乳酸菌胞外多糖对水稻生长及土壤理化性质的影响
2019-01-24张文平黄诗宸吴佩佳
张文平,王 清,黄诗宸,吴佩佳,程 新
(江西农业大学 生物科学与工程学院,江西 南昌 330045)
近年来,为追求高产高效,农业生产上大量使用农药化肥的生产方式导致土壤生态环境急剧恶化,土壤板结等现象日益严重。将微生物及其代谢产物作用于农作物,对于提高农作物的生长特性及改善农业生态环境有重要意义[1]。现有报道表明,分离自土壤、根际、植物内部等来源的功能性微生物不仅有固氮、解磷、改善土壤结构、拮抗植物病原菌等作用,而且菌体分泌的植物激素、多糖等次级代谢产物还能起到促进植物生长的良好效果[2]。
微生物多糖是一种新型植物调节剂,具有促进作物生长和提高植物抗逆性等特性。Sandhya等[3]发现,接种高产胞外多糖的恶臭假单胞菌(PseudomonasputidaGAP-P45)对干旱胁迫条件下向日葵幼苗的存活率、生物量、根冠比等指标均有显著的提高作用;闫亚南[4]的研究发现,高产胞外多糖的植物促生菌(Rhizobiumsp. W33)对小麦生长有很好的促进作用,且以对根部生长的促进作用尤为明显,同时,土壤中脲酶及蔗糖酶活性也得到了显著提高;Xu等[5]研究发现,蓝藻多糖可以显著提高灌木柠条(CaraganakorshinskiiKom)种子的发芽率及幼苗代谢活性,同时通过清除活性氧、提高抗氧化酶活性等方式减少植物的氧化损伤,具有恢复荒漠地区生态系统的潜在能力;马晓颖等[6]发现,不同来源的微生物多糖均对玉米生长有一定的促生及诱导抗逆性效果,以真菌多糖的效果最好。
综合近年来的研究结果,微生物多糖对作物生长具有一定的促进效果,这一点与葡萄糖等单糖的作用类似,但关于多糖与单糖在促进作物生长的机理上是否存在差异这一点,在本研究检索范围内尚未见报道。本研究以水稻为试验对象,探讨乳酸菌胞外多糖对水稻幼苗生长的影响,并通过土壤农化分析及酶学测定等方式研究微生物多糖与葡萄糖对水稻生长促进作用机理的异同,为微生物多糖应用于农业生产提供理论与实践依据。
1 材料与方法
1.1 材料
1.1.1 试验材料
供试菌株为植物乳杆菌(Lactobacillusplantarum),江西农业大学微生物资源开发与利用实验室保藏。水稻品种为准两优608。供试土壤为红壤,取自江西农业大学生物科学与工程学院实训基地。土壤有机质含量27.31 g·kg-1,碱解氮含量22.81 mg·kg-1,有效磷含量1.63 mg·kg-1,速效钾含量76.88 mg·kg-1,pH 5.80。
1.1.2 培养基的配制
改良乳酸细菌培养基(MRS):蛋白胨10.0 g、牛肉膏10.0 g、酵母膏5.0 g、蔗糖20.0 g、乙酸钠5.0 g、柠檬酸氢二铵2.0 g、K2HPO4·3H2O 2.0 g、MgSO4·7H2O 0.2 g、MnSO4·H2O 0.05 g、吐温-80(聚山梨酯-80) 1 mL,加蒸馏水至1 000 mL,pH 6.8,121 ℃灭菌20 min[7]。
1.1.3 胞外多糖的提取
将活化的植物乳杆菌接种于改良的MRS,37 ℃发酵24 h。参照Rimada等[8]的方法提取胞外多糖。
1.2 试验设计
试验于2017年7月10日在江西农业大学生物科学与工程学院实训基地进行。将不同浓度的多糖溶液施用于装有2 kg过20目筛菜园土的育秧盘(规格:57.5 cm×27.5 cm×2.5 cm)中,混匀,水稻种子经消毒、浸种、催芽后,挑选大小一致的水稻种子播种于育秧盘,每盘200粒,播种方式为旱育秧。铺上一层过0.25 mm筛的细土,定期定量浇水,保持土壤湿润不见明水状态。设置葡萄糖和蒸馏水作为对照。根据前期试验基础,设置多糖浓度为100、1 000 mg·L-1。共设5个处理,分别是施加蒸馏水的空白处理(CK),施加100 mg·L-1胞外多糖的处理(E1),施加1 000 mg·L-1胞外多糖的处理(E2),施加100 mg·L-1葡萄糖的处理(G1),施加1 000 mg·L-1葡萄糖的处理(G2)。每处理设置3次重复。
1.3 测定项目与方法
1.3.1 形态指标
在播种后第15天、第30天时随机取样,测定水稻的形态指标,包括株高、叶长、叶鞘、根长、根体积等形态指标:株高为茎基部至第二长叶叶尖的长度;叶长为第二片叶的长度;叶宽为第三片叶的宽度;叶鞘为包围着茎基部呈鞘状的长度;根长为主根长度。根体积:取20株洗净的秧苗根系完全浸入事先装好3 mL清水的5 mL量筒,上升的刻度数即为20株秧苗根系的总体积。
1.3.2 土壤理化性质
分别于播种后第15、30天取水稻根际土测定土壤理化性质。采用湿筛法测定水稳性土壤团聚体比例[9-10];土壤pH、有机质、碱解氮、有效磷、速效钾等指标参照鲍士旦主编的《土壤农化分析》[11]测定;土壤脲酶活性采用苯酚-次氯酸钠比色法测定;蔗糖酶活性采用3,5-二硝基水杨酸比色法测定;酸性磷酸酶活性采用磷酸苯二钠比色法测定[12];过氧化氢酶活性采用紫外分光光度法测定[13]。
1.4 统计分析
试验结果采用DPS 7.5进行单因素方差分析,对有显著(P<0.05)差异的处理采用Duncan法进行多重比较。采用SIMCA-P 13.0软件进行主成分分析(principle component analysis, PCA)。采用Origin 8.1制图。
2 结果与分析
2.1 对水稻秧苗生物学性状的影响
与空白组(CK)相比,在土壤中添加不同浓度的乳酸菌多糖对水稻幼苗生长有显著(P<0.05)的促进作用(表1)。生长15 d,添加100 mg·L-1乳酸菌多糖的处理组(E1)水稻幼苗的株高、叶长、叶鞘、根长、根体积分别比CK提高了24.77%、15.83%、37.57%、17.76%、31.82%,但其促进效果略逊于同等剂量的葡萄糖处理。当幼苗生长30 d,2个乳酸菌多糖处理(E1、E2)与低浓度葡萄糖处理(G1)对水稻幼苗的促进效果基本相当,而高浓度葡萄糖处理(G2)对水稻幼苗部分指标(株高、叶长等)相较CK反而有一定的抑制效果。
2.2 对土壤结构及理化性质的影响
幼苗生长至15 d时,添加不同浓度的乳酸菌多糖和葡萄糖对土壤pH均无显著影响,但在30 d时,添加乳酸菌多糖可以显著(P<0.05)提高土壤的pH,而葡萄糖并无类似效果。添加不同浓度乳酸菌多糖和葡萄糖后,土壤有机质含量均有提高,特别是30 d时低浓度的糖(E1、G1)效果最为明显。不同浓度的乳酸菌多糖和葡萄糖处理均可以显著(P<0.05)提高土壤中碱解氮含量,1 000 mg·L-1乳酸菌多糖处理还能显著(P<0.05)提高土壤速效钾含量,但各处理对土壤有效磷含量无显著作用。
土壤团聚体是评价土壤肥力的重要指标。由图1可知,添加乳酸菌多糖可以提高水稳性土壤团聚体比例,随着乳酸菌多糖浓度增加,水稳性土壤团聚体的比例亦提高。15 d时,添加1 000 mg·L-1乳酸菌多糖处理(E2)的土壤中水稳性土壤团聚体比例较CK显著(P<0.05)提高了39.74%,但外源添加葡萄糖处理(G1、G2)对水稳性土壤团聚体作用不显著。第30天与第15天相比,添加乳酸菌多糖的处理组土壤团聚体比例有所下降,但与空白组相比仍有显著性差异。
表1不同处理对水稻秧苗生物学性状的影响
Table1Effect of different treatments on biological traits of rice seedling
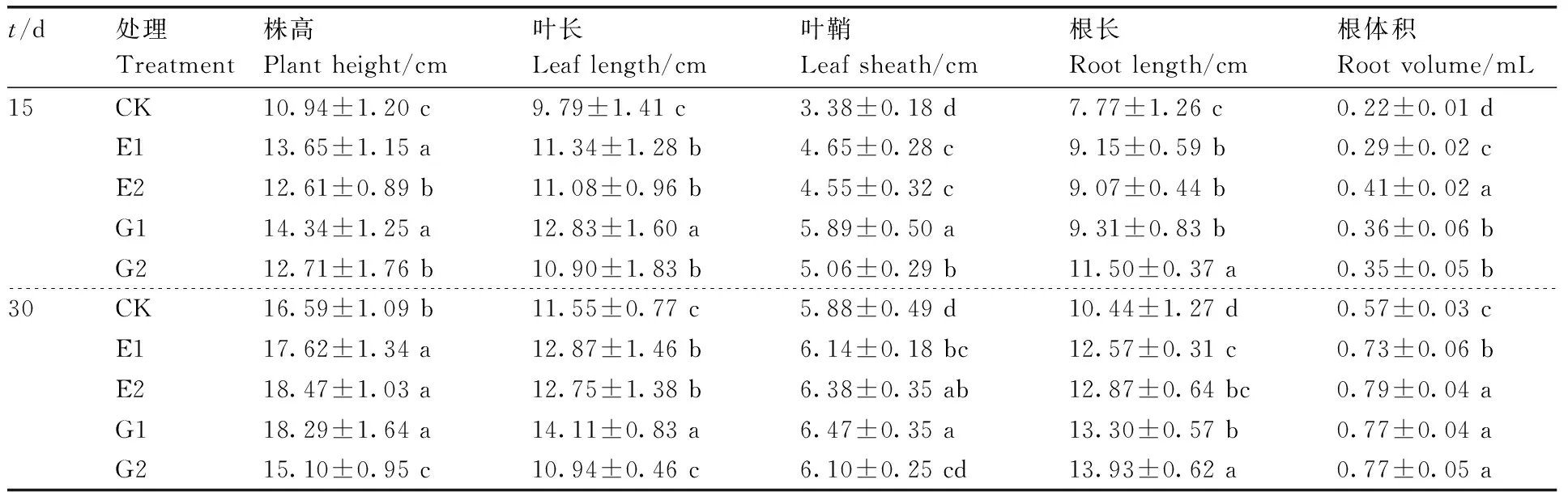
t/d处理Treatment株高Plant height/cm叶长Leaf length/cm叶鞘Leaf sheath/cm根长Root length/cm根体积Root volume/mL15CK10.94±1.20 c9.79±1.41 c3.38±0.18 d7.77±1.26 c0.22±0.01 dE113.65±1.15 a11.34±1.28 b4.65±0.28 c9.15±0.59 b0.29±0.02 cE212.61±0.89 b11.08±0.96 b4.55±0.32 c9.07±0.44 b0.41±0.02 aG114.34±1.25 a12.83±1.60 a5.89±0.50 a9.31±0.83 b0.36±0.06 bG212.71±1.76 b10.90±1.83 b5.06±0.29 b11.50±0.37 a0.35±0.05 b30CK16.59±1.09 b11.55±0.77 c5.88±0.49 d10.44±1.27 d0.57±0.03 cE117.62±1.34 a12.87±1.46 b6.14±0.18 bc12.57±0.31 c0.73±0.06 bE218.47±1.03 a12.75±1.38 b6.38±0.35 ab12.87±0.64 bc0.79±0.04 aG118.29±1.64 a14.11±0.83 a6.47±0.35 a13.30±0.57 b0.77±0.04 aG215.10±0.95 c10.94±0.46 c6.10±0.25 cd13.93±0.62 a0.77±0.05 a
同一处理时间内同列数据后无相同字母的表示差异显著(P<0.05)。下同。
Data marked without the same letters in the same column of the same treatment time indicated significant difference atP<0.05. The same as below.
表2不同处理对土壤理化性质的影响
Table2Effect of different treatments on soil physicochemical properties

t/d处理TreatmentpH有机质Organic matter/(g·kg-1)碱解氮Alkali hydrolysablenitrogen/(mg·kg-1)有效磷Available phosphorus/(mg·kg-1)速效钾Available potassi-um/(mg·kg-1)15CK5.40±0.02 a26.27±4.15 b15.21±0.18 e2.08±0.13 a52.02±2.07 cE16.08±0.33 a33.06±4.15 ab19.01±0.22 d2.00±0.20 a69.57±2.07 bE26.55±1.29 a35.78±4.15 a22.81±0.26 c2.29±0.35 a84.19±6.20 aG15.86±0.40 a30.35±4.15 ab26.61±0.31 a2.68±0.58 a56.41±4.14 cG26.04±1.07 a34.42±4.15 a23.57±0.27 b2.11±0.52 a53.48±4.14 c30CK5.43±0.23 c20.83±4.15 c15.97±0.18 e1.86±0.09 ab37.40±2.07 bE16.64±0.29 a42.57±4.15 a22.81±0.26 c2.35±0.33 a41.78±4.14 bE26.15±0.30 b28.99±4.15 b26.61±0.31 a2.06±0.59 ab69.57±6.20 aG15.34±0.09 c42.57±4.15 a20.53±0.24 d2.11±0.14 ab40.32±2.07 bG25.52±0.11 c35.78±4.15 ab23.57±0.27 b1.54±0.48 b41.78±4.14 b
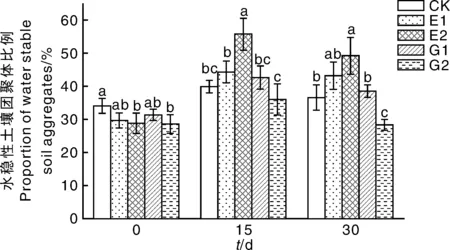
同一处理时间内不同处理间没有相同小写字母的表示差异显著(P<0.05)。下同。The bars without the same lowercase letters in the same treatment time indicated significant difference at P<0.05. The same as below.图1 不同处理对水稳性土壤团聚体的影响Fig.1 Effect of different treatments on proportion of water-stable soil aggregate
2.3 对土壤酶活的影响
添加乳酸菌多糖可以显著(P<0.05)提高土壤脲酶及土壤酸性磷酸酶活性(图2),随着乳酸菌多糖浓度升高,土壤脲酶及酸性磷酸酶活性均呈上升趋势,与CK相比,添加1 000 mg·L-1乳酸菌多糖的处理组(E2),在第15天和第30天土壤脲酶活性分别提高了23.70%和29.66%,土壤酸性磷酸酶活性分别提高了26.17%和28.10%。添加葡萄糖对土壤脲酶的促进效果不及乳酸菌多糖,但对土壤酸性磷酸酶活性的促进效果更明显。
处理15 d,与CK相比,除E1外,其余处理的土壤蔗糖酶与CK无显著差异,但各处理对土壤过氧化氢酶活性均有显著(P<0.05)促进作用。处理30 d时,添加乳酸菌多糖的处理可以显著(P<0.05)提高土壤蔗糖酶及过氧化氢酶活性,随着乳酸菌多糖浓度增加,蔗糖酶及过氧化氢酶活性有所下降。与CK相比,添加100 mg·L-1乳酸菌多糖的处理组(E1)中,土壤蔗糖酶及过氧化氢酶活性分别提高了28.89%、68.19%,添加葡萄糖的处理(E1、E2)对于土壤蔗糖酶活性有一定的促进效果,但对土壤过氧化氢酶活性则表现出抑制作用。
添加乳酸菌多糖和葡萄糖均会对土壤酶活性产生较大影响,但影响效果有异,说明二者的作用机理可能并不相同。
2.4 水稻生理指标与环境因子的相关性分析
由表3可知,水稻株高与叶长、有效磷、脲酶指标存在显著(P<0.05)相关性,相关系数分别为0.89、0.82和0.85;根长与根体积、酸性磷酸酶有极显著(P<0.01)相关性,相关系数分别达到0.93和0.95;根体积与碱解氮、酸性磷酸酶的相关系数分别为0.88、0.86,呈显著(P<0.05)相关性;土壤pH与土壤蔗糖酶、过氧化氢酶显著(P<0.05)相关,土壤有效磷和脲酶、过氧化氢酶均表现出显著(P<0.05)相关性。以上结果说明,土壤理化及酶学指标与水稻植株生长间存在紧密的关联性,需要做更进一步的分析。
2.5 水稻生理指标与环境因子的主成分分析
主成分分析是将多个变量通过线性变换选出较少个数重要变量的一种降维的多元统计分析方法,可以去除不重要的信息,保留所需要的重要信息[14]。对第15天及第30天所测定的试验指标进行主成分分析,结果如表4和表5所示。

图2 不同处理对土壤酶活的影响Fig.2 Effect of different treatments on soil enzymes activities
表3各水稻生理指标及土壤环境因子的相关性
Table3Correlation of physiological indexes of rice and soil environmental factors

指标PLLLLSRLRVWSApHOMAhnAphApoUASAApaCAIndexPL1LL0.89*1LS0.690.791RL-0.050.180.651RV0.250.350.810.93**1WSA0.580.300.47-0.120.151pH0.350.160.080.060.25-0.181OM0.210.550.570.710.63-0.460.281Ahn0.220.120.590.730.88*0.210.540.361Aph0.82*0.780.39-0.140.070.050.660.410.111Apo0.490.150.500.220.520.740.40-0.200.760.161UA0.85*0.630.520.010.340.360.780.230.510.86*0.631SA0.310.280.420.550.67-0.210.87*0.620.800.520.420.681Apa-0.060.200.680.95**0.86*0.04-0.240.590.58-0.290.17-0.150.281CA0.530.320-0.28-0.06-0.100.91*0.150.210.84*0.250.82*0.63-0.531
PL,株高;LL,叶长;LS,叶鞘;RL,根长;RV,根体积;WSA,土壤团聚体;OM,有机质;Ahn,碱解氮;Aph,有效磷;Apo,速效钾;UA,脲酶;SA,蔗糖酶;Apa,酸性磷酸酶;CA,过氧化氢酶。下同。*,**分别表示相关性达显著(P<0.05)或极显著(P<0.01)水平。
PL, Plant height; LL, Leaf length; LS, Leaf sheath; RL, Root length; RV, Root volume; WSA, Water-stable soil aggregate; OM, Organic matter; Ahn, Alkali hydrolysable nitrogen; Aph, Available phosphorus; Apo, Available potassium; UA, Urease activities; SA, Sucrase activities; Apa, Acid phosphatase activities; CA, Catalase activities. The same as below. *, ** represented significant correlation atP<0.05 orP<0.01, respectively.
当水稻生长至15 d时:决定第一主成分(PC1)的主要有株高、叶鞘、根体积、碱解氮等指标,反映了原始数据48.45%的信息;决定第二主成分(PC2)的主要有脲酶、蔗糖酶、有效磷、速效钾等指标,前2个主成分的累计贡献率为78.27%;决定第三主成分(PC3)的主要有水稳性土壤团聚体比例、根长和过氧化氢酶等指标,前3个主成分的累计贡献率为91.67%。当水稻幼苗生长至30 d时:决定第一主成分的主要有脲酶、蔗糖酶、叶鞘等指标,反映了原始数据44.31%的信息;决定第二主成分的主要有酸性磷酸酶、过氧化氢酶、根长,前2个主成分的累计贡献率为70.96%;决定第三主成分的指标为水稳性土壤团聚体比例,前3个主成分的累计贡献率为87.26%。
表4各指标主成分分析
Table4Principal component analysis of each index

项目Item第15天 15th dayPC1PC2PC3第30天 30th dayPC1PC2PC3特征值Eigen value7.274.472.016.654.002.44方差Variance/%48.4529.8213.3944.3126.6616.29累计贡献率Cumulative/%48.4578.2791.6744.3170.9687.26
表5各指标主成分分析旋转后的成分载荷矩阵
Table5Rotated component matrix of principal component analysis of each index

指标Index第15天 15th dayPC1PC2PC3第30天 30th dayPC1PC2PC3PL0.310.130.130.28-0.22-0.28LL0.290.23-0.040.27-0.09-0.16LS0.310.250.110.370.17-0.24RL0.200.090.450.210.410.12RV0.34-0.01-0.170.300.310.00WSA0.16-0.27-0.490.10-0.05-0.60pH0.30-0.24-0.080.25-0.240.31OM0.30-0.210.130.230.180.32Ahn0.320.23-0.040.300.180.03Aph0.190.32-0.370.26-0.320.08Apo0.20-0.35-0.280.24-0.02-0.32UA0.10-0.45-0.050.33-0.26-0.05SA0.23-0.310.270.320.000.38Apa0.290.28-0.130.150.46-0.03CA0.21-0.190.410.20-0.390.22
从图3能够直观地看出,当水稻幼苗生长至15 d时,CK落在第三区间,E1、E2都落在第一区间,G1、G2落在第四区间,与E1、E2差异明显。当培养至30 d时,葡萄糖处理(G1、G2)有明显向左移动的趋势,G2处理组已移至第三区间,而E1、E2仍落在第一区间,与单糖处理的差异更加明显。上述结果说明,葡萄糖和多糖均影响水稻的生长及土壤性质,但其作用方式有较大区别。
3 讨论
添加外源糖类物质已成为促进作物生长、提高作物品质的有效途径[15]。据现有文献报道,多糖和单糖的添加对作物生长均具有较好的促进作用[16-20],但关于多糖与单糖促进作物生长机理的差异研究则鲜见报道。根据现有报道,单糖对水稻的促进效果更多地表现为作为营养物质被植物直接吸收利用或者加速土壤微生物的生长,进而促进植物的快速生长[21],这在某种程度上也解释了本研究中葡萄糖的促进效果作用时间较短,且短期内葡萄糖的促进效果略微好于多糖这一现象。多糖对植物生长促进作用的机理研究目前尚不多见,从本研究结果可以看出,尽管短期内多糖对作物生长的促进效果较葡萄糖略差,但随着生长时间延长,两者之间的差异逐渐缩小,表明乳酸菌多糖的添加可以持续性地促进水稻生长,同时对土壤团聚体、pH等土壤理化性质亦有极大影响,这与艾雪[22]的研究结果一致。添加相同浓度的葡萄糖处理对前述指标作用并不明显,说明两者的作用机理并不相同,后续的PCA分析结果也证明了这一点。
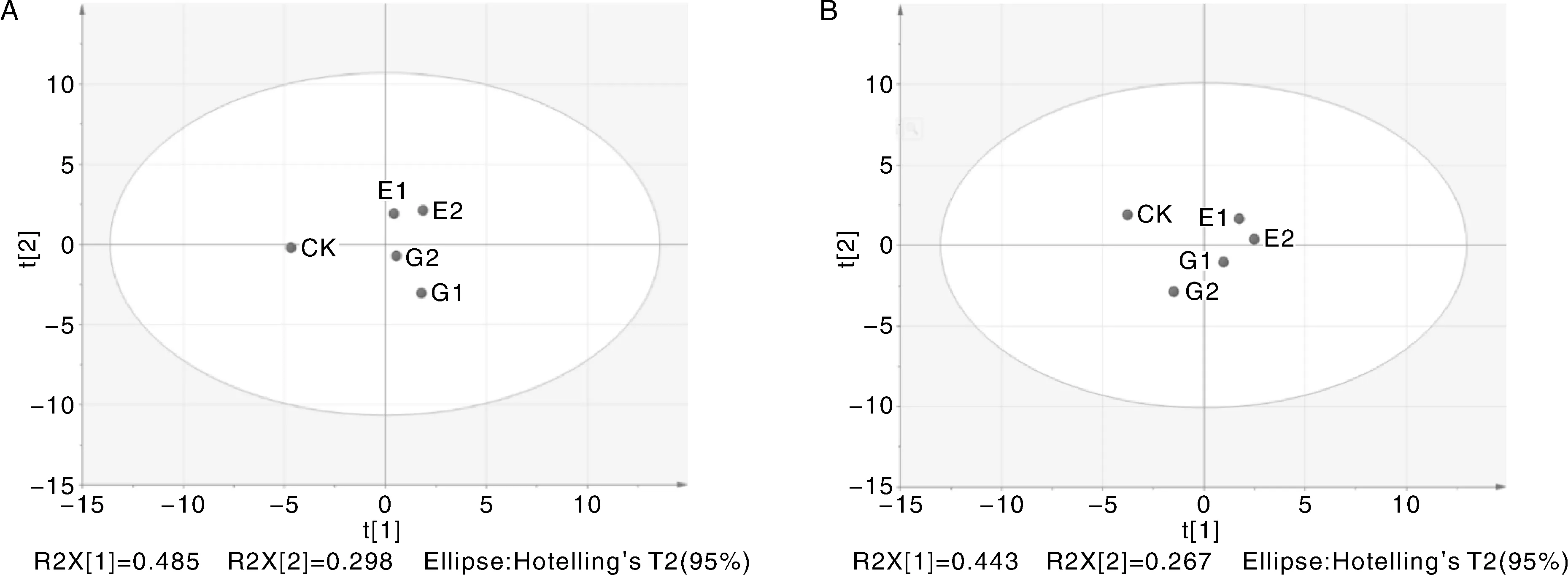
图3 第15天(A)和第30天(B)5个处理的主成分分析得分图Fig.3 Principal component analysis score chart for 5 treatments on 15th day (A) and 30th day
土壤是作物生长的环境基础,对作物生长起着至关重要的作用。水稻是喜微酸的作物,但土壤的过度酸化对其种子萌发及幼苗生长均会产生不利的效果[23-24],这一点在南方红壤地区表现得更加明显。本研究表明,添加乳酸菌多糖可以有效提高土壤pH。随着乳酸菌多糖浓度的增加,其土壤pH有逐步升高的趋势,而且随着时间延长,乳酸菌多糖对提升土壤pH的效果更加明显。然而,添加葡萄糖对土壤pH并无显著影响。这与崔文明[25]的试验结果并不一致,说明不同类型的土壤可能会对试验结果有较大影响。土壤酶活是土壤生物学活性的重要组成部分,也是反映土壤生物化学动向和强度的重要指标[26-27]。本研究发现,添加乳酸菌多糖可以提高土壤脲酶、蔗糖酶、酸性磷酸酶、过氧化氢酶等酶活,与孙姗姗[28]利用3种外源糖处理玉米种子显著提高根际土壤酶活、改善土壤环境、促进作物生长的研究结果类似。然而添加葡萄糖对脲酶、蔗糖酶活性均无显著效果,对过氧化氢酶活性甚至还有一定的抑制作用。综合上述结果,在本试验条件下,微生物多糖对土壤理化性质的改善效果更明显。
综合本研究结果,在土壤中添加一定浓度的乳酸菌胞外多糖可以有效促进水稻幼苗的生长,改善土壤结构,提高土壤酶活。但微生物多糖与葡萄糖对作物生长和土壤特性的影响效果并不一致,说明两者之间的作用机理可能存在较大差异,需要进一步分析。
