黑藻叶片PSⅡ叶绿素荧光参数对Cd、Zn单一及复合胁迫的光响应
2019-01-09高桂青吕顺华游济康
高桂青, 吕顺华, 卢 龙, 李 威, 计 勇, 游济康
(1.南昌大学资源环境与化工学院,江西南昌 330031; 2.南昌工程学院土木与建筑工程学院,江西南昌 330099)
重金属作为不可降解的污染物,对生物的危害日益受到全社会的关注[1]。镉(Cd)是植物非必需和毒性最强的重金属元素之一,镉胁迫可诱发PSⅡ捕光复合体改变,从而抑制PSⅡ的捕光色素吸收光能、能量传递和能态激发效率[2];另外,Cd胁迫可通过降低植物叶片叶绿素含量[3]、降低PSⅡ的活性、抑制环式和非环式光合磷酸化[4]等途径,抑制植物对光能的吸收与电子传递、CO2固定等光合作用各过程。锌(Zn)参与植物体光合作用、蛋白质与核酸代谢,过量的锌又会对植物细胞结构造成毒害[5]。由于重金属元素之间的加和、协同、拮抗等作用使水体重金属污染的评价和监测更加复杂,如何正确评价重金属元素之间的联合作用对水环境治理具有重要的意义[6]。
黑藻是我国湖泊中广泛存在的沉水植物,对重金属具有较强的吸收积累能力[7]。目前,国内外重金属胁迫黑藻的研究主要集中在重金属生理生化的伤害反应[8-9]、细胞内部亚显微结构变化[10]等方面,而采用叶绿素荧光动力学技术研究镉和锌复合污染对黑藻胁迫机制的影响甚少。叶绿素荧光参数在探测逆境对光合作用的影响等方面具有快速、简便、灵敏、可靠等特性,能够反映光合系统“内在性”特点[11],被看作研究植物光合作用与环境胁迫程度关系的有效探针[12]。
本试验以黑藻为研究对象,分析了Cd、Zn单一及复合胁迫下黑藻叶片PSⅡ的最小荧光(Fo)、最大荧光(Fm)、最大光化学效率(Fv/Fm)、潜在光化学效率(Fv/Fo)、有效量子产量(YⅡ)、调节性能量耗散的量子产量(YNPQ)、非调节性能量耗散的量子产量(YNO)、光化学荧光淬灭系数(qP)、非光化学淬灭系数(qN)等参数的变化,结合前人研究探讨黑藻受胁迫后的光合生理状态,旨在为阐明黑藻光合系统受Cd、Zn胁迫时光系统响应机制提供参考,为修复湖泊生态系统提供科学依据。
1 材料与方法
1.1 材料培养与处理
黑藻取自赣抚平原灌区六干渠,于2017年6月初选取长势一致(长度为18 cm左右)的植物作为试验材料,将试验植株先种在装有10%Hoaglands营养的塑料箱中驯化7 d,再选取长势一致的黑藻20株移栽到圆形白色塑料桶(高度 47.5 cm、直径22.5 cm)中,桶中底泥厚5 cm,种植后注入自来水,水深40 cm。整个试验在南昌工程学院生态园内完成,塑料桶置于南昌工程学院生态园内,环境温度范围为25~29 ℃。试验分为13组,种植20 d后,除对照组外,其余12组均加入不同浓度的CdCl2溶液和ZnSO4溶液进行处理(浓度设计见表1)。重金属的取值范围结合了预试验结果,以对植株有一定胁迫效应为宜。
1.2 测定方法
1.2.1 叶绿素荧光参数的测定 Cd、Zn单一及复合胁迫黑藻5 d后,采用德国WALZ公司的水下饱和脉冲调制叶绿素荧光仪(DIVING-PAM)测定叶绿素荧光参数,测定于太阳光尚未照射到水面之前进行。测量前,叶片经暗适应20 min后将叶夹原位夹于黑藻体成熟叶片距离叶尖约2 cm处,开启检测光[0.15μmol/(m2·s)],得到暗适应条件下的最小荧光Fo,再由饱和脉冲光[4 000 μmol/(m2·s)]测定最大荧光Fm。

表1 Cd、Zn单一及其复合胁迫的浓度设计
1.2.2 快速光响应曲线 测定于09:30—11:30进行,光合有效辐射(PAR)梯度分别为0、93、249、372、490、555、799、999、1 222 μmol/(m2·s),每个强度的光化光照射10 s,测定最大光化学效率(Fv/Fm)、潜在光化学效率(Fv/Fo)、有效量子产量(YⅡ)、调节性能量耗散的量子产额(YNPQ)、非调节性能量耗散的量子产额(YNO)、光化学荧光淬灭系数(qP)、非光化学淬灭系数(qN)、相对电子传递速率(ETR)等参数。
1.3 数据分析
数据结果为“平均值±标准差”,使用Excel 2016进行绘图。用SPSS 19.0对数据进行单因素方差分析,并采用SNK进行多重比较分析,P<0.05表示差异显著。
2 结果与分析
2.1 Cd、Zn单一及复合胁迫对黑藻Fo、Fm、Fv/Fm与Fv/Fo的影响
由图1可以看出,充分暗适应下各处理的Fo、Fm、Fv/Fm均随着各重金属处理组浓度的增大而降低。这表明随着Cd、Zn处理组浓度的增加,黑藻的PSⅡ反应中心受到的损害程度增大。
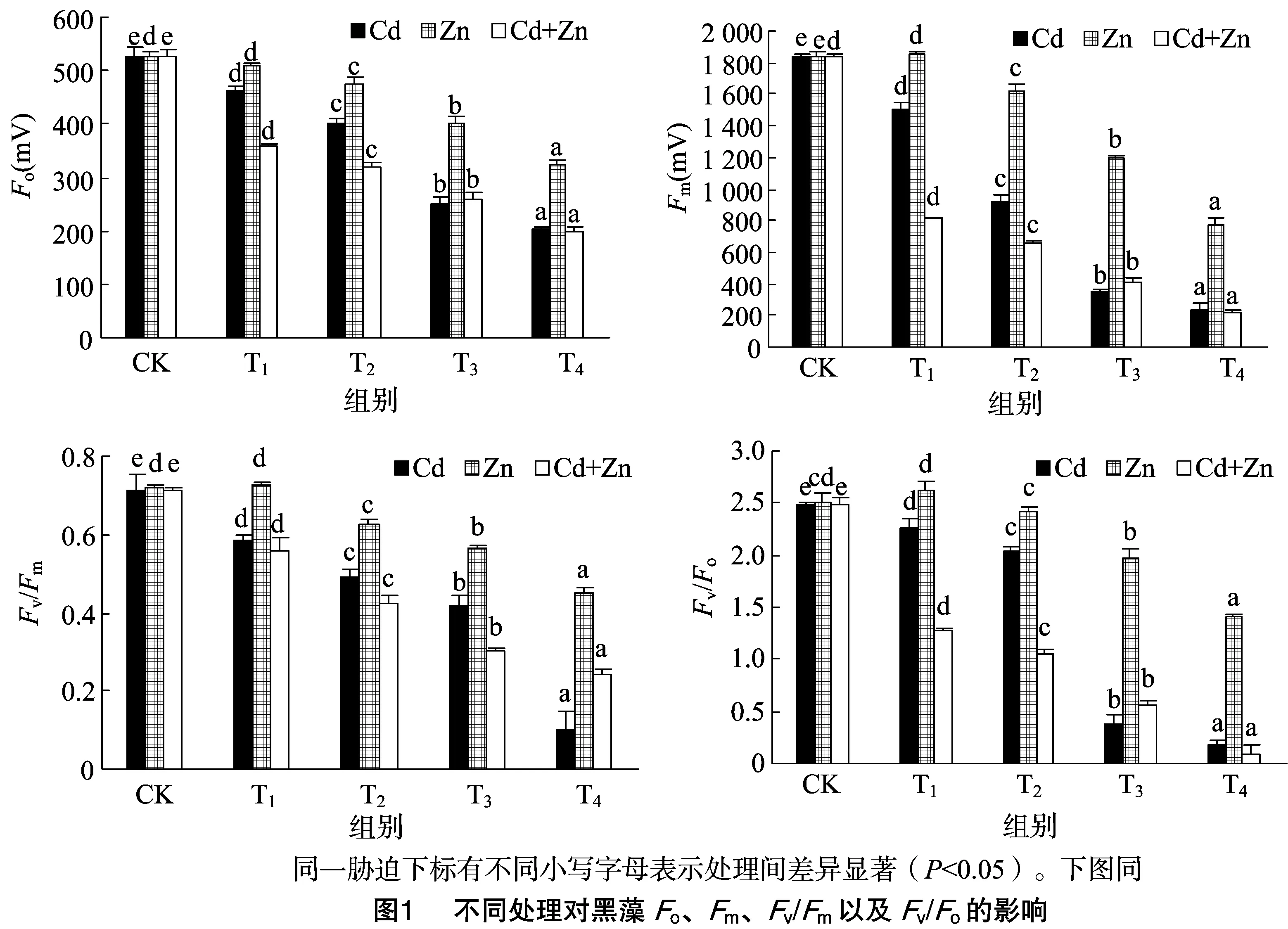
单一Zn胁迫下,Fo和Fm下降幅度最小。而单一Cd胁迫,低浓度(≤2 mg/L)Fo和Fm降低幅度小于相同处理组复合胁迫(Cd+Zn)下的值;而在Cd2+较高浓度(>2 mg/L)胁迫下,单一Cd胁迫下,Fo、Fm与Cd+Zn复合胁迫下的值较接近。这说明低浓度的Cd+Zn复合表现为协同作用。
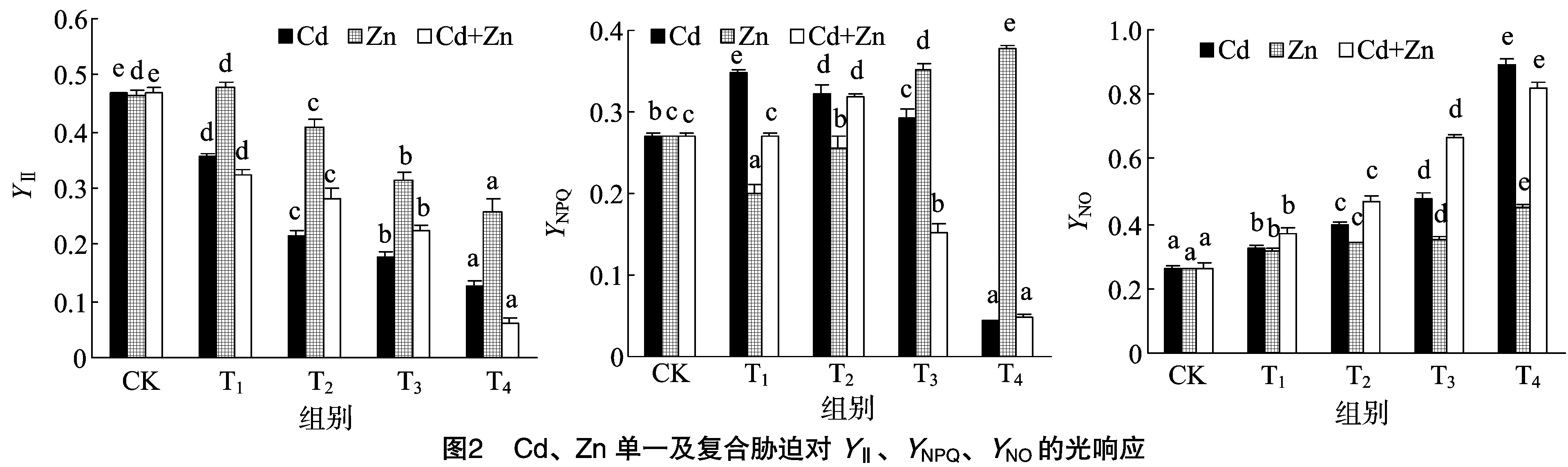
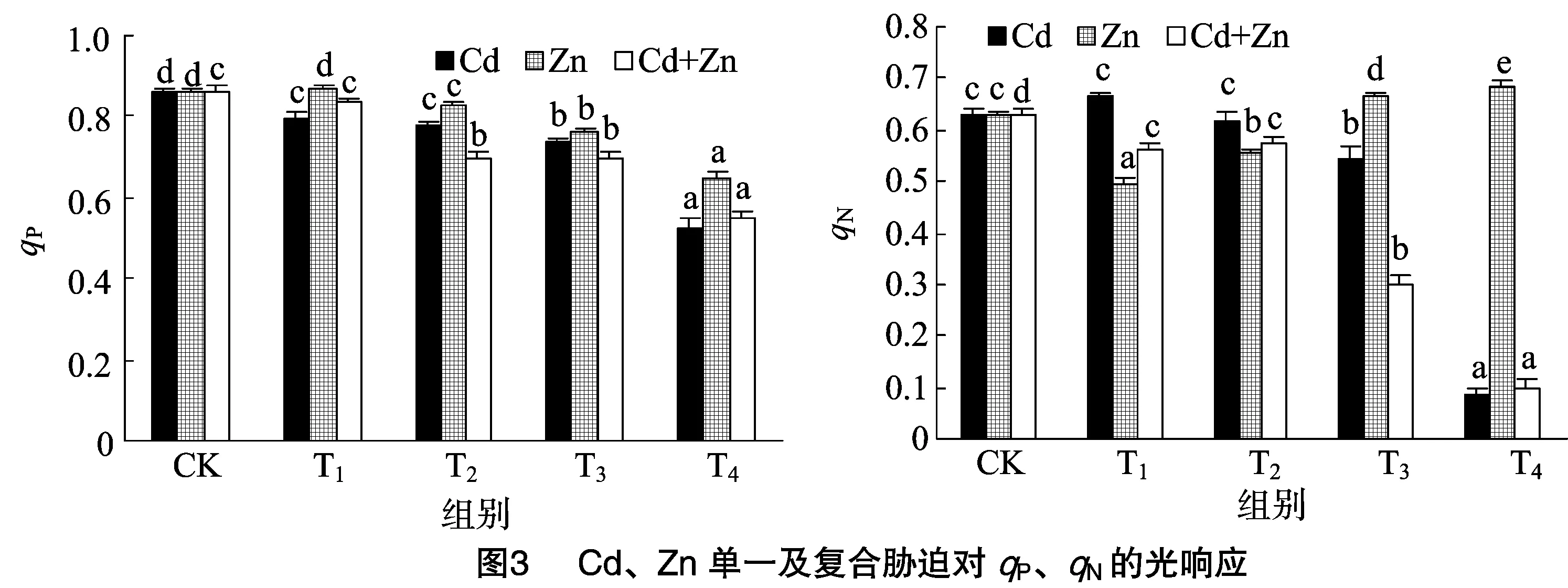
与对照组相比,除单一Zn 3 mg/L胁迫下Fv/Fm略有上升外,其他处理均随着处理组浓度的增大而降低。而低浓度(Cd2+≤2 mg/L、Zn2+≤5 mg/L)时,在相同处理组下,单一Zn胁迫影响最小,Cd次之,复合胁迫降低最大。而在较高浓度(Cd2+>2 mg/L、Zn2+>5 mg/L)下,单一Cd影响最大,单一Zn胁迫影响最小,复合胁迫次之。这表明重金属对黑藻的损害程度表现为:Zn胁迫 Fv/Fo与Fv/Fm呈现相同的趋势,Fv/Fo反映了系统 PSⅡ 潜在活性,除了3 mg/L Zn胁迫Fv/Fo略有升高,其余均随着处理组浓度的增大,PSⅡ潜在光化学效率均呈现不同程度的降低。表明低浓度(3 mg/L)的Zn处理能提高黑藻PSⅡ潜在活性。 由图2可知,随着处理组浓度的增大,无论何种胁迫光系统Ⅱ的有效量子产量YⅡ基本显著减小,但低浓度(≤3 mg/L)Zn单一胁迫时YⅡ略有增加。相同处理组下,复合胁迫使得YⅡ降低幅度最大,达86.99%,单一Zn胁迫降幅最小,单一Cd胁迫降低幅度次之。这表明重金属离子使PSⅡ反应中心受损,抑制光合作用的原初反应,阻碍光合电子的传递过程,金属离子对有效量子产量YⅡ的损害程度表现为:Cd+Zn 复合胁迫>Cd胁迫>Zn胁迫。 图2显示,与CK相比,浓度小于 4 mg/L 的Cd单一胁迫下的YNPQ均显著增加(P<0.05),最高增加了27.94%,出现在Cd浓度为0.5 mg/L处;而当Cd浓度在7 mg/L时,YNPQ却降低了83.82%,可能是较高浓度致死或破坏了植物调节性耗散机制。单一Zn胁迫中,低浓度(≤5 mg/L)降低了YNPQ,而高浓度增加了YNPQ;复合胁迫下,低浓度(Cd2+≤2mg/L、Zn2+≤5 mg/L)增加了YNPQ,而较高浓度(Cd2+>2 mg/L、Zn2+>5 mg/L)则降低了YNPQ。 由图2可知,YNO均随处理组浓度增加而增加。其中,Zn单一胁迫增幅最小,单一Cd胁迫增幅最大,而复合胁迫居中。说明Cd、Zn单一及复合胁迫均对PSⅡ造成一定程度的伤害,导致叶片吸收的部分光量子以非调节性能量耗散,其耗散程度表现为:Cd胁迫>Cd+Zn复合胁迫>Zn胁迫。 qP是PSⅡ天线色素吸收的光能用于光化学电子传递的份额。由图3可知,浓度小于 5 mg/L 的Zn胁迫下qP无明显变化,较高浓度(Zn2+≥5 mg/L)明显降低。随着单一Cd和复合胁迫浓度升高,qP都一直降低,说明PSⅡ中开放的反应中心比例和参与CO2固定的电子减少。 qN反映PSⅡ反应中心对天然色素吸收光能后以热能形式耗散掉的光能部分,其值表示光合机构的损伤程度。由图3可知,随浓度增加,Cd单一胁迫的qN呈先增后降趋势,Zn单一胁迫是先降后增趋势,而复合胁迫则是一直处于降低趋势,很有可能是复合胁迫造成某些机制不可逆转的伤害。 由图4可知,Cd、Zn单一及复合胁迫下,电子传递速率ETR随着光合辐射强度PAR的增加均呈现先增加后下降的趋势,最大值基本出现在PAR为249 μmol/(m2·s)处。同种处理下,随着处理组浓度的增加,ETR最大值明显降低;且相同PAR下,随着处理浓度的增加,ETR逐渐降低,表明随着处理浓度的增加,黑藻的PSⅡ光化学反应受到了抑制。 无论单一胁迫还是复合胁迫,除浓度≤3 mg/L的ZnFo、Fm、Fv/Fm、Fv/Fo胁迫略有升高外(图1),其余总体趋势都是降低的,黑藻叶片PSⅡ反应中心发生了光抑制。Fo降低可能是由Cd、Zn胁迫使得PSⅡ反应中心部分关闭或者失活,即原初电子受体QA全部氧化时的荧光水平降低引起的;也可能是PSⅡ天线色素的热耗散导致Fo的降低。因沉水植物的生活特性,大量重金属加入水体后,重金属的毒性直接作用于植物叶片和茎,植物叶片叶绿体类囊体膜上的蛋白复合体部分失活造成了电子传递和光合磷酸化受阻致使Fm降低。重金属处理后PSⅡ反应中心出现光抑制的现象,PSⅡ电子传递受阻,因此在受到胁迫后Fv/Fm和Fv/Fo明显降低(图1)。 单一Zn胁迫下,黑藻Fo、Fm、Fv/Fm、Fv/Fo的降幅均小于单一Cd及Cd+Zn复合胁迫。表明Zn对黑藻的毒害作用小于Cd+Zn复合,可能因为Zn是植物必需元素,低浓度的Zn能促进叶片光和色素的形成,高浓度则是抑制;Cd能导致气孔关闭,扰乱植物水分状况,降低叶绿素含量和叶绿体数量,减少营养元素吸收,削弱光合作用,并可诱导产生金属螯合肽和结合蛋白等解毒物质,所以才出现Zn毒性 低浓度的Cd、Zn复合对Fv/Fm、Fv/Fo和YNPQ表现为协同作用,原因可能是:(1)低浓度的Zn隔断Cd对硫蛋白生物合成过程的诱导表达信息的传导途径[8],增强了Cd的毒害作用;(2)Zn刺激根部产生更多的转运载体[13],促进了与Zn相似性质Cd的吸收和转运,降低了植物对Cd的控制力。较高浓度表现为拮抗作用,原因可能是:(1)Zn和Cd具有相同的价态(2+),近似相同的离子半径,相似的化学性质,可以相互取代,发生竞争;(2)Cd进入细胞之后,竞争Zn酶中Zn的结合位点,进而取代锌,使锌酶活性降低,甚至完全丧失[8]。 PSⅡ反应中心吸收的光量子主要通过3个途径进行转化与耗散,即YⅡ、YNPQ和YNO,所有占量子产量比例总和始终接近于1,即YⅡ+YNPQ+YNO=1。已有研究表明,Cd、Zn使植物超微结构叶绿体膨胀、叶绿体膜断裂;线粒体脊突膨胀和部分溶解、部分类囊体溶解和散到细胞质中,造成光能的吸收、能量的转换及光合电子传递受阻,从而降低光化学能量途径转化能的比例[14]。在一定浓度范围内,黑藻有能力通过自身的调节机制耗散掉过剩的光能而使PSⅡ反应中心受伤害程度降低,实现自我保护,但超过一定浓度范围后,黑藻的自我调节机制无法承受,致使PSⅡ反应中心受到不可逆转的伤害。 qP反映了PSⅡ天线色素吸收的光能用于光化学电子传递的比例,由图3可知,较高浓度(Zn2+>5 mg/L、Cd2+>2 mg/L)Cd、Zn单一及复合胁迫可以明显抑制黑藻叶片PSⅡ反应中心的开放比例和参与光化学反应电子传递的能量,致使植物光合活性降低。 qN对叶绿体能量状态的改变是非常敏感的,本研究中Zn单一胁迫下qN先降低后升高,说明一定浓度范围(5~10 mg/L)的Zn会刺激黑藻使其光保护能力增强;Cd单一胁迫下qN先升高而降低,说明低浓度(≤2 mg/L)Cd会刺激黑藻的光保护能力增强,但超过一定范围(≥7 mg/L),它的光保护机制会受到不可逆转的损害;Cd+Zn复合胁迫下,qN均是一直降低,说明复合胁迫对黑藻光系统保护机损害强于Cd、Zn单一胁迫(图3)。 本研究中3种胁迫下,在PAR为249 μmol/(m2·s)时,黑藻叶片ETR达到最大饱和,同时随PAR增强呈现平缓下降的趋势,黑藻对光的响应能力随光强增加而减弱(图4)。这意味着PSⅡ反应中心部分关闭或失活,过剩的能量以非光化学淬灭的形式耗散掉,一种形式是用于自身光合作用,另一种是增加热耗散来避免强光对光合系统的伤害[15]。 Cd、Zn单一和复合胁迫均使黑藻受到不同程度的毒害,其中单一Cd及Cd+Zn复合毒害程度严重。Cd污染严重水域通过测定荧光参数来指示污染程度。 低浓度(≤3 mg/L)Zn2+下,黑藻叶片Fm、Fv/Fm、YⅡ等略有增加,可以适当施加一定Zn促进植物光合。 受到不同浓度重金属毒害时,黑藻通过调节PSⅡ反应中心开放程度与活性,实现光保护能力,对Zn胁迫表现出较强的耐性,Cd次之,Cd+Zn复合最小。高浓度重金属胁迫导致PSⅡ反应中心关闭或不可逆失活,表现出光抑制。2.2 Cd、Zn单一及复合胁迫对黑藻YⅡ、YNPQ、YNO的光响应
2.3 Cd、Zn单一及复合胁迫对黑藻qP、qN与电子传递速率(ETR)的光响应

3 结果与分析
4 结论
