张广才岭森林时空变化对步甲群落多样性的影响
2019-01-09孙晓杰杨鑫桑卫国
孙晓杰,杨鑫,桑卫国
中央民族大学生命与环境科学学院,北京 100081
由于人类砍伐森林而造成的生物多样性的丧失已经引起了全球的广泛关注(Zou et al.,2015a)。1990—2015年间,全球森林减少了约1.3×108hm2,有3.1×107hm2的原始森林已经被改造或清理,74%的成熟林被划入次生林中(FAO,2015;Keenan et al.,2015)。为了减缓全球森林消失的速度,大力开展人工造林成为恢复森林面积和控制森林消减的关键措施(陈幸良等,2014)。到2015年,全球人工林面积增加了1.1×108hm2,其中最大的贡献来自中国。近几年中国的植树造林活动使森林覆盖面积每年至少增加3.0×106hm2(FAO,2015)。据报道,在 2010—2015年间,中国的森林覆盖面积净增加1.5×108hm2,是世界增加天然次生林和人造林等永久性森林地区的典范(FAO,2015;陈景和等,2015)。
人工林和次生林的覆盖率逐年增长,除满足人类对木材的需求外,同时在涵养水源、保持水土和防风固沙上也起到重要作用(Sayer et al.,2004),但是目前其对生物多样性保护的价值存在争议,对区域生物多样性和组成的影响也是未知的(Zou et al.,2015a)。一些关注中国次生林和人工林对生物多样性影响的研究表明,人工林对物种保育的作用是消极的(Yu et al.,2002;Yu et al.,2006;Warren-Thomas et al.,2014;Hua et al.,2016),而林冠郁闭度低的人工林相比天然林具较高的林下物种丰富度和多度(Lemenih et al.,2004;Zou et al.,2015a),与此同时,由于毗邻天然林或中度干扰而使得天然林和人工林,次生林和成熟林中物种多样性没有差异(孟庆繁,1996;Zou et al.,2016)。
张广才岭属于长白山支脉,位于中国东北地区东部山地北段的中轴部位(孙红阳等,2015),近年来,该区域大面积成熟林被砍伐,随后建立了落叶松(Larix gmelinii)人工林,同时部分裸地形成次生林。尽管如此,张广才岭地区仍然残余部分成熟森林,此区域的成熟林至少达 400 a,自禁止商业性采伐政策执行后,次生林和人工林林龄也约达50 a。目前,此区域由人工林、次生林和成熟林形成的森林景观为研究不同森林类型中生物多样性模式提供了理想的环境。
在对森林生物多样性的调查中,选取栖息在森林中物种最丰富的类群之一(Zou et al.,2015a)——步甲(Carabidae)进行调查。目前,被描述和定名的步甲超过3.5×104种(Lorenz,2005)。步甲对环境变化高度敏感并且表现出强烈的栖息地特异性,是研究环境变化对生物多样性影响的较好的指示生物(Lövei et al.,1996),常被用来监测北半球温带森林环境的变化(Gao et al.,2009),评估环境质量。此外,它们易于使用简单的陷阱法进行标准化采样,相比于其他昆虫类群更容易被鉴定(Gnetti et al.,2015)。目前为止,多数的研究是针对不同森林类型之间的地表步甲群落生物多样性进行的,而针对地表步甲群落随时间变化的研究较为匮乏(Warren-Thomas et al.,2014;Liu et al.,2015;Zou et al.,2014,2015b,2015a)。步甲具有特殊的生态学特性,在山地景观中,其分布受非地带性环境因子的影响更大,已有研究表明,时间动态变化引起的气候变化对地表甲虫种类的分布具有影响(Judas et al.,2002;Negro et al.,2014;Johansson et al.,2017)。
本研究以张广才岭不同森林类型地表步甲为研究对象,分析其物种多样性、群落结构组成及分布格局,并探究不同林型中物种群落结构时间动态变化,旨在从时间和空间上综合了解不同森林类型对物种的保育作用,选取出最适合的森林类型作为生物避难所,通过采取措施加强保护避难所从而达到保护更多物种的目的。试验选取不同生境类型的森林,在步甲主要活动时间内对其群落展开调查,主要探讨以下科学问题,(1)成熟林、次生林和人工林中步甲物种的丰富度、多度和多样性是否存在明显差异?(2)不同森林类型的步甲群落结构是否相似?(3)随时间动态变化,人工林、次生林和成熟林步甲群落结构发生怎样的变化?变化是否一致?研究结果可为生物避难所的选取和保护工作及生物多样性保护提供科学依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于黑龙江省东南部,牡丹江地区海林市 张 广 才 岭 ( 44°24.2'~44°24.7'N , 128°24.1'~128°24.9'E,图 1),属温带大陆性季风气候。年平均气温为2.8~3 ℃,无霜期平均为110~125 d,初霜为9月上旬,终霜为5月中、下旬,年平均降水量为800 mm,春季降水量少,夏秋季降水多(孙红阳等,2015)。张广才岭地区的天然林面积占森林总面积的91.8%,人工林占总面积的8.2%。该地区作为重要森林保护区,其生态系统得到了很好的保护,针阔混交林覆盖了大部分区域,针叶树种主要有红松(Pinus koraiensis)、云杉(Picea asperata)、冷杉(Abies fabri)和落叶松等。成熟针阔林被砍伐之后形成了以阔叶树种为主的次生林,主要物种有桦树(Betula platyphylla)、杨树(Populus davidiana)及栎树(Quercus wutaishanica)等(刘爽,2014)。

图1 张广才岭森林研究区域样方分布图Fig. 1 Map of the study region in Zhangguangcai Mountain星形符号代表研究区域位置,不同形状符号分别代表20个样方设置的位置Study region marked as black star and 20 sampling plots marked as different black shapes
1.2 样方设置与取样方法
在海拔749~985 m之间选取人工落叶松林、次生白杨林、次生混交林、成熟混交林及成熟针叶林等5种典型的森林类型,其中,落叶松林和白杨林林龄超过50 a,成熟针叶林超过400 a。取样时每种森林类型设置4个独立的20 m×20 m的样方作为重复,每个样方之间至少相隔50 m,以避免边缘效应,保证样方的独立性(Digweed,1995)。使用 GPS记录各样方的地理信息。
利用容积为250 mL,口径为7.5~8.0 cm的塑料杯设置陷阱,将每个样方分成4个10 m×10 m的小样方,分别在小样方的中心设置塑料杯陷阱。塑料杯中盛约占总体积3/4的饱和食盐水,以及适量的洗涤剂用来打破表面张力。塑料杯口与土壤表面保持水平,同时保证土壤表面和杯子之间没有空隙,以确保在自然条件下取样。在陷阱上方离地面6 cm处设置10 cm×10 cm的金属顶以防止水分蒸发和枯枝落叶进入杯中。一般情况下,在山地生态系统中,步甲活动时间多集中于较短的夏季中(Harry et al.,2011),同时,冻土或积雪分布区罕见步甲活动(Gnetti et al.,2015),所以本试验选择2016年5月25日—8月30日开展陷阱试验。为防止水分过多蒸发而无法捕捉步甲或昆虫腐烂,每隔2周收集1次陷阱中的昆虫样本,挑选步甲昆虫用于标本制作并进行鉴定。将每个样方中4个陷阱的数据合并作为该样点的取样数据进行后续分析。本次试验共设置20个样方(图1),80个陷阱,收取5次样本,共获得9073头步甲标本。
1.3 数据处理
物种累积曲线被广泛应用于判断试验设计取样量是否充分以及估计物种丰富度(李巧,2011)。为了验证本次野外试验获取的数据是否具代表性,利用EstimateS 9.1.0对数据进行检验并制图。如果曲线一直上升,表明样本量不足,需增加试验;如果曲线最终变为渐近线,上升平缓,则表明抽样充分,可对数据作进一步分析(李巧,2011;Longino,2000)。
步甲物种周转率采用基于和弦标准化的预期物种共享(Chord-normalized expected species shared,CNESS)相异指数的非度量多维标度法(non-metric multi-dimensional scaling,NMDS)进行分析。CNESS相异指数能够计算具有不同样方大小的样点间基于概率的相似性(刘云慧等,2011)。CNESS相异矩阵计算过程中取样的大小(参数m)可以变化,但必须大于最小取样值且小于最大取样值。m值取值较大时,强调样方间包含的稀有物种的差异,反之则强调优势种。CNESS相异矩阵由 R软件计算得出。通过计算所得的CNESS相异矩阵数据以NMDS形式进行绘图,应力值(stress)是衡量NMDS的拟合效果及方法可行性的指标,当 stress的值≤20%时,模型的拟合结果是可以接受的,可用于数据的解释;在5%~10%时,拟合效果良好;≤5%时,拟合效果最优。运用 STATISTICA 6.0软件计算 NMDS(StatSoft,2001)。
步甲多样性特征主要采用 Shannon-Wiener多样性指数H'、Simpson优势度指数C和Pieluo均匀度指数J来表征(马克平等,1994),并采用Simpson指数量化不同月份之间步甲 α多样性的变化(Barlow et al.,2007)。所有指数均运用R语言(R Development Core Team,2011)中的“vegan”包(Oksanen et al.,2012)进行计算。
Shannon-Wiener多样性指数H':

Simpson优势度指数C:

Pieluo均匀度指数J:

式中,S代表步甲总类群数;Pi代表第 i类步甲的个体数与步甲总个体数的比;ni为第 i类群个体数;N为步甲总个体数。
2 结果与分析
2.1 物种概况
2.1.1 物种累计曲线
基于物种丰富度估计的结果表明,研究区内物种丰度估计量为48种,在5种栖息地获取的步甲总数为42种,抽样完整度为88.4%。依据样地分布的步甲数据绘制物种累积曲线,由图2可知,曲线先急剧上升再逐渐平缓,最后形成一条渐近线,表明本次试验取样充分,样本数据对实际整体数据的代表性强,可进行下一步分析。
2.1.2 不同生境下步甲组成情况

图2 取样区域步甲累积曲线Fig. 2 Species accumulation curves of all samples
试验期间,共采集到步甲9073头,隶属18属42种。多度最高的 5种步甲同属于通缘步甲属(Pterostichus),占总个体数的71.9%。同时,个体数10头以下的属有22个。不同森林类型间优势种差异明显。在落叶松林中,通缘步甲未定种1(Pt.sp.1)和黯通缘步(Pt. adstrictus)占有绝对的优势,分别占落叶松林步甲总个体的38.0%和21.2%;白杨林中通缘步甲未定种1(Pt. sp.1)为优势种,占白杨林步甲种总数的22.0%;成熟混交林的优势种为通缘步甲未定种2(Pt. sp.2),占成熟混交林步甲总数的25.4%;成熟针叶林和次生混交林的优势种相同,同为通缘步甲未定种2(Pt. sp.2)和断通缘步甲(Pt. interruptus),分别占成熟针叶林步甲总数的 25.5%和 21.9%,占次生混交林步甲数量的20.0%和25.1%。
由图 3可知,白杨林步甲物种丰富度最高(n=33),落叶松、成熟混交林、成熟针叶林和次生混交林步甲的丰富度相近(n=25,26,27,27)。各森林类型的物种多度差异较大,相比其他3种森林类型,白杨林和成熟混交林的物种多度(n=1977,2014)明显更高,而成熟针叶林和次生混交林步甲多度(n=1885,1860)次之,落叶松林最低(n=1337)。
2.1.3 不同生境下步甲物种多样性
通过比较5种森林类型步甲群落的多样性发现(表1),人工落叶松林中步甲物种多样性指数H'、优势度指数C和均匀度指数J均为最低,说明人工落叶松林步甲丰富度最低。天然林中不同林型步甲物种H'指数和C指数表现为次生白杨林>次生混交林>成熟混交林>成熟针叶林;J指数表现为次生混交林>成熟混交林>次生白杨林>成熟针叶林,说明成熟针叶林步甲物种多样性较其他天然林低。
2.2 不同森林类型步甲群落结构的关系
对不同森林类型中步甲物种周转率进行分析,结果显示,无论依据步甲群落优势种(m=1)还是稀有种(m=14)进行分析,均发现群落组成格局明显分化成两个区域(图4)。人工落叶松和次生白杨林(第2和3个样方)相互聚集,相比之下,白杨林(第1和4个样方)、次生混交林和成熟针叶林相互聚集,形成一个集群,表明人工落叶松和白杨林(第2和3个样方)的步甲群落结构相似,而次生白杨林(第1和4个样地)、次生混交林和成熟针叶林步甲群落结构相似。
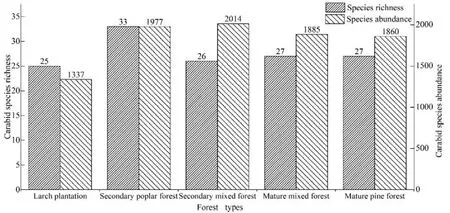
图3 不同森林类型中步甲物种丰富度和多度Fig. 3 Carabid species richness and abundance indifferent forest types

表1 不同森林类型步甲多样性Table 1 Carabid diversity in different forest types
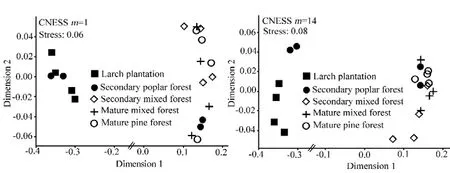
图4 不同生境步甲基于CNESS相异指数的NMDS分析Fig. 4 Non-metric multi-dimensional scaling of carabid samples based on the CNESS dissimilarity matrix for a minimum sample sizes
2.3 步甲群落随时间和生境的变化
随着时间变化,天然林中步甲多度变化趋势基本相似,均为随时间变化呈现先上升后下降的趋势,而人工林中步甲多度呈现缓慢下降的趋势,同一时间内人工林步甲多度明显低于天然林。各林型中步甲多度的变化同时受到时间和生境变化的显著影响(S<2.0×10-16,H=2.6×10-5)(图 5A)。在丰富度上,次生林和成熟林中步甲物种呈现先下降后上升再下降的变化趋势,峰值出现在7月初;次生混交林呈现先上升后下降的变化趋势,峰值出现在8月初;人工林则呈现一直下降的趋势,在相同时间内,人工林步甲物种丰富度均明显低于天然林。时间和生境变化都对步甲物种丰富度产生显著影响(S=2.8×10-9,H=2.1×10-4),两者之间存在极显著交互作用(I=3.4×10-3)(图 5B)。应用 Simpson指数比较不同时间和林型中步甲物种多样性的差异:在天然林中,步甲多样性均随时间变化而缓慢下降,在8月初时达到最低后又逐渐上升;人工林步甲多样性的变化趋势明显与天然林不同,其先逐渐上升,在7月初达到峰值后逐渐下降,除在8月初外,其余时间内其步甲物种多样性均低于天然林。步甲物种多样性变化受到时间和生境变化的显著影响(S=1.7×10-15,H=1.0×10-12),且两者相互作用显著(I=1.1×10-2),即时间变化对物种多样性的影响效应依生境变化而变化(图5C)。
3 讨论
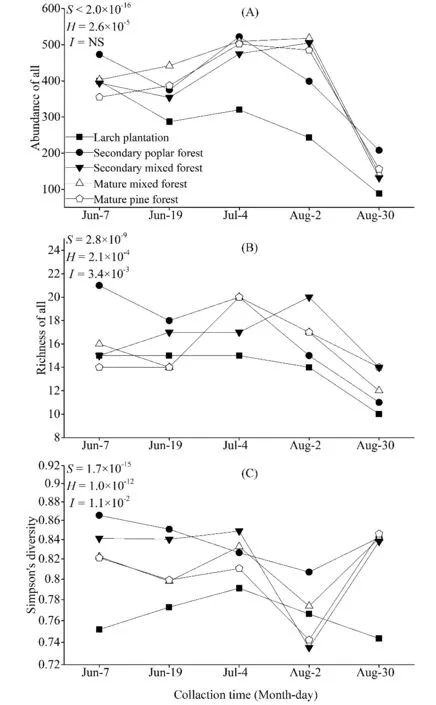
图5 不同森林类型步甲群落多度、丰富度和多样性随时间动态变化趋势Fig. 5 Comparisons variation of carabids richness, abundance and diversity dependent upon temporal variationS:不同季节重复测量双因素方差P值;H:不同森林类型重复测量双因素方差的P值;I:季节和生境之间的相关性值;NS:无相关性P-values for two-way repeated measure ANOVAS are shown for season (S), habitat (H) and the interaction between them (I); values represent mean and standard deviation for each forest type; NS, not significant
在本研究区域内,成熟林、次生林和人工林中步甲物种的多度、丰富度和多样性存在显著区别。人工落叶松林中步甲群落多度、丰富度和多样性均为最低,这一试验结果与其他研究学者结论相一致(Warren-Thomas et al.,2014;Oxbrough et al.,2016)。人工林一般不是本土原有树种,林型结构与本土森林差异较大,其生境也较单一,因而不适应本土物种的生存。本研究区域内的人工落叶松林林龄在50 a以上,郁闭度很高,喜阳性的步甲因不适应阴暗环境而不生存于此林型中(Butterfield et al.,1995;Jukes et al.,2001;Taboada et al.,2008)。与此同时,单一树种构成的森林生态环境的生物量相对较低,由于捕食性类群步甲不能获得足够的食物,而故其在人工林中的分布量较少。
很多研究表明,成熟林中的动植物物种多度与丰富度明显高于其他森林类型(Barlow et al.,2007;Gardner et al.,2007;Zou et al.,2015a),但在本试验研究中,次生白杨林步甲多度、丰富度及物种多样性均明显高于成熟林。这一现象可以通过中度干扰理论(Connell,1978)进行解释,即中等程度的干扰水平有利于物种共存,使物种维持较高的多样性。在本研究区域内,人们为获取经济效益在白杨林里种植一些红松和云杉等针叶树种幼苗,为了提高幼苗存活率而定期砍伐白杨林中的灌木物种,从而导致白杨林地郁闭度降低。郁闭度较低有利于喜温、喜阳的步甲物种的生存。同时,次生白杨林中的白桦和山杨树生长在排水良好、土壤肥沃的地区,为喜湿的步甲提供了优越环境,保持了较高的步甲物种多样性。
步甲物种周转率反映了森林类型中步甲群落结构的相似性和相异性,同时也反映步甲生存条件的相似性和相异性(Zou et al.,2015a)。成熟林中多样的异质性生境与人工落叶松均匀且单一的栖息地条件形成对比,具有多样微生境的成熟森林孕育了更丰富的步甲。本试验中,次生白杨林、次生混交林、成熟混交林和成熟针叶林中多样的微生境类型有利于庇护更多的步甲群落物种,NMDS分析结果显示,成熟林和次生林步甲群落相互聚集,结构相似。人工落叶松林营造的非本土生境中孕育的步甲多属于狭域分布类型,使得其步甲群落结构与成熟林差异较大,突出显现本土步甲物种对森林栖息地环境条件差异的敏感性及对成熟林生境的依赖性。
极端天气条件下的生物栖息地,如山地生态系统,由于冬季气温较低,只有少数无脊椎动物物种活动(Vanin et al.,2007)。一般而言,在山地生态系统环境下,步甲活动仅限于较短的夏季(Harry et al.,2011)。本研究区域位于张广才岭,属于山地生态系统环境,5种森林类型的步甲群落在初夏表现出季节性活动,在6月初,相比于其他4种森林类型,次生山杨林具有最高的步甲物种丰富度、多度和物种多样性,主要是由于大多数步甲物种喜欢在开阔的土地冬眠(Gnetti et al.,2015),次生白杨林乔木和灌木层结构简单,秋季枝叶凋落后,开阔的森林环境可以为步甲提供适宜的冬眠地点(Lövei et al.,1996);7月上旬其他森林类型的步甲多样性明显增多,归因于森林内产生的大量动植物资源为步甲提供了充足的食物,同时次生山杨林步甲由于生存空间或食物竞争导致其向周围栖息地迁移而使得多样性降低。在8月初,物种多度和丰富度急剧减少,主要是由于环境温度较低,步甲活动减少,准备越冬,但同时物种多样性呈上升状态。某些步甲物种越冬时间较早,如本试验中包含种数最多的大步甲属(Carabus)和通缘步甲属(Pterostichus)这类优势属(Gnetti et al.,2015),这将为耐寒和竞争能力弱的步甲种类的生存提供条件。天然林因其微生境多样,资源丰富,为这些类型步甲提供食物和空间资源,而使得天然林中步甲群落物种多样性在8月底呈上升状态。相反,人工林种植单一种类的非本地物种,其营造的特殊生态环境,如较高的森林郁闭度和地表腐殖质较低的腐败速率等,均会导致人工林孕育较少的物种(Warren-Thomas et al.,2014)。在研究步甲群落结构随时间动态变化过程中,发现人工林步甲群落结构在各阶段变化趋势均明显与天然林不同,相同时间内其物种多度、丰富度及多样性均较天然林低,充分说明在构建群落生态过程中人工林物种群落结构与天然林仍存在较大差异,进而推断出随时间动态变化引起的演替对人工林保育物种的作用仍与天然林存在较大差距。
在保护森林中生物物种方面,本研究结果表明,人工林种植应该选取本地树种,且以在成熟林中占主导地位的树种为最好选择,同时,由早期先锋物种构成的桦树或杨树次生林也可以通过人为中度砍伐其林下密被的灌木来提高林下生物多样性(Yu et al.,2006,2008;Zou et al.,2014)。总体而言,本研究认为目前大规模种植单一的非本地物种的方法对提高森林生物多样性是不利的,与此同时,物种多样性的减少也会导致森林丧失控制害虫爆发等生态系统服务功能。改善因森林面积扩张而引起的森林生物多样性降低最有效方法是任林地自然发展,减少人工林的种植,倡导裸地自然演替。同时可以通过在次生林区有针对性地种植当地晚期演替树种来加速演替进程,增加区域成熟林面积,进而提高区域生物多样性。
4 结论
本研究应用传统生态学多样性指数(H'、C和J)对张广才岭不同森林类型步甲物种多样性进行分析,得出天然林中步甲物种多样性较人工林高的结论。利用CNESS相异指数的NMDS分析方法,结合步甲多度、丰富度和多样性(Simpson指数)分析步甲空间分布和时间动态变化,结果表明,成熟林和次生林步甲群落周转率均较人工林高;天然林中步甲群落结构随时间动态变化趋于一致,明显不同于人工林,相同时间内天然林步甲物种多度、丰富度和多样性明显高于人工林,说明天然林步甲比人工林步甲对环境的适应性强,天然林能够保育更多的生物物种。基于以上研究结果,本研究认为天然林对于保育物种发挥的作用是无可替代的,可以通过种植乡土树种,裸地自然生长或在次生林种植当地晚期演替树种以加速森林演替,增大成熟林面积等措施,提高区域生物多样性。
