竹林鞭根系统研究综述
2019-01-24李平衡周国模周宇峰
童 亮, 李平衡, 周国模, 周宇峰, 李 翀
(1.浙江农林大学 浙江省森林生态系统碳循环与固碳减排重点实验室,浙江 杭州 311300;2.浙江农林大学 省部共建亚热带森林培育国家重点实验室,浙江 杭州311300;3.浙江农林大学 环境与资源学院,浙江 杭州 311300)
竹子为禾本科Gramineae竹亚科Bambusoideae植物,全球有150属1 225种,竹林面积有3 100多万hm2,被称为 “世界第二大森林”。中国地处世界竹子分布的中心,竹子资源十分丰富,竹子广泛分布于福建、江西、浙江、湖南、四川、广东、广西、安徽、湖北、重庆、云南等省。全国现有竹类植物34属534种,约占世界竹种的40%,竹林面积已达601万hm2,约占世界竹林面积的20%,是名符其实的 “世界竹子王国”[1]。竹林鞭根系统是竹林生长发育的基础。以毛竹Phyllostachys edulis为例,它通过鞭根繁殖新竹,且其特有的大小年交替生长特征与地下鞭根结构及其动态生长有着密切的关系,如大年,主要是新笋萌发和新竹的 “爆发式”高生长,新笋萌发决定于鞭根系统的年龄与空间结构,而新笋的数量与质量影响新竹的生长与该年的地上生产力,新竹短时间内完成高生长也是由不同的老竹通过地下相连的鞭根系统给新竹集中输送碳、水和营养物质而造成的[2];小年,新笋萌发和新竹生长非常少,地上立竹光合固碳大部分都是输入地下相连的鞭根系统,用于新鞭根在地下空间的生长发育与蔓延扩展,小年地下鞭根的生长发育由地上立竹提供物质能量,因此很大程度上受到地上立竹年龄和空间结构的影响。因此,竹林鞭根系统对竹林生态系统功能及生产力长期维持尤为重要。对竹林根鞭结构的研究可以追溯到20世纪60年代,经过50余年的研究,已经对竹林鞭根系统结构有了较多的了解,包括根鞭的数量特征、空间分布、年龄结构、生长规律等。经营措施如何影响鞭根系统,鞭根系统的探测等都是竹林鞭根系统研究的重要问题。因此,本研究将从竹林鞭根系统的结构特征、经营对鞭根系统结构的影响以及鞭根系统观测方法等论述国内外竹林地下鞭根研究现状和存在的问题,为今后开展竹林鞭根系统研究提供参考。
1 竹林鞭根系统结构特征
根据地下茎的类型,竹子可以分为合轴丛生、合轴散生、单轴散生、复轴混生等4种类型[3],其中合轴丛生型不同于其他3种,并无真正的地下茎,因此不在本研究的讨论范围。关于竹林地下系统,最早是由熊文愈[4]提出的,他指出毛竹的地下系统包括竹鞭、鞭根、竹蔸和竹根,其中鞭根是指竹鞭上所生长的根,而竹根则是指竹蔸上所生长的根。鞭根系统是由同一母竹发出,各年龄竹鞭和鞭根相互联结的组合[5]。
1.1 竹林鞭根系统的数量特征
鞭根系统的数量指标包括单位面积或体积土壤内竹鞭鞭段数、鞭长和根长、鞭径大小、鞭体积、根和鞭质量、表面积等。根系具有相似的数量特征,即根系在质量和长度上都大于鞭系。如周本智[6]在研究竹林地下系统动态时发现,毛竹林竹鞭总量为6 179.44 kg·hm-2,根系总量为9 071.2 kg·hm-2,分别占鞭根系统的40.52%和59.48%;根系总长约是鞭根系统的1 000倍,表面积也是鞭根系统的20倍。这些都与吴炳生[7]的结果类似,均表明在鞭根系统中根系的数量占主导地位,这也反映了竹林拥有强大的地下吸收能力。
1.2 竹林鞭根系统的年龄结构
毛竹个体寿命约18 a,其有限的生命依靠竹鞭不断抽发新鞭得以延续[8]。鞭系中每年新老鞭的交替,日积月累形成了竹鞭多龄级结构。根据不同年龄鞭段的外部特征差异可以识别竹鞭的年龄[8-9]:如随鞭龄增加,鞭体颜色逐渐加深,表现为从淡黄色到黑色变化,同时鞭段上的鞭根支根数量也会增加并最后衰老死亡。而对竹鞭年龄划分[10]目前尚无统一标准,一般按鞭龄分为幼龄鞭(1~2 a)、壮龄鞭(3~6 a)和老龄鞭(7 a以上)。幼壮龄鞭在竹林鞭根系统中占的比例极大,毛竹林[11]中幼壮龄鞭鞭长、鞭质量分别占 81.3%和 81.8%;雷竹 Phyllostachys violascens林[12]中壮龄鞭占总鞭数的 85%;篌竹 Phyllostachys nidularia林[9]壮龄鞭占竹鞭总量的65%。不同竹种的各鞭龄具有明显的分层分布现象,且均表现为幼壮鞭普遍较老龄鞭分布浅,原因可能是由于上层土壤土质疏松,通气透水性好,且肥料分布于土层表面,而幼壮龄竹鞭具有较强的养分吸收能力[13],加上竹鞭生长的趋肥性,使得幼龄鞭分布在上层。此外,竹鞭抽鞭发笋能力与鞭龄及其生长状况关系密切。如毛竹林中立竹基本长于壮鞭和老鞭上,占到98%, 幼龄鞭鲜有竹株[14]。
1.3 竹林鞭根系统的空间分布
竹林鞭根系统在土壤中的垂直分布具有明显的规律,不同竹种、不同年龄的鞭根系统都表现出分层分布的现象。例如,众多学者对毛竹[7,11,14]地下结构的研究均表明:毛竹作为一种浅根系植物,其竹鞭主要分布在0~60 cm土层,占竹鞭总数的95%以上;雷竹[12,15]竹鞭大量分布于 0~40 cm土层;苦竹Pleioblastus amarus[16]竹鞭集中在0~20 cm土层。与竹鞭类似,竹根分布也呈现出垂直分层特点,其中毛竹[6]竹根集中在20~30 cm土层,毛竹竹根质量、长度、体积、表面积均占40%左右;篌竹[9]根系有77%分布于0~20 cm土层,20%分布于20~40 cm土层。竹鞭生长在水平方向也具有方向性,可分为水平、向上、向下共3个方向[16]。根据经验,竹鞭走向大致与竹秆基密、疏节间圆切方向平行。此外,竹鞭鞭梢具有顶端优势,一定程度上抑制了侧芽萌发,但在竹鞭延伸过程中,鞭梢抵触到硬物或受到机械损伤而折断时顶端优势减弱,会于断点附近萌发岔鞭。常见的分岔类型一般为一侧单岔、一侧多岔、两侧单岔和两侧多岔4种[12]。毛竹岔鞭多集中于断点附近,距离断点9节内产岔鞭数量占到72.7%[5]。
1.4 竹林鞭根系统的生长特性
竹林的鞭根系统生长规律由于竹种的差异会略有不同,但总体生长季节都在3-11月。以毛竹为例,毛竹竹鞭一般以2 a为1个周期,循环交替生长。竹鞭活动开始于春季,结束于冬季,生长期长达6~8个月,生长速度呈现慢—快—慢的规律;通常,当气温上升到8℃时,鞭梢便开始生长,于6-7月达到生长高峰,由于8月毛竹进入了孕笋期,竹鞭生长趋于渐缓并停止生长[17]。大小年竹鞭生长也存在差异,大年竹鞭生长期普遍短于小年,一般为6个月。毛竹的根系生长于3月下旬开始,到11月上旬结束,6-9月为生长高峰期。由于毛竹鞭根是伴随着竹鞭发育产生的,竹鞭和鞭根的年龄差异一般不超过1个月,所以毛竹鞭根的生长规律与竹鞭相近[17]。通常,鞭根着生于离鞭梢先端10~15 cm的节上,随鞭龄增加会产生1~4级支根,鞭根更新依靠伸长或分生出新的支根[8]。鞭根是在竹鞭生长的基础上进行的,与鞭龄关系密切;而鞭根生长的状况又直接关系到竹鞭营养物质的吸收,两者彼此依存,相互促进。
1.5 竹林鞭根系统结构与生物量
竹林地下部分在生态系统物质循环和能量流动中发挥重要作用。就目前研究来看,竹子各器官生物量分配存在差异。李振基等[18]对闽南毛竹林生物量和生产力研究发现:毛竹竹根和竹鞭生物量分别占总生物量的17.25%和10.16%,生物量大小表现为竹秆>竹根>竹枝>竹鞭>竹叶;雷竹地下生物量占16.66%,其中竹根生物量大于竹蔸[19]。许多研究均表明竹林地下部分与地上部分生物量比值约0.047~0.391[20], 吴炳生[7]却指出毛竹鞭、 根总量为 15.68 kg·m-2是地上部分生物量的 2 倍。 周本智[6]发现细根生物量随季节呈单峰变化,7月最高,1月最低,且细根生物量随土层深度增加而减少。
2 经营措施对鞭根系统结构的影响
2.1 施肥对鞭根系统结构的影响
施肥是一种常用的竹林经营措施,不仅可以促进发笋、壮竹并提高竹林产量,同时也对地下鞭根系统产生重要的影响。施肥可以为竹鞭的主鞭提供养分,使得支鞭能正常发育。施肥也改变了竹鞭在土层中的分布情况,毛培坚等[21]通过连续4 a对毛竹林施肥与不施肥的对照试验中发现:施肥后的鞭段数在0~15 cm土层增加了27.44%,而在16~30 cm土层则减少了10.68%,30 cm以上土层则鞭段数变化不明显。说明长期施肥会导致竹林新鞭上浮,其原因可能是施肥导致上层土壤水肥条件好于下层,而竹鞭生长存在趋肥性,因此为了提高产量建议在施肥的同时进行埋鞭,加深竹鞭在土层中的分布;此外,施肥对鞭根生物量的增加也有明显的作用。朱强根等[22]研究发现毛竹细根各参数与施肥方式和取样位置均存在显著差异,3 m沟施(TR3)的细根生物量显著高于2 m(TR2)和1 m(TR1)水平,且毛竹根长、根尖数和根表面积均表现为 TR3(0.36 kg·m-1)处理显著大于 TR1(0.14 kg·m-1)和 TR2(0.26 kg·m-1)处理, 说明随施肥量增加,毛竹可以通过在施肥点上增加细根生物量来提高对养分的利用,这是毛竹细根生长 “趋富特化”的体现。而从细根总生物量来看,毛竹细根生长主要集中在0~1 mm径级,说明这个直径的细根是毛竹吸收水分和养分的主要器官。
2.2 林地覆盖对鞭根系统结构的影响
竹林覆盖是利用有机材料的增温、保温和保湿作用,来促进笋芽分化、提早萌发并提高竹林产量的经营手段。该技术已在雷竹林、毛竹林中得到了大规模推广与应用。其中对毛竹林覆盖可以显著增加鞭笋产量,如在春笋小年11月对毛竹林地覆盖,经过45~60 d后鞭笋产量是不覆盖产量的2~3倍,且达到极显著差异水平[26]。
但林地覆盖也会对鞭根系统结构造成一些负面影响,比如随着覆盖物分解产生的酚酸类物质的累积对竹林鞭根系统生长有抑制作用,而覆盖也改变了土壤微环境,残留的有机覆盖物往往也影响了土壤及鞭根系统结构的稳定[24],并会随着覆盖年限增加最终导致竹林生产力衰退。此外,金爱武等[25]对雷竹林覆盖处理后发现:覆盖经营的雷竹林鞭根系统结构较差。具体表现为覆盖林分比未经覆盖林分鞭段数虽然明显增加,总鞭长和幼壮龄鞭比例却明显下降;覆盖后芽的绝对数量远低于未覆盖的情况,仅为丰产林单位芽数量的53%;覆盖还改变了竹鞭在土层中的分布情况,覆盖雷竹林竹鞭在0~10 cm和11~20 cm土层分布都在25%以上,而未覆盖雷竹林0~10 cm土层竹鞭分布很少,仅占总鞭的7.39%,竹鞭这种分布情况不利于孕笋发竹。而通过对覆盖林地鞭笋的合理采挖从而优化鞭根系统结构,可以有效解决以上的问题,具体措施包括及时挖除细鞭笋、留养和填埋粗鞭笋,做到弱鞭强挖、壮鞭弱挖等[23]。
2.3 其他经营措施对鞭根系统结构的影响
不同经营类型会对鞭根系统的生长造成多方面的影响。垦复作为竹林抚育的主要措施,可以改善土壤理化性质,优化鞭根系统的结构。周文伟[26]发现:毛竹林在垦复后竹鞭鞭段数、节间长度、鞭长、鞭芽数量均明显增加;垦复也改变了鞭系在土层中的分布,使得竹鞭上浮;垦复还改变了鞭系的年龄结构,而通过疏松土壤、挖除老鞭,可以促进新鞭的生长;此外,垦复鞭径大小影响不大但能减少跳鞭的情况。
毛竹纯林由于其林分结构简单,抗逆性和稳定性较差,林分的质量和产量不断下降,而竹阔混交不仅可以丰富林分结构、调节林内温湿度,还拥有复杂的凋落物,生态位利用程度也较高,更适宜毛竹生长,使得林分抗逆性和稳定性得到提升。郑郁善等[27]在杉木Cunninghamia lanceolata-毛竹混交林研究中发现:一定密度的杉木对毛竹地下结构具有促进作用,杉木密度在0~1 350株·hm-2时,毛竹竹鞭长、鞭质量均随密度增大而增大,但当杉木密度为2 100株·hm-2时,竹鞭长比杉木密度在1 350株·hm-2减少36.3%。毛竹地下竹鞭年龄分布也受混交影响,具体表现为在一定杉木密度下,幼壮龄鞭比例随密度增大,幼壮鞭比例均大于毛竹纯林,说明混交可以促进毛竹地下结构的更新,提高林分活力。同时,由于杉木根系占据了土壤表层,原本大量集中在0~20 cm土层的竹鞭鞭长比例减少,20~40 cm及40 cm以上土层竹鞭鞭、长鞭重比例增加,可见杉木能促使毛竹根系向更深土层延伸,这一规律在马尾松Pinus massoniana+苦竹Pleioblastus amarus混交林、檫木Sassafras tsumu+毛竹混交林中均有体现[28-29]。此外,一定密度的杉木会抑制竹鞭生长,促使毛竹鞭中营养物的积累,使得鞭径增大,这是由于竹鞭遇到杉木根系无法穿越造成机械损伤或断裂,从而解除了鞭梢的顶端优势,导致了岔鞭的萌发,增加鞭段数而减小平均鞭长。由于杉木根系所占面积小,部分面积可以被竹鞭利用,形成互补。
合理挖鞭笋也能优化竹林地下根鞭结构,提高竹鞭粗度、节间长、岔鞭数量等生长指标。通过比较挖鞭笋与不挖鞭笋地下竹鞭鞭径变化发现,挖鞭笋后2.5~3.0 cm鞭段数增加而1.9~2.5 cm鞭段数减少;毛达民等[30]也发现挖鞭笋竹鞭整体鞭径增加了0.3 cm,节间长增加0.31 cm,岔鞭数量增加且差异达到显著水平。及时挖鞭笋可以促进萌发岔鞭,通过弱鞭强挖、壮鞭弱挖留下了生理吸收能力强的根鞭系,不仅提高了土壤空间利用率,也增加了鞭笋产量[13]。
3 竹林地下鞭根系统研究方法
3.1 损伤性方法
3.1.1 挖掘法 竹林鞭根系统的研究方法中最常用的就是挖掘法(表1)。挖掘法的原理就是利用田园工具或高压水气等将土壤移走,再对剩下的根系进行绘图、拍照和测量[31]。其优点在于能直观而准确获取信息,如鞭根系统的数量、质量、长度、鞭径大小、分布情况等。有大量的研究借助挖掘法来探究鞭根系统在地下的结构及分布情况[11,12,32]、鞭根系统的生长情况及其影响因素[33-35]、鞭根系统的生态学研究[36-37]等。通常根据研究目的,设置一定大小的样方(如50 cm×50 cm,2 m×2 m)并分层,在外围挖一定深度的沟,然后挖取样方内每一层的土壤,使其中的鞭系和根系均裸露出土表。在不破坏鞭根系统的基础上对其进行测量,并当场记录相关的参数或挖取样方内所有鞭系和根系带回实验室进一步处理[12,38]。但是对竹林而言,挖掘法对鞭根系统只能进行局部挖掘,无法获得整个林分尺度的信息。同时,挖掘具有严重的破坏性,会对竹林生态系统造成扰动,不利于后续的研究;此外,挖掘过程中对细根破坏严重,使得该方法无法对竹林细根进行研究。
3.1.2 土芯法 土芯法也称土钻法,是一种通过人工或机械手段将不同类型的钻头敲入土壤中进行根系取样的方法[39],可以应用于对根系长度、质量、根系形态学以及根系分布的研究,对于细根研究多采取该方法(表1)。细根直径通常小于2 mm,存在巨大的吸收表面积,是吸收养分和水分的主要器官[40]。细根每年周转1次消耗的净初级生产力占全球陆地生态系统的33%左右[41],其重要性不言而喻。杨清培等[38]采用土芯法选取钻头为5 cm的土钻分6个土层对毛竹的细根进行取样;范少辉等[42]使用土芯法来测定毛竹林细根生物量及其周转。土芯法工具简单便携,操作灵活方便,相比挖掘法能大大降低外业工作强度;又由于受地形限制小且对环境破坏程度低,可以进行大面积重复取样,适合空间异质性高的研究对象,但也存在一些问题。比如钻头大小的选取、采样后对细根的后期处理、难以将零碎有机物与细根分离等都会造成精度问题[43]。
3.2 非损伤性方法
3.2.1 微根管法 微根管技术是一种细根根系观测技术(表1),它通过向土壤中插入透明观察管,形成一个小型观察窗,并利用长筒观察镜或者微型数码相机定期在小观察窗内拍摄记录观察外壁根系生长情况[44]。通常典型的微根窗系统包括以下几部分[45]:一个插入土壤的微根窗管、彩色微型摄像机、控制器和计算机。利用微根管技术可以定点、无损、直观、可重复地观测地下的根系,尤其是对于细根的观测,适用于研究细根的产生、分解和周转过程[6,46-47]。周本智[6]通过微根管法研究竹根生长动态,掌握了竹林根系的生长进程、垂直分布、死亡动态。陈红[48]利用微根管技术了解了毛竹根鞭的密度以及直径的变化。微根管技术也有许多局限,如大面积安装设备带来的高昂费用[49]、存在较长的采样间隔期[50]、以及安装过程中会改变土壤和水文而影响根系生长[51]、无法鉴别微根管图像内的活根和死根等[52]。
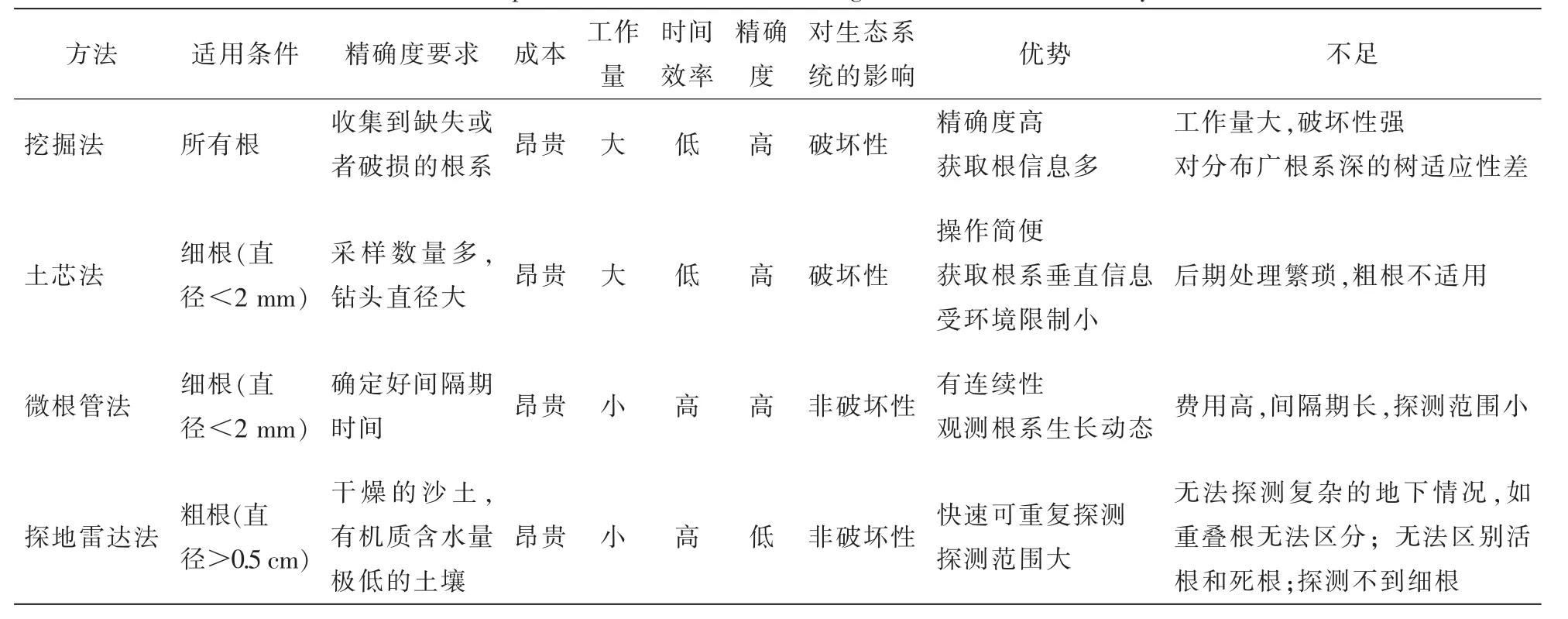
表1 几种竹林鞭根系统观测方法的比较Table 1 Comparison of methods for observing bamboo’s rhizome-root system
3.2.2 探地雷达法 探地雷达法是一种用于定位地下目标的非损伤性地球物理技术[53](表1)。自20世纪90年代起就有学者将其应用于植物粗根根系探测研究,经过10余年的实践与发展,该技术已日臻成熟并在未来根系探测方面具有光明前景。相比较于破坏性的根系研究方法,探地雷达法具有无损、可重复、探测范围广且高效等优点。最初探地雷达法被应用于根系绘图及根系结构的观测。HRUSKA等[54]和ˇCERMÁK等[55]分别使用探地雷达法研究橡树Quercus petraea和槭树Acer campestre的根系分布。随后BARTON等[56]尝试使用探地雷达法估计根系直径,结果表明提取的雷达参数与根系平均直径模型拟合效果良好,R2为0.89,用该模型估计的根径大小的均方根误差为0.56 cm。近年来,随着对根系在全球碳循环中作用的重视,许多学者着眼于使用探地雷达法估算树木的地下生物量[57-66],其中崔喜红等[66]均从雷达中提取了新的参数来估计根径大小并估算根系生物量。也有不少学者对探地雷达法探测的精度问题进行了研究[61-62],表明探地雷达法对根径小于0.5 cm的根系探测效果不理想,并且当根系含水量低于20%时,会直接导致探测失败;此外TANIKAWA等[63]及GUO等[64]就根系生长方向对探测精度的影响等问题展开了研究,并优化了采样方案,提高了探测精度。经过十几年的发展,探地雷达技术目前可应用于树木根系制图、根径大小及根系生物量的估算[61,65]。相对于其他树种根系的分布土层较深、生长方向朝下、粗细不一等复杂情况给探地雷达法探测根系结构带来的困难[66],竹子尤其是毛竹林的鞭根系统均分布在浅层土壤(5~40 cm土层)、竹鞭生长方向多为水平方向、直径相对均一且较大(2~4 cm),且毛竹林为单一树种的林分,地表灌草层相对较少。因此,探地雷达法探测毛竹鞭根空间结构更为容易,能为竹林鞭根系统及其对竹林生产力的影响提供先进的技术支撑。
4 问题与展望
鞭根系统是竹林生长发育的基础,也是竹林生态系统的重要组成部分。随着地下部分在全球碳平衡中的重要性日益凸显,对竹林鞭根系统的研究也随之重视。但就目前来看,其研究在广度和深度上远远滞后于地上部分,在研究掌握鞭根系统的生长和分布规律的基础上,缺乏相关机理以及生态功能(如碳汇)等方面研究。竹子鞭根系统深入地下,观测十分不便,很难进行长期动态调查,传统的挖掘法、土壤剖面法等探测方法不仅费时费力,也无法对鞭根系统进行大范围、长期、重复性的观测和研究[20],甚至会对土壤环境和鞭根系统造成破坏,从而导致研究结果的不确定性。
针对竹林鞭根系统研究中存在的不足以及该领域的发展趋势,未来竹林鞭根系统的研究可以从以下几个方面展开:①在时空尺度下探索竹林鞭根系统生长规律及其与地上立竹的交互关系。传统研究对象停留在某个时间段的竹林鞭根系统,而对不同生长阶段林分尺度的生长特性和物质分配的研究很少,对于鞭根系统在分子、细胞等微观尺度的研究也不多见。竹鞭发鞭、孕笋、成竹决定立竹位置从而影响竹林地上空间结构,而林分结构又能反馈调节地下鞭根系统的生长发育,这些往往在林分尺度上得到体现,其调控机制则需要通过细胞、分子微观尺度才能解释。研究竹林鞭根系统同地上立竹的关系,可以为毛竹这类高效固碳竹种 “爆发式”生长背后的物质和能量分配机制提供理论依据。②研究竹林鞭根系统细根周转及其与根际微生物组成的复杂竹林地下生态系统对全球环境变化的响应机制。诸多研究表明,细根的作用不可忽视。仅直径<2 mm的细根每年生长死亡消耗的净初级生产力占全球陆地生态系统的30%以上[67]。细根通常伴随菌根,拥有更大的吸收表面积,若不考虑细根的生长、死亡及分解过程,土壤中有机质和养分元素周转将被低估20%~80%[40]。因此研究竹林细根的数量和动态,及其对环境的反应能为掌握竹林生态系统结构和功能及竹林经营管理提供科学指导。③竹林鞭根系统研究方法的探索。目前鞭根系统研究滞后的原因在于研究方法落后,伴随计算机等数字成像技术的发展,如探地雷达法、微根管等方法在许多根系研究中得到应用,为竹林地下研究提供了参考。
