5-7]。
当前,最成熟的植物生态策略研究方法是Pierce等人[2]根据Grime[8]提出的Competitor(C)-Stress tolerator(S)-Ruderal(R)策略理论发展而来的全球植物竞争-忍耐-杂草型策略分类法。该方法以植物比叶面积(SLA)、叶干物质含量(LDMC)、叶面积(LA)3个核心叶功能性状之间的数量关系为标准,判断该植物个体或种群的生态策略。全球植物CSR策略分类法超越以往单一使用CSR对策或是功能性状谱系分析植物生态策略的研究思维,采用先分析物种功能性状谱系,再结合CSR对策归纳总结得出植物生态策略。目前,该方法已在多个领域的生态学研究中应用[9-12]。
然而,该方法在群落构建方面的应用却存在争议。Negreiros[13]、Paula[14]、Rosado[15]等人分别在物种策略分布集中的生境中验证CSR策略分类法对群落物种组成的解释。前两者支持了该方法,Rosado等人[15]的研究结果不支持CSR策略分类法,他认为需要在植物生态策略多样化的地区,但需要进一步验证该方法的适应性。
热带和亚热带是世界上物种共存方式最多样的地区[2],虽然中国研究者们已经在此区域开展了大量植物功能性状的工作[6,16],但并没有研究者进一步将分散性状整合为综合的生态策略,并将与群落构建机制相联系。格氏栲自然保护区位于福建省三明市,区内以林龄约为300年的常绿阔叶林为主,共有维管束植物110种,隶属于42科68属,其中壳斗科、茶科、松科、樟科、紫金牛科和冬青科植重要值大都在10以上,具多样化的植物生态策略类型[17]。本研究以格氏栲自然保护区常见木本植物为研究对象,通过总结叶功能性状变化规律,主要探讨2个内容:(1)格氏栲自然保护区常见植物的生态策略;(2)验证生态策略分类与该区群落构建结果的一致性。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于福建三明格氏栲自然保护区内(26°7′~26°10′N,117°24′~117°27′E),处于福建武夷山东伸支脉地带, 是低山丘陵地貌, 海拔一般在250~500 m。林下土壤主要为暗红壤,其次为红壤和紫色土。该区属中亚热带季风气候,多年平均气温19.5 ℃,多年平均降水量1 700 mm。
1.2 植物类型分析
本研究选取格氏栲保护区内22种常见木本植物,包括11种小乔木/灌木、11种乔木,隶属于10科28属(表1)。
表 1 格氏栲自然保护区22种木本植物基本特征Table 1 Basic characteristic of 22 woody plants in the C.kawakamii reserve

名称拉丁文名生活型生活史茜草树Aidia cochinchinensis Lour.小乔木/灌木常绿米槠Castanopsis carlesii (Hemsl.) Hay. var. carlesii乔木常绿甜槠Castanopsis eyrei(Champ.) Tutch.乔木常绿丝栗栲Castanopsis fargesii Franch.乔木常绿格氏栲Castanopsis kawakamii Hay.乔木常绿华南桂Cinnamomum austrosinense H. T. Chang乔木常绿青冈栎Cyclobalanopsis glauca(Thunb.) Oerst.乔木常绿树参Dendropanax dentiger(Harms) Merr.乔木常绿狗骨柴Diplospora dubia(Lindl.) Masam.小乔木/灌木常绿短梗幌伞枫Heteropanax brevipedicellatus Li乔木常绿矩叶鼠刺Itea oblonga Hand.-Mazz.小乔木/灌木常绿华南木姜子Litsea greenmaniana Allen小乔木/灌木常绿黄绒润楠Machilus grijsii Hance小乔木/灌木常绿野含笑Michelia skinneriana Dunn小乔木常绿短梗新木姜子Neolitsea brevipes H. W. Li小乔木/灌木常绿香港新木姜子Neolitsea cambodiana Lec. var. glabra Allen小乔木/灌木常绿木荚红豆Ormosia xylocarpa Chun ex L. Chen乔木常绿马尾松Pinus massoniana Lamb.乔木常绿黄牛奶树Symplocos cochinchinensis (Retz.) Wall小乔木/灌木常绿山矾Symplocos sumuntia Buch.-Ham. ex D. Don小乔木/灌木常绿木荷Schima superba Gardn. et Champ.乔木常绿光叶山矾Symplocos lancifolia Sieb. et Zucc.小乔木/灌木常绿
1.3 测定项目与方法
1.3.1 比叶面积、叶干物质含量的测定
从测定光合参数的植株上,随机选取健康未损坏的叶片10枚,用电子天平称取叶鲜重(LFW),随后使用WinRHIZO/WinFOLIA根叶分析系统(WinRhizo, Régent Instruments, Quebec, Canada)测定叶面积(LA),最后将叶片放于70 ℃烘箱中烘72 h至恒重,用电子天平称取叶干重(LDW)。计算比叶面积:SLA(mm2·mg-1)= LA(mm2)/LDW(mg),叶干物质含量:LDMC =LDW(mg)/LFW(mg)[18]。
1.3.2 叶片全碳和全氮含量的测定
将烘干至恒重的植物叶片研磨成粉末状,用Vario ELⅢ型元素分析仪(Elementar Analysensysteme GmbH, Hanau, Germany)测定单位质量叶碳含量Cmass(mg·g-1)和叶氮含量Nmass(mg·g-1),并计算单位面积叶碳含量Carea(mg·cm-2)和叶氮含量Narea(mg·cm-2)[18]。
1.3.3 叶片全磷含量的测定
将烘干至恒重的植物叶片研磨成粉末状,用HClO4-H2SO4消煮法处理后,连续流动注射分析仪(Skalar San++,Netherland)测定单位质量叶磷含量Pmass(mg·g-1),并计算单位面积叶磷含量Parea(mg·cm-2)[18]。
1.3.4 群落学调查的方法
通过查阅相关文献[17]以及前期的植被状况调查,在本区较为典型的格氏栲-马尾松群系、格氏栲-木荷群系和米槠-格氏栲群系设置临时样地,每个群系类型设置3个重复。野外调查采用相邻格子样方法,样方大小定为5 m×5 m。取样时按正方形布置样方,并由中心向四周逐渐扩展进行,以亚热带常绿阔叶林最小表现面积1 200 m2为取样面积[19]。在各样方内,对林木进行每木调查,记录每个个体的高度、胸径、冠幅和生活力等。同时,对样方内出现乔木幼树、幼苗、灌木、草本和藤本同样记载,并分别估测乔木层、灌木层和草本层的总盖度。
1.4 数据处理方法
用CSR分析工具‘StrateFy’对22种植物的生态策略计算分析[2]; 用Excel计算物种的相对重要值;用Spearman相关性检验生态策略与相对重要值的相关性;用非参数检验分析不同生态策略间植物叶功能性状和相对重要值的差异;以上分析采用SPSS 22.0以及xlsTAT完成,并使用sigmaplot完成作图。
2 结果
2.1 叶功能性状分布范围
叶功能性状在物种间普遍存在较大的差异。其中,叶面积(LA)、叶鲜重(LFW)与叶干重(LDW)的变化范围较大,分别为478.8~44 567.1 mm2、45.9~4 620 mg、19.73~1 384 mg。比叶面积(SLA)和叶干物质含量(LDMC)的变化范围则分别在5.509~64.381 mm2·mg-1,0.214~0.668。以面积为基础的碳(C)、氮(N)、磷(P)单位数量值相对较小,但也呈十倍的数量级变化,其中Carea:0.069~0.864 mg·mm-2,Narea:0.008~0.021 mg·mm-2,Pmass:0.001~0.011 mg·mm-2。
2.2 生态策略与性状组合
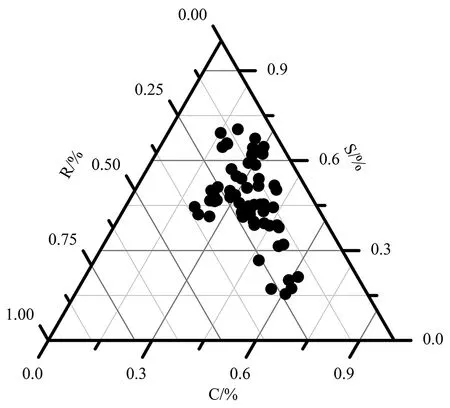
图 1 22种木本植物生态策略分类Figure 1 Calculation of ecological strategy among the 22 woody plants
以Pierce等人[2]划分的全球尺度下植物CSR策略划分方法,对22种木本植物CSR权衡策略分类归纳(图1),共分为七类策略,包含物种数目相对较多的有竞争-忍耐型/竞争-忍耐-杂草型(CS/CSR,32.3%)、忍耐/竞争-忍耐-杂草型(S/CSR3,34.1%)和竞争/竞争-忍耐型(S/CS,15.4%),包含物种数目相对较少的有竞争/竞争-忍耐-杂草型(C/CSR,6.2%)、竞争-忍耐型(CS,4.1%)、竞争-忍耐-杂草型(CSR,4.1%)、忍耐-杂草型(SR,3.8%)。其中C/CSR - CS/CSR - S/CSR,这三者形成了一个具备一定抗干扰能力(R)的C-S权衡策略轴,代表物种有树参(C∶S∶R = 62%∶18%∶20%)、茜草树(C∶S∶R = 40%∶39%∶21%)、华南木姜子(C∶S∶R = 27%∶51%∶22%)。同时,C/CSR-CS-S/CS这3个策略形成了抗干扰能力(R)较弱的C-S权衡策略轴,代表物种有黄楠(C∶S∶R=41%∶50%∶9%)、米槠(C∶S∶R = 19%∶66%∶15%)。而CSR策略植物是所有策略中各部分投资和能力最均衡的策略,代表物种有野含笑(C:S:R = 26%∶41%∶32%)。另外,SR策略植物则是S-R权衡策略的中间型,代表物种有光叶山矾(C∶S∶R = 20%∶45%∶35%)。
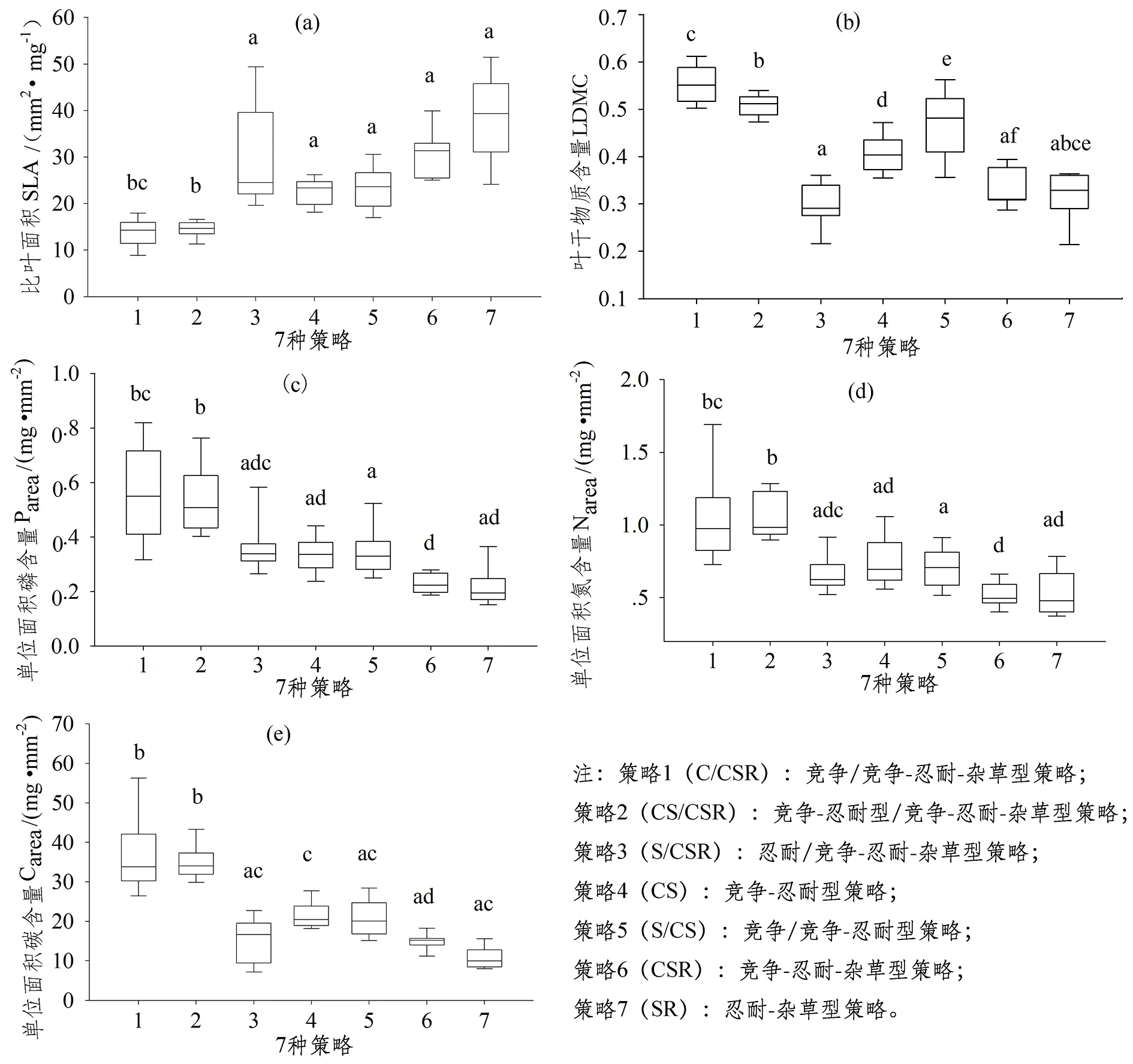
通过对各策略的比叶面积(SLA)、叶干物质含量(LDMC)、单位面积碳(Carea)、氮(Narea)、磷(Parea)含量比较(图2),结果表明格氏栲自然保护区植物在C-S策略轴上存在性状间经济权衡。从C/CSR-CS-S/CS这一个R部分投资较少的策略轴上看,LDMC、Carea、Narea、Parea性状平均值表现为S/CS≥CS>C/CSR或S/CS>CS>C/CSR,SLA则相反。在C/CSR-CS/CSR-S/CSR策略轴上,除了LDMC外,三者在其余的性状上皆没有表现出显著性差异,但在Carea、Narea性状平均值上,显示出C/CSR 表 1 22种木本植物生态策略与群落重要值的Spearman非参数相关性分析Table 1 Spearman Non-parametric correlation analysis of plant strategies and importance index 植物策略竞争能力(C%)压力忍耐能力(S%)抗干扰能力(R%)相对重要值-0.083∗∗0.139 1∗∗-0.208 注:**表示在0.01水平上显著相关。如表1所示,Spearman非参数相关性分析的结果表明,植物在群落中的相对重要值与代表物种竞争能力的C策略部分呈负相关关系,与代表物种忍耐能力的S策略部分呈正相关关系,与代表物种抗干扰能力的R策略部分不呈相关性。然而,不论是C策略部分还是S策略部分,与相对重要值都表现出了线性相关性拟合度较低的现象(图3),特别是在相对重要值较低的物种之间,竞争和压力忍耐能力都存在较大的变异。 通过全球植物生态策略方法(StrateFy),对格氏栲自然保护区常绿阔叶林22种常见木本植物进行生态策略分类,可分为七类:C/CSR策略、CS/CSR策略、S/CSR策略、CS策略、S/CS策略、CSR策略、SR策略。这与Pierce等人[2]所归纳的热带和亚热带阔叶林包含有多种生态策略的结果一致。保护区中存在2种主导策略:S/CSR和CS/CSR策略,所占的比例分别为34.1%和32.3%,S/CS策略所占的比例也达到15.4%,这与Pierce等人[2]的结果不完全一致,他认为亚热带阔叶林植物以CS-CSR策略为主。这是两者研究尺度不同的缘故,前者的研究以气候带为基本单位,每个气候带包括有多个演替阶段、多种生活型的森林植物,格氏栲自然保护区以林龄300年的常绿阔叶林为主,根据顶级演替理论,该区植物群落应处于演替的中后期阶段。相较于演替前期,处于演替中后期阶段的森林林冠郁闭度较高,光是大部分的幼苗甚至成年个体的限制性因子[20-22],压力忍耐力强的S策略植物有更多的优势,因而靠近S策略一端的植物有更大的生存机遇。 图 2 各生态策略植物功能性状箱Figure 2 Box plots of functional trait values among the seven strategies 功能性状是能够反映植物对外在环境适应的核心属性[23],将植物各功能性状的表现综合,就是量化植物对外在环境的适应性。本区七类生态策略的性状组合差异体现了C-S策略轴植物间存在的经济权衡。不论是C/CSR-CS-S/CS策略轴,还是C/CSR-CS/CSR-S/CSR策略轴,靠近C策略一端的植物在代表收益能力的SLA性状上大于靠近S策略一端的植物,而在代表构建成本的LDMC、Carea、Narea、Parea性状上小于靠近S策略的植物。事实上,这就是叶经济谱理论的核心——生长与生存的权衡[4,24]。C策略植物选择生长,以快速占领资源为主要目标,需要相对较大的比叶面积(SLA)提升光拦截效率,增加光合作用的生产收益,提高植物的相对生长率[25]。S策略植物优先考虑生存,以维持基本新陈代谢为主要目标,因而保持较高的干物质含量(LDMC)以维持叶片保卫构造的投入(防止水分流失),同时在叶片投资较多碳氮磷资源保留养分,应对不利条件[26]。本区地处中亚热带,光照充足,气候温暖湿润,土壤养分丰富且异质程度中等[27],适合多种策略类型植物生存,故不论是靠近C策略一端的植物还是靠近S策略一端的植物皆有。然而,本区森林已经历300年以上的自然演替,处于演替中后期阶段,种间种内对光资源的激烈竞争使各策略植物都不得不面对光资源不足的状况[28-29],这对采用高构建成本缓慢收益的S策略植物是极为有利的。这也从侧面解释了该区S策略以及靠近S策略一端的植物有着更多生存机遇,而C策略以及靠近C策略一端植物仅在林缘或林窗区域的现象。CSR策略是投资和收益最均衡的类型,SR策略则是位于S-R策略轴上的类型,然而,二者却一同表现出了高收益低投资的性状组合,这可能是植物R部分投资增加的缘故。R策略植物适应于干扰强烈的环境下,倾向于投入大量资源于繁殖,以抵消干扰对种群的影响,达到种群生存延续的目的,因此R策略植物需要比C策略更快速的收益,以维持强大的繁殖力[30]。然而,该区植被受中等程度以上的干扰时间可能要追溯200到300年前,不属于干扰强烈的环境,R部分投资增加可能是植物通过提升繁殖力适应演替中后期,种间种内竞争激烈,光资源不足的状况。 当前,基于全球植物生态策略分类对群落构建机制的研究依然存在争议,争议的焦点之一就是该方法是否能合理解释不同生境的群落构建结果[7,9,11,15]。本区植被属典型的亚热带常绿阔叶林,存在多种策略植物共存的状况[17]。从群落物种组成的结果而言,本研究与Rosado等人[15]的结论部分一致。虽然相对重要值与竞争能力(C)以及忍耐能力(S)存在显著相关性,但是线性拟合度较低,相对重要值最大的格氏栲并不是竞争能力或忍耐能力最好的物种,全球植物生态策略分类结果无法完全解释本区的群落物种组成。小尺度的物种共存机制的解释需考虑多种因素,光、温度、水分、土壤等任一因子的差异都可能对群落构建产生影响。此外,该区植被所处的自然演替阶段不同,群落组成同样存在巨大差异。处于演替初期阶段的植被,资源丰富但环境波动大,适合繁殖力强的R策略物种生存,此时相对重要值与繁殖能力(R)可能存在紧密的联系;处于演替前期阶段的植被,光资源充足且环境稳定,是依靠快速生长优先占据资源的C策略物种喜好的生境,此时相对重要值与竞争能力(C)的关系更紧密;而在本研究所处的演替中后期阶段的植被,环境稳定但光资源不足条件下,压力忍耐能力(S)应与相对重要值有着更紧密的相关。因此,本研究结果支持CSR分类法对解释亚热带演替中后期常绿阔叶林群落构建的有效性——随着植物竞争能力(C)减弱,压力忍耐能力(S)增强,物种的相对重要值表现出了升高的趋势。但在相对重要值低的物种之间,竞争和忍耐能力存在较大的变异,这可能是拟合度较低的原因。Shen[31]曾在广东鼎湖山的群落功能性状多样性研究中发现类似的现象,矮小幼树的群落功能多样性比高大成年树高,他认为这是因为矮小的幼树还未经过生境的过滤作用,故保留有较高的功能多样性,而高大的成年树经过生境过滤作用,功能性状趋向一致。本研究所测定的22种常见植物中相对重要值大的物种较高大,相对重要值小的物种较矮小且生于林下。在植物生长的过程中,生境的过滤使靠近S策略一端的植物生存下来,远离S策略一端的植物随着生长高度的增加逐渐被淘汰,因此,在相对重要值低的区间,物种生态策略存在较大的变异。 格氏栲自然保护区22种常见木本植物可以分为七类生态策略,竞争/竞争-忍耐-杂草型(C/CSR)、竞争-忍耐型/竞争-忍耐-杂草型(CS/CSR)、忍耐/竞争-忍耐-杂草型(S/CSR)、竞争-忍耐型(CS)、竞争/竞争-忍耐型(S/CS)、竞争-忍耐-杂草型(CSR)、忍耐-杂草型(SR)。而各生态策略之间的权衡主要发生在C-S策略轴上,靠近C策略一端的植物选择生长为主要目的,倾向于大比叶面积(SLA)与低叶干物质含量(LDMC)、单位面积碳(Carea)、氮(Narea)、磷(Parea)含量的性状组合,靠近S策略一端植物选择生存为主要目标,性状组合与C策略一端的相反。群落构建结果表明,植物生态策略分类法对群落重要值排序存在一定的解释度,靠近S策略一端的植物在演替中后期的常绿阔叶林有着更大的生存机遇。植物生态策略是植物为达到生存、生长、繁衍目的而采用的方法[32],是多种功能性状的综合性表现,Pierce等人[2]所提出的全球植物生态策略分类方法虽具备一定代表性且易操作,但还需要考虑到多种影响植物生态策略的性状,比如体现植物水分利用策略的叶片导水率(Kleaf)、膨压丧失点(Ψtlp),体现植物繁殖能力更新能力的种子质量(SM)、种子萌芽力(SR)等。另外,本研究仅选取格氏栲自然保护区常见木本植物,是否不同系统发育类群也有类似的变化规律还需进一步研究。2.3 生态策略与群落构建结果

3 讨论
3.1 基于叶功能性状的植物生态策略
3.2 基于生态策略的群落构建机制研究
4 结论
