巨噬细胞极化相关差异基因的筛选及其在脓毒症中的潜在治疗价值
2019-01-31杨蕊萍
杨蕊萍,钟 武
(西南医科大学附属医院急诊科,四川 泸州 646000)
脓毒症是机体对感染反应失调的一系列全身炎症反应综合征,严重时可导致器官功能障碍或循环衰竭,也是诱发脓毒性休克、多器官功能障碍综合征的重要原因[1-3]。脓毒症早期机体呈现出一种促炎状态,主要表现为过量炎性因子、促炎介质的释放及大量炎性细胞的激活,晚期由于免疫细胞的凋亡及炎性介质的耗竭,机体表现为免疫抑制状态[4-5]。单核/巨噬细胞在脓毒症中不仅参与非特异性免疫监视和防御作用,还发挥抗原提呈作用,并产生大量细胞因子激活其他免疫细胞。巨噬细胞主要可极化为经典活化的M1型和选择性活化的M2型2种类型。M1型巨噬细胞主要参与免疫应答,发挥促炎作用,M2型巨噬细胞则以抑制炎症反应、促进组织修复为主[6]。目前,对脓毒症的早期诊断及规范的治疗使患者的急性期存活率有所提高,但远期效果尚不理想,病死率仍居高不下[7]。本研究旨在筛选巨噬细胞极化差异基因,寻找调节脓毒症炎症反应的潜在治疗靶点,提高脓毒症患者的生存率,改善预后。
1 资料与方法
1.1数据来源从综合性基因表达(gene expression omnibus,GEO)数据库选取GSE5099和GSE57614 2套基因芯片数据集,其中GSE5099种属为人,芯片平台为GPL96[HG-U133A] Affymetrix Human Genome U133A Array、GPL97[HG-U133B] Affymetrix Human Genome U133B Array,选取其中的M1型、M2型巨噬细胞样本各3例。选取条件:γ-干扰素(interferon-γ,IFN-γ)及脂多糖(lipopolysaccharide,LPS)作用于巨噬细胞后将其极化为M1型,白细胞介素-4(interleukin-4,IL-4)作用于巨噬细胞后将其极化为M2型。GSE57614种属为人,芯片平台为GPL6480Agilent-014850 Whole Human Genome Microarray 4x44K G4112F (Probe Name version),选取其中M1型、M2型巨噬细胞样本各9例,选取条件同GSE5099基因芯片。
1.2差异表达基因的筛选2个基因芯片的原始数据采用GEO2R在线分析软件标准化处理后进行分析,以P<0.01的条件筛选出相关差异基因,再将2组基因芯片筛选出的差异基因取交集,得到最终的差异基因。
1.3差异基因的功能富集分析DAVID(The Database for Annotation,Visualization and Integrated Discovery,网址:https://david.ncifcrf.gov/)是一款在线基因功能分析软件,能对大规模的基因信息进行处理分析。将筛选出的差异基因提交至该网站,通过进行GO (Gene Ontology)分析及KEGG (Kyoto Encyclopedia of Genes and Genomes)通路分析,对差异基因进行富集统计,找出差异基因主要富集的生物学进程或细胞通路,从而找到可能与巨噬细胞极化相关的生物学功能及通路。差异基因显著性富集的临界值为P<0.05。
1.4蛋白质交互作用网络构建STRING (Search Tool for the Retrieval of Interacting Genes,网址:https://string-db.org)数据库是一个在线分析蛋白质的功能及活动形式,反映蛋白质之间的相互作用,构建蛋白质之间交互作用网络的数据库。通过蛋白质交互网络,不仅可以系统地了解蛋白质的功能及相互作用,还能依据交互网络中蛋白质的模块化属性预测蛋白质的功能及蛋白质之间可能的相互作用。将得到的差异基因提交至STRING数据库,通过在线分析并整合蛋白质的相互作用关系构建出蛋白质交互作用网络。
2 结果
2.1差异基因筛选结果GSE5099共筛选出 3 023 个差异基因,其中在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的基因有974个,在M1型巨噬细胞中表达下调而在M2型巨噬细胞中表达上调的基因有2 049个;GSE57614共筛选出2 944个差异基因,其中在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的基因有1 464个,在M1型巨噬细胞中表达下调而在M2型巨噬细胞中表达上调的基因有1 480个。将2组基因芯片筛选结果取交集后得到1 241个差异基因,其中在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的基因有591个,在M1型巨噬细胞中表达下调而在M2型巨噬细胞中表达上调的基因有650个。
2.2差异基因GO分析结果结果见表1。在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的差异基因主要富集在免疫应答、细胞凋亡、炎症反应的调控等,在M1型巨噬细胞中表达下调而在M2型巨噬细胞中表达上调的差异基因主要富集在能量代谢、氧化磷酸化、细胞呼吸、细胞吞噬等生物进程。
表1差异基因富集最显著的前10位生物学功能
Tab.1Themostsignificanttop10biologicalfunctionsofdifferentialgene
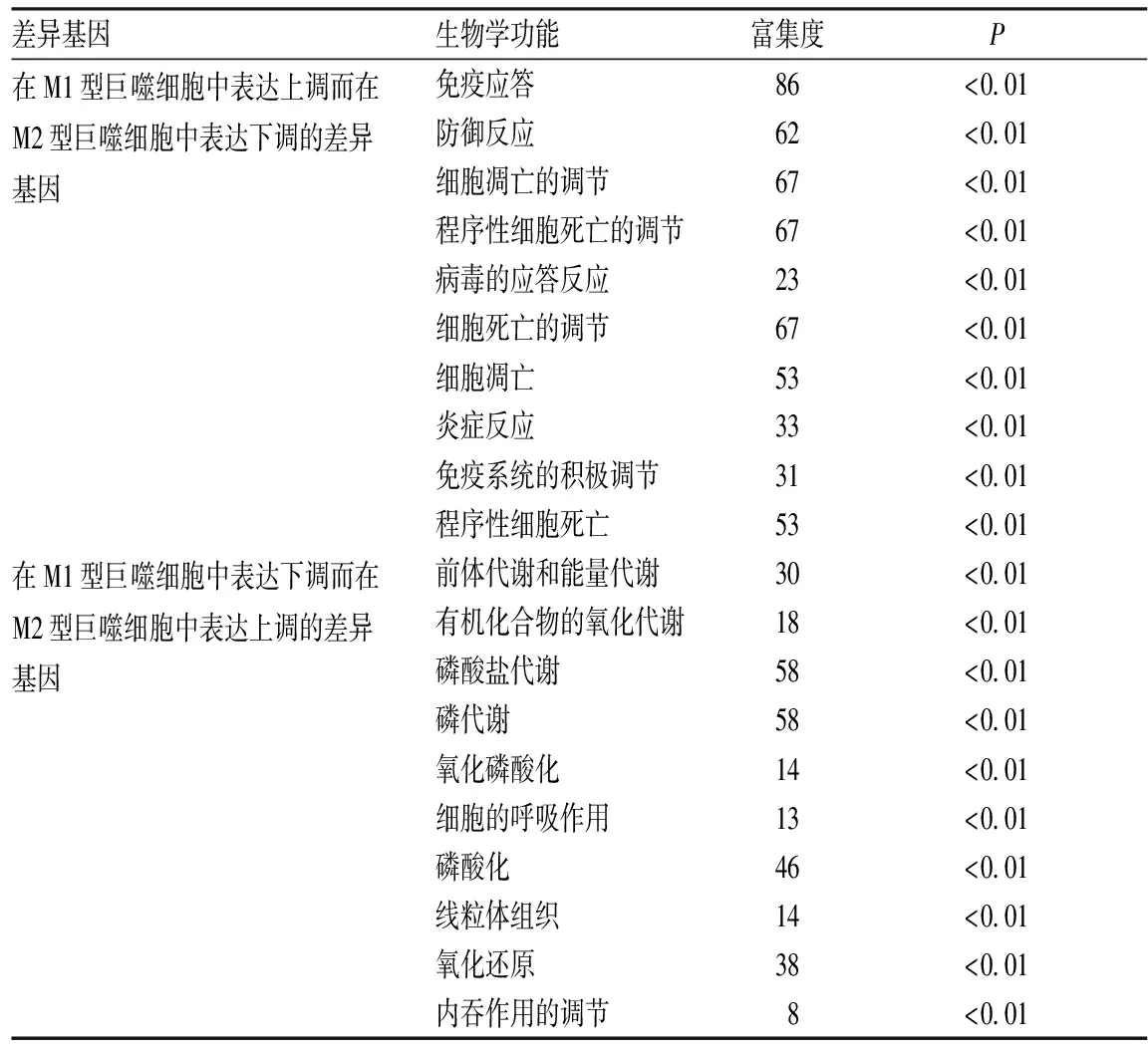
差异基因生物学功能富集度P在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的差异基因 免疫应答86<0.01 防御反应 62<0.01 细胞凋亡的调节67<0.01 程序性细胞死亡的调节67<0.01 病毒的应答反应23<0.01 细胞死亡的调节67<0.01 细胞凋亡53<0.01 炎症反应33<0.01 免疫系统的积极调节31<0.01 程序性细胞死亡53<0.01在M1型巨噬细胞中表达下调而在M2型巨噬细胞中表达上调的差异基因 前体代谢和能量代谢30<0.01 有机化合物的氧化代谢18<0.01 磷酸盐代谢58<0.01 磷代谢58<0.01 氧化磷酸化14<0.01 细胞的呼吸作用13<0.01 磷酸化46<0.01 线粒体组织14<0.01 氧化还原38<0.01 内吞作用的调节8<0.01
2.3KEGG信号通路分析结果结果见表2。KEGG信号通路分析结果显示,在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的差异基因主要涉及多种受体样信号通路、细胞凋亡、趋化因子信号通路等,在M1型巨噬细胞中表达下调而在M2型巨噬细胞中表达上调的差异基因主要涉及氧化磷酸化、促性腺激素释放激素(gonadotropin releasing hormone,GnRH)信号通路,Wnt信号通路等。
表2差异基因富集最显著的前10位信号通路
Tab.2Themostsignificanttop10signalpathwaysofdifferentialgene
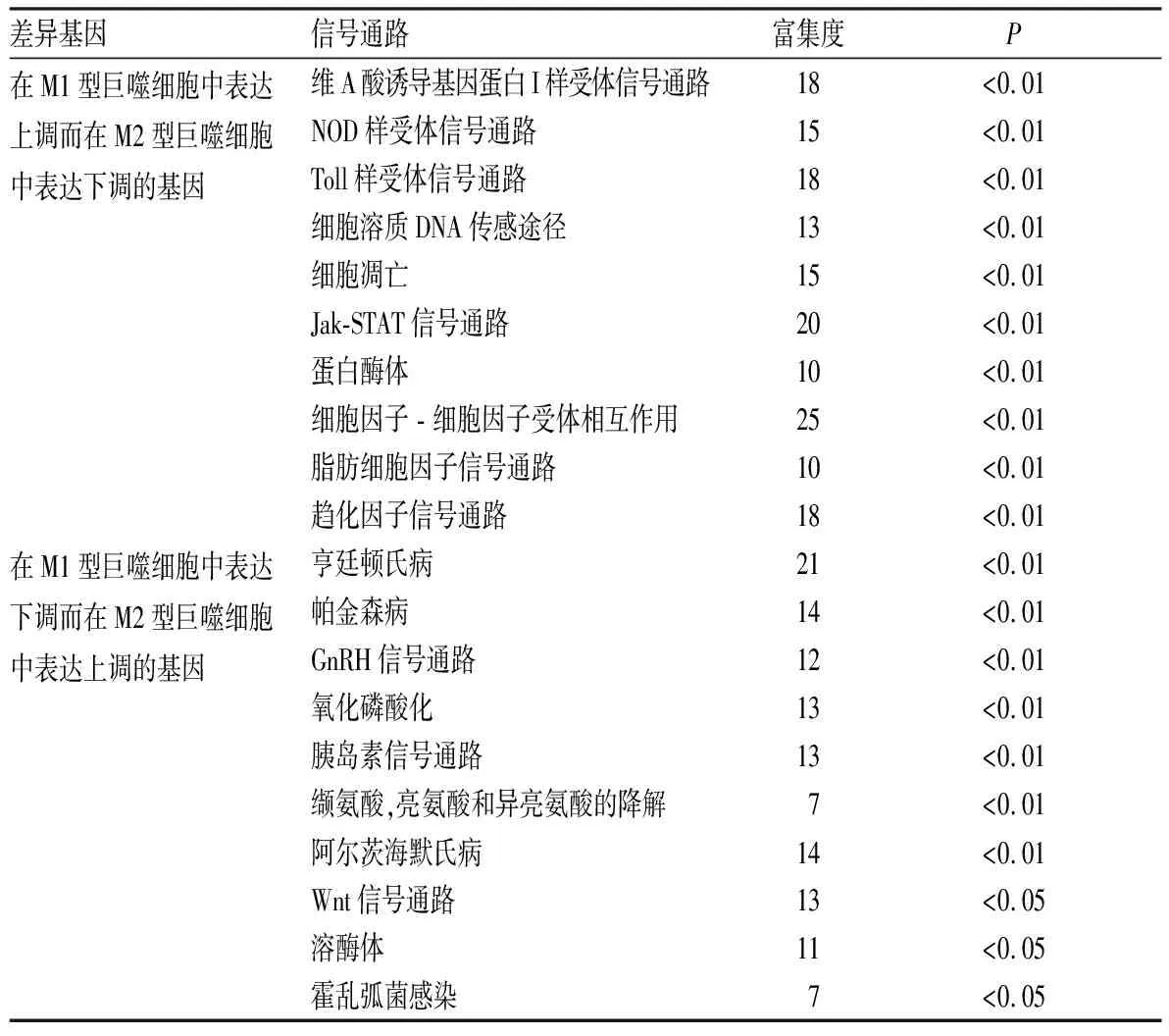
差异基因信号通路富集度P在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的基因维A酸诱导基因蛋白I样受体信号通路18<0.01 NOD样受体信号通路 15<0.01 Toll样受体信号通路18<0.01 细胞溶质DNA传感途径13<0.01 细胞凋亡15<0.01 Jak-STAT信号通路20<0.01 蛋白酶体10<0.01 细胞因子 - 细胞因子受体相互作用25<0.01 脂肪细胞因子信号通路10<0.01 趋化因子信号通路 18<0.01在M1型巨噬细胞中表达下调而在M2型巨噬细胞中表达上调的基因 亨廷顿氏病 21<0.01 帕金森病14<0.01 GnRH信号通路12<0.01 氧化磷酸化13<0.01 胰岛素信号通路 13<0.01 缬氨酸,亮氨酸和异亮氨酸的降解7<0.01 阿尔茨海默氏病14<0.01 Wnt信号通路13<0.05溶酶体11<0.05霍乱弧菌感染7<0.05
2.4蛋白质交互作用分析结果结果见图1。从蛋白质交互作用网络图可以看出,NDUFS3、CHCHD10、ATP5G2、ATP5O、ATP5D、ATP5I、NDUFB8、NDUFA12、NDUFB10、UQCR1、UQCR2等基因编码的蛋白质与多种蛋白存在相互作用关系,在网络图中占据了较为核心的位置。
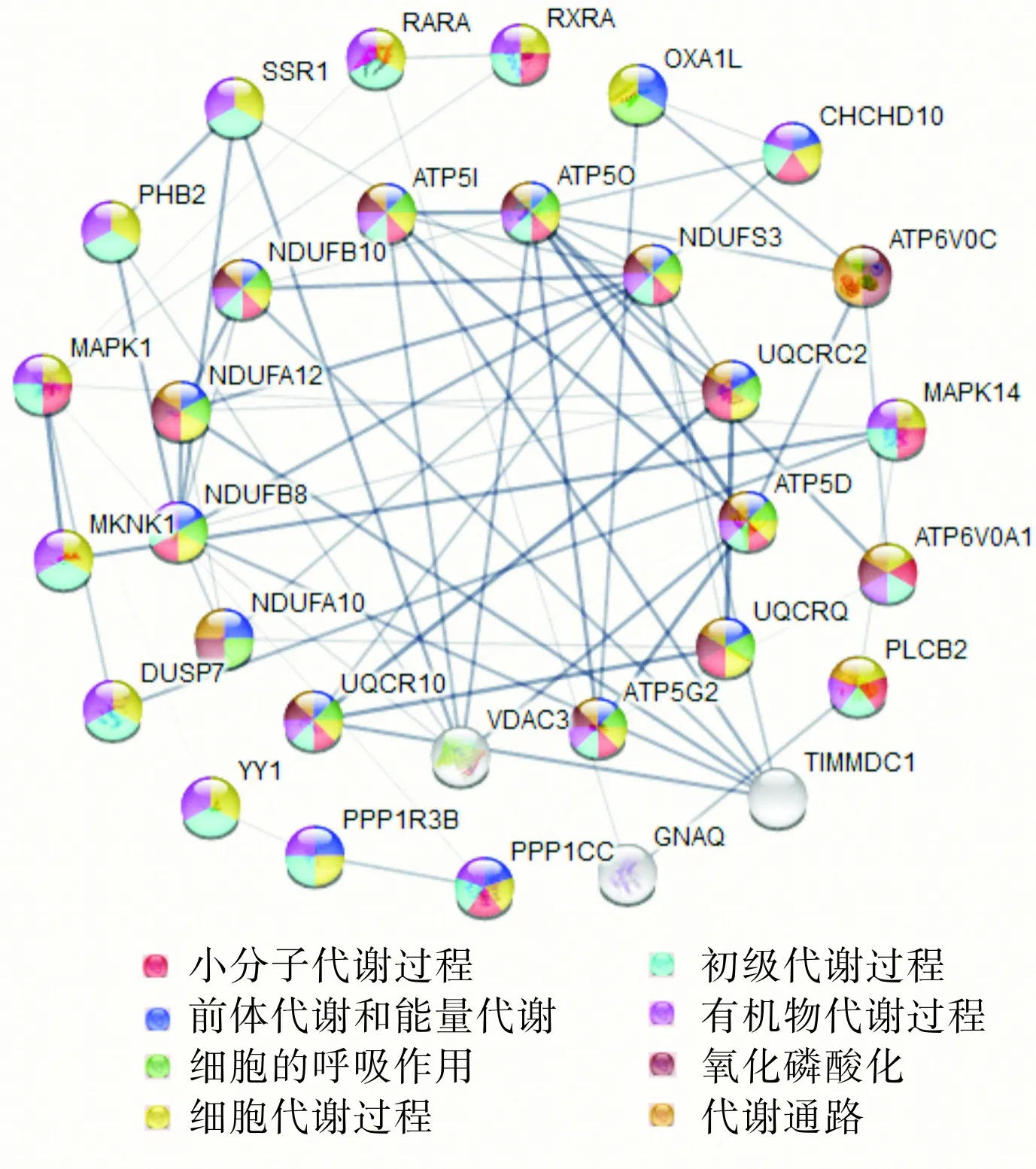
图1差异基因蛋白质交互作用网络图
Fig.1PPInetworkdiagramofdifferentiallyexpressedgenes
3 讨论
目前,脓毒症的治疗方法仍以控制感染、体液支持为主。研究表明,免疫调节的方法在脓毒症治疗中起着举足轻重的作用[8-9]。巨噬细胞是脓毒症炎症反应的重要免疫细胞,其极化为M1、M2型巨噬细胞对炎症反应起调节作用,且M1、M2型巨噬细胞在一定环境下能够相互转化[10-11]。M1型巨噬细胞能促进炎症反应,而M2型巨噬细胞则抑制炎症反应,本研究通过筛选M1、M2型巨噬细胞差异基因,寻找在M2型巨噬细胞中高表达但在M1型巨噬细胞中低表达的基因,以及在M1型巨噬细胞中高表达但在M2型巨噬细胞中低表达的基因,以促进M1、M2型巨噬细胞的相互转化。在脓毒症早期促进M1型巨噬细胞向M2型巨噬细胞转化可适当抑制过度的炎症反应,晚期促进M2型巨噬细胞向M1型巨噬细胞转化可以刺激免疫反应。
本研究共筛选出1 241个差异基因,其中在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的基因有591个,在M1型巨噬细胞中表达下调而在M2型巨噬细胞中表达上调的基因有650个。GO富集分析结果显示,在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的基因主要生物学功能为免疫应答、炎症调节及细胞凋亡等。脓毒症是由一系列级联放大的炎症反应,最终引起微循环障碍和多器官功能衰竭的全身炎症反应综合征,大量的炎性介质、细胞因子的分泌可引起免疫细胞及组织细胞的异常凋亡,细胞凋亡异常又反过来促进脓毒症的发展[12-13]。M1型巨噬细胞中表达下调而在M2型巨噬细胞中表达上调基因则主要富集在能量代谢、氧化磷酸化、细胞吞噬等生物学功能,这些生物学功能与组织的修复存在密切联系。
在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的差异基因的KEGG信号通路分析结果表明,Toll样受体信号通路富集程度较高。有研究表明,Toll样受体(Toll-like receptor,TLR)及其信号通路在脓毒症的免疫反应中起重要作用。作为LPS的主要受体,TLR4与LPS结合后激活下游信号通路进而激活核因子-κB(nuclear factor-κB,NF-κB),从而介导大量炎性介质、细胞因子、趋化因子、黏附分子的产生,导致炎症的持续存在[14-15]。因此,TLR/NF-κB的激活被认为在脓毒症发病中起关键作用[16]。富集显著的几个通路如维A酸诱导基因蛋白I样受体信号通路、核苷酸结合寡聚化结构域样受体信号通路均被报道在脓毒症中起着重要的促炎作用[17-18]。在M1型巨噬细胞中表达上调而在M2型巨噬细胞中表达下调的基因的KEGG信号通路分析结果中,富集显著的信号通路的基因多参与细胞的能量代谢、线粒体呼吸链、磷酸化等生物功能,与GO分析结果一致,因此,这些生物学途径可能与M2型巨噬细胞的组织修复、炎症抑制作用密切相关。
经过STRING数据库的蛋白质交互网络分析发现,一些基因如NDUFS3、ATP5D、ATP5I、NDUFA12、NDUFB8、NDUFB10参与了脓毒症的众多重要的生物学功能途径,在脓毒症的发病机制中起着重要的调控作用。有文献报道,NDUFS3裂解后能破坏线粒体膜电位,干扰烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NADH)氧化和腺嘌呤核苷三磷酸(adenosine triphosphate,ATP)的合成,诱导细胞凋亡[19]。ATP5D、ATP5I等基因则编码ATP合酶的亚基,参与腺苷二磷酸生成ATP[20-21]。基因NDUFA12、NDUFB8、NDUFB10编码的蛋白为线粒体膜呼吸链NADH脱氢酶的亚基,NADH脱氢酶参与线粒体氧化磷酸化,转运电子,形成线粒体内膜质子梯度,为ATP产生提供质子动力[22]。因此,推测上述基因在脓毒症中发挥着重要作用,可能成为脓毒症免疫调节治疗的靶基因。
脓毒症的高病死率使其成为全球死亡的主要原因,本研究采用多种生物信息学方法,通过分析脓毒症中的重要免疫细胞——巨噬细胞极化的基因表达差异,找出核心差异基因,从免疫学层面为脓毒症治疗靶点的选择提供参考,期望能降低患者的病死率。但本研究仅是理论上对基因数据的分析,仍需要大量的实验来加以实践、论证。
