斜纹夜蛾保幼激素环氧水解酶基因的克隆和原核表达
2019-01-03张丽丽武怡琼杨正飞魏佳平阚云超
张丽丽,武怡琼,杨正飞,郭 丽,魏佳平,阚云超
(1.信阳师范学院 生命科学学院/大别山农业生物资源保护与利用研究院,河南 信阳464000; 2.南阳师范学院/河南省伏牛山昆虫生物学实验室,河南 南阳 473061)
斜纹夜蛾(Spodopteralitura)属鳞翅目夜蛾科,为全球热带和亚热带主要农业害虫,国内以华南、华东地区发生为重。斜纹夜蛾以幼虫咬食叶片及叶柄,是一种间歇性猖獗发生的害虫[1]。随着近年温室效应导致气候持续变暖以及种植业结构变化、蔬菜种植面积不断扩大,斜纹夜蛾发生危害逐年加重,已成为农作物重要害虫之一[2]。
保幼激素(Juvenile hormone,JH)是昆虫后脑两侧咽侧体合成分泌的一种萜烯类化合物[1]。JH在不同龄期昆虫的血淋巴中通过滴度的变化(合成与代谢)调控昆虫的生长发育,其代谢通过保幼激素酯酶(Juvenile hormone esterase,JHE)、保幼激素环氧水解酶(Juvenile hormone epoxide hydrolase,JHEH)和保幼激素二醇激酶(Juvenile hormone diol kinase,JHDK)3种特异性酶催化完成[3-5]。JHEH属于mEH家族,与组织中的膜结合,催化血淋巴中JH和保幼激素酸(Juvenile hormone acid,JHa)不可逆地转化为保幼激素二醇(Juvenile hormone diol,JHd)和保幼激素酸二醇(Juvenile hormone acid diol,JHad)并中断咽侧体JH合成[3]。
JHEH基因首先从烟草天蛾(Manducasexta)中分离得到[6],之后在鳞翅目昆虫粉纹夜蛾(Trichoplusiani)[7]、家蚕(Bombyxmori)[8]、谷食夜蛾(Helicoverpazea)[9]、棉铃虫(H.armigera)[10]等中分别克隆了JHEH基因,其表达存在差异。粉纹夜蛾和烟草天蛾JHEH基因主要在卵中特异表达[6-7];谷食夜蛾JHEH基因主要在卵巢中表达[9];而家蚕的脂肪体、中肠等几乎所有组织中都有JHEH基因表达[11-12]。Zhou等[13]解析了BmJHEH的晶体结构,该蛋白质是一个典型的α/β水解酶二聚体,N末端的XWG氨基酸结合细胞膜,与亚细胞定位有关。序列分析发现,其功能区氨基酸残基都非常保守,有一个较深的疏水口袋结合JH,其中2个酪氨酸残基Tyr298、Tyr373和HGWP花样结构组成阴氧离子洞,催化三联体Asp227、Glu403、His430氨基酸残基高度保守。半翅目绿盲蝽(Apolyguslucorum)JHEH蛋白结构的模拟结果显示,其存在与家蚕相似的功能结构域,RNAi干扰JHEH基因表达导致绿盲蝽幼虫存活率明显下降,且部分幼虫蜕皮困难,甚至死亡无法延续下一代[14]。半翅目褐飞虱(Nilaparvatalugens)JHEH基因的表达下调对幼虫存活率影响较小,但是显著增加长翅型褐飞虱种群中短翅个体数量,达到总量的50%以上,且以雌性为主[15]。
本研究对斜纹夜蛾JHEH基因(SlJHEH)进行克隆和生物信息学分析,并通过原核表达系统高效表达SlJHEH蛋白,为研究该蛋白质的体外活性和生物功能奠定基础。
1 材料和方法
1.1 试验材料
斜纹夜蛾幼虫来自华南师范大学昆虫研究所。饲养方法:(1)卵消毒: 在卵孵化前1 d,放入5%的甲醛溶液中消毒15 min,用无菌水漂洗,晾干;(2)幼虫饲养: 将初孵幼虫接入塑料培养盒中,待长到3龄时,分盒饲养,直到预蛹期,然后将预蛹幼虫转入干净的饲养盒中化蛹;(3)成虫产卵: 在蛹羽化前2 d,区分雌、雄蛹,将蛹按雌雄配对,置于单独的培养盒中羽化,蛹羽化后,用15%的蜂蜜饲养,以利于产卵。饲养条件:温度25~27 ℃,相对湿度70%~75%,光周期16 h∶8 h (光∶暗)。
RNA提取试剂RNAiso plus、反转录酶M-MLV、DL5000 DNA Marker、PCR常用试剂(dNTPs、10×PCR buffer和rTaq)、DNA限制性内切酶、pMD18-T载体、Solution Ⅰ、T4 DNA ligase、10× T4 DNA ligase buffer、Premixed protein marker(low)均购自TaKaRa公司(大连,中国);原装进口低熔点BIOWEST琼脂糖、质粒提取试剂盒、胶回收试剂盒、SDS-PAGE和Western blotting相关试剂购自郑州宝赛生物科技有限公司;Western blotting所用硝酸纤维素膜为英国Whatman公司产品;大肠杆菌(E.coli) DH5α、BL21(DE3) 感受态细胞和原核表达用pET32a 质粒由华南师范大学昆虫与科学技术研究所冯启理教授馈赠。
1.2 方法
1.2.1 中肠总RNA提取与cDNA的获得 选取斜纹夜蛾6龄进食期3 d的幼虫,按照RNAiso plus说明书提取中肠总RNA后,用Nanodrop紫外分光光度计和1%的琼脂糖凝胶电泳检测RNA的质量,-80 ℃保存备用[16]。按照TaKaRa公司M-MLV RTase cDNA Synthesis Kit说明书合成第一链cDNA,保存于-20 ℃。
1.2.2 引物设计与序列扩增 从NCBI数据库获得SlJHEH全长序列(GenBank登录号:XM_022981795),设计上游引物:5′-GGATCCATGGCGCGCCTGTTGTTTA-3′(划线处为BamH Ⅰ 酶切位点);下游引物:5′-GTCGACTTACAAATCAGTCTTGAC-3′(划线处为SalⅠ 酶切位点)。引物由安徽通用生物技术有限公司合成。以上述反转录获得cDNA为模板,用特异性引物扩增JHEH目的序列。PCR反应程序为:94 ℃预变性3 min;94 ℃变性30 s,59 ℃退火30 s,72 ℃延伸90 s,32个循环; 72 ℃延伸10 min。PCR产物经琼脂糖凝胶电泳分离并回收。将目的片段与pMD18-T 载体连接,转化E.coliDH5α,经过0.1 g/L氨苄青霉素(Amp)筛选单克隆,运用PCR和双酶切鉴定后,送安徽通用生物技术有限公司测序。将重组载体命名为pMD18T-SlJHEH。测序正确的序列用于原核表达载体的构建。
1.2.3 序列分析及系统发育树构建 运用DNAstar分析开放阅读框(ORF)、蛋白质分子质量、等电点。在NCBI 数据库对本研究获得基因推导的氨基酸序列进行同源性搜索; 使用Clustal W2(http://www.ebi.ac.uk/Tools/msa/clustalw2/)和ESPript (http://espript.ibcp.fr/ESPript/cgibin/ESPript.cgi)进行同源性比较[13];通过Mega 6.0软件构建系统发育树[17]。
1.2.4SlJHEH原核表达载体的构建 运用限制性内切酶BamHⅠ和SalⅠ对pMD18T-SlJHEH和pET32a空载体进行酶切,割胶回收目的片段,经T4连接酶16 ℃过夜连接,转化E.coliDH5α,经过氨苄青霉素筛选单克隆,运用PCR和双酶切鉴定,然后送到安徽通用生物技术有限公司测序。将重组载体命名为pET32a-SlJHEH。
1.2.5 SlJHEH重组蛋白的诱导表达 将pET32a-SlJHEH重组质粒转入BL21(DE3)感受态细胞,获得的阳性单菌落接种于LB液体培养基(含0.1 g/L Amp)中,37 ℃、200 r/min培养过夜。按1∶50 接种于5 mL 含Amp的LB培养基中继续培养至OD600=0.6,加入IPTG至终浓度为1 mmol/L,然后于28 ℃、200 r/min摇床中培养4 h,收集1.5 mL菌液,于12 000×g下离心10 min以收集细胞沉淀。用200 μL PBS缓冲液(NaCl 8 g、KCl 0.2 g、Na2HPO41.44 g、KH2PO40.24 g,加水至1 L,pH值7.4) 悬浮沉淀,超声波破碎至液体透明(超声功率为400 W,工作5 s,间隔5 s,重复一定次数),于12 000×g离心10 min,分别收集上清和沉淀。以未重组载体pET32a作为对照,所有试验条件保持一致。
1.2.6 SDS-PAGE蛋白电泳和Western blotting分析 取菌体、上清和沉淀各10 μL,并分别加入10 μL的2×上样缓冲液,100 ℃变性5 min,冰上放置3 min。采用12% SDS-PAGE凝胶电泳分离15 μL的样品,以未重组载体pET32a作为对照,处理条件保持一致。蛋白质凝胶用考马斯亮蓝R250染色,最后成像分析。
蛋白质经SDS-PAGE电泳分离后,电转移至硝酸纤维素膜上,用封闭液(含3%牛血清白蛋白)于37 ℃封闭2 h。然后与His标签抗体(1∶10 000稀释) 于30 ℃下反应1 h,用洗涤液洗膜3次,每次10 min。与碱性磷酸酶标记的羊抗兔IgG酶标抗体 (Anti-IgG AP conjugate) (1∶2 000稀释) 于30 ℃下反应1 h,然后用洗涤液洗膜4次,每次5 min。最后用四氮唑兰(NBT)/5-溴-4-氯-3吲哚-磷酸盐(BCIP)进行显色反应。
2 结果与分析
2.1 斜纹夜蛾中肠RNA的提取
利用RNAiso plus提取斜纹夜蛾6龄进食期3 d中肠的总RNA,Nanodrop紫外分光光度计测得其质量浓度为3 265 ng/μL,A260/A280值为1.942,A260/A230值为2.078。经1%琼脂糖凝胶电泳后,在凝胶成像仪下观察检测总RNA的质量,结果(图1)显示,28S和18S rRNA条带清晰,提取质量良好,无明显降解。
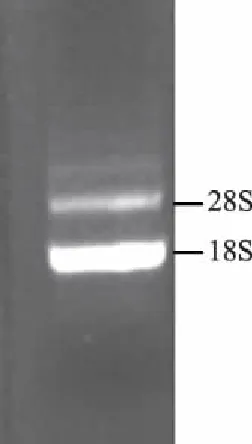
图1 斜纹夜蛾中肠总RNA琼脂糖凝胶电泳
2.2 斜纹夜蛾JHEH基因的扩增与分析
利用特异性引物,以cDNA为模板进行RT-PCR,得到SlJHEH基因片段(图2)。该基因ORF长为1 389 bp,编码462个氨基酸,预测分子质量为52 ku,等电点为8.68。其中有179个疏水性氨基酸和91个极性氨基酸,经分析该蛋白质可能为可溶性蛋白。

M:DNA分子质量标准; 1:PCR产物图2 SlJHEH基因的PCR扩增产物
2.3 斜纹夜蛾JHEH氨基酸序列同源性比对及进化树分析
与其他不同物种的JHEH氨基酸序列同源性比对发现,SlJHEH存在与细胞膜结合的高保守性XWG序列(图3),推测与其亚细胞定位有关[13]。Tyr297、Tyr372和HGWP花样结构氨基酸残基组成疏水性阴氧离子洞,活性位点催化三联体Asp226、Glu402和His429氨基酸残基发挥其水解活性,这2个基本结构是昆虫或哺乳动物环氧水解酶发挥作用的功能性基序[13-14,18],在不同物种中高度保守(图3)。
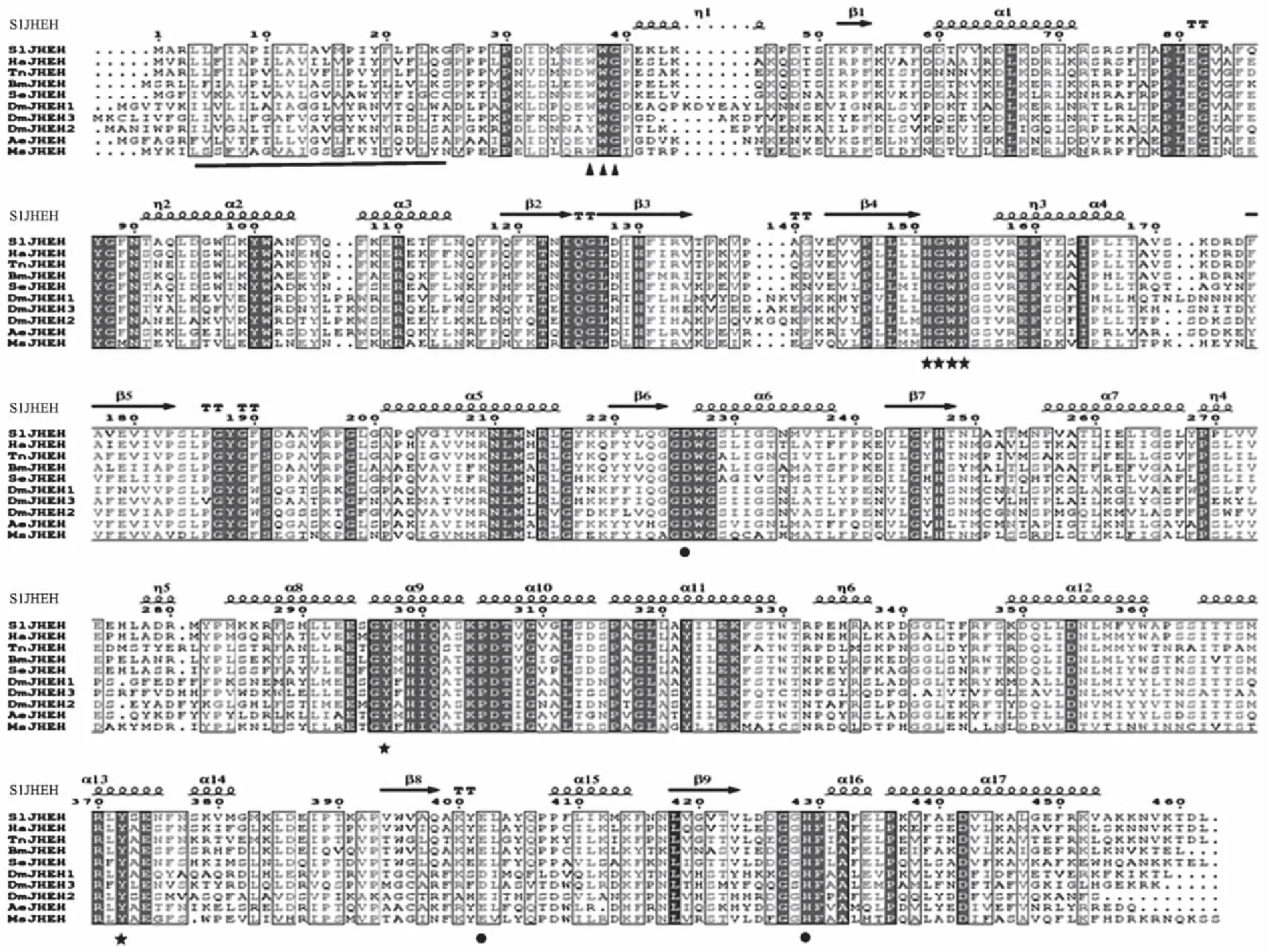
除SlJHEH外,从上至下分别为Helicoverpa armigera(ACM78602)、Trichoplusia ni(AAB88192)、Bombyx mori(NP_001037201)、Spodoptera exigua(ABD85119)、Drosophila melanogaster(ACV04637、NP_611386、AAM88329)、Aedes aegypti(AAM88326)、Manduca sexta(AAC47018)的JHEH蛋白,氨基酸序列均从NCBI数据库下载,图4同;▲标记N端膜结合氨基酸;●标记活性位点催化三联体的Asp226、Glu402、His429残基;★标记组成阴氧离子洞的Tyr297、Tyr372残基和HGWP花样结构
系统发育树分析发现,SlJHEH与鳞翅目的棉铃虫(ACM78602)、粉纹夜蛾(AAB88192)、家蚕(NP_001037201)、小菜蛾(Spodopteraexigua,ABD85119) 和烟草天蛾(AAC47018)的JHEH蛋白很好地聚在一起,尤其与棉铃虫JHEH氨基酸序列同源性高达78%,而与双翅目果蝇(Drosophilamelanogaster,NP_611386、ACV04637和AAM88329)和埃及伊蚊(Aedesaegypti,AAM88326)的亲缘关系较远(图4)。
2.4 重组表达载体pET32a-SlJHEH的构建
将构建的重组表达载体pET32a-SlJHEH转化E.coliDH5α菌株,挑取5个菌落直接作为模板进行PCR扩增,在1 400 bp处均能扩增出单一条带(图5A),随机挑选其中的单菌落扩大培养后提取质粒进行双酶切,在1 400 bp处也出现单一条带(图5B),说明JHEH基因已连入pET32a表达载体。经进一步测序验证,重组表达载体pET32a-SlJHEH构建成功。
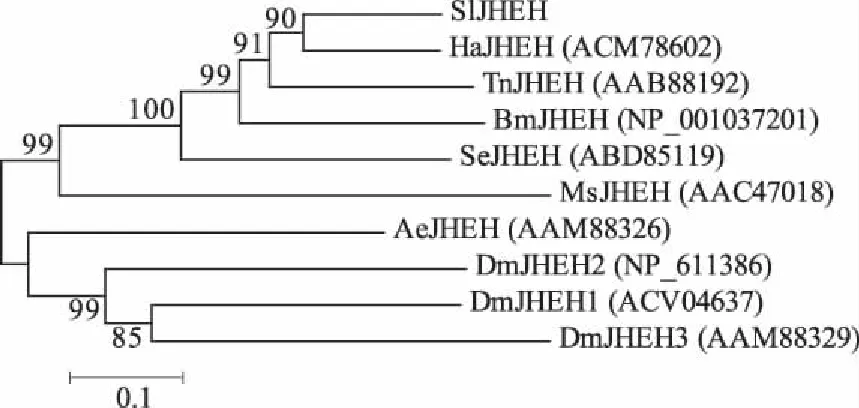
图4 SlJHEH的系统发育树
2.5 斜纹夜蛾JHEH重组蛋白的诱导表达及Western blotting鉴定
表达产物经SDS-PAGE电泳、考马斯亮蓝R250染色后,pET32a-SlJHEH转化菌在分子质量接近66 ku处,比空载体pET32a转化菌多出一条明显的蛋白质条带,表达蛋白为目的蛋白(分子质量52 ku)和pET32a标签(分子质量约18 ku)的融合蛋白,检测结果与预测的融合蛋白分子质量相符(图6A)。目的蛋白主要以包涵体形式存在,上清液中有少量可溶性蛋白(图6A)。采用His标签抗体进行Western blotting分析,进一步确认该蛋白质为与His标签融合的目的蛋白(图6B)。
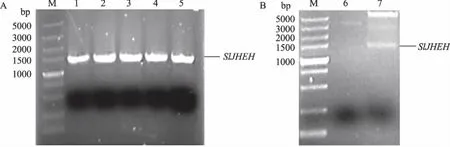
M:DNA分子质量标准; 1—5:菌落PCR产物; 6:pET32a-SlJHEH质粒酶切前; 7:pET32a-SlJHEH质粒酶切后图5 重组表达载体pET32a-SlJHEH的PCR(A)和酶切(B)鉴定
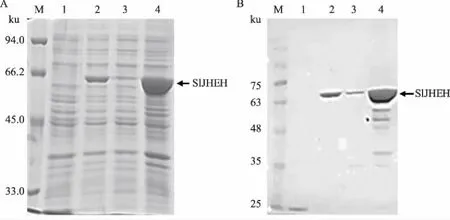
M:蛋白质分子质量标准; 1:pET32a诱导表达产物; 2:pET32a-SlJHEH诱导表达产物;3:pET32a-SlJHEH诱导表达产物上清液; 4:pET32a-SlJHEH诱导表达产物沉淀图6 pET32a-SlJHEH表达产物的SDS-PAGE(A)和Western blotting(B)鉴定
3 结论与讨论
昆虫末龄幼虫到蛹期的变态依赖保幼激素的快速降解[14]。JHEH作为保幼激素降解代谢中的关键酶之一,将JH和JHa降解为JHd和JHad[ 9,19 ]。自从烟草天蛾中克隆出第1个JHEH基因以来[6],又先后在鳞翅目粉纹夜蛾、家蚕、谷食夜蛾,蚤目猫栉(Ctenocephalidesfelis),双翅目果蝇,直翅目蟋蟀(Gryllusassimilis),半翅目褐飞虱、绿盲蝽,膜翅目蜜蜂(Apismellifera),鞘翅目赤拟谷盗(Triboliumcastaneum),同翅目棉蚜(Aphisgossypii)等多个目的昆虫中研究了JHEH基因[7-9,12,14,19-24]。本研究从斜纹夜蛾克隆了JHEH基因的ORF序列,并对其编码的氨基酸序列进行生物信息学分析。SlJHEH基因ORF大小为1 389 bp,编码462个氨基酸。该序列具有明显的JHEH酶的典型特征,即氨基酸序列中具有保守的天冬氨酸(Asp226)、组氨酸(His429)和谷氨酸(Glu402)组成的活性位点催化三联体,HGWP结构域和2个酪氨酸残基(Tyr297和Tyr372)组成其阴氧离子洞,N端有膜结合XWG基序,这些均与家蚕和绿盲蝽JHEH三维结构的研究结果一致[13-14]。
本研究在原核表达系统大肠杆菌中,经过一般诱导表达条件28 ℃诱导4 h,获得大量的目的融合蛋白,分子质量约为66 ku。目的蛋白主要以包涵体形式存在,但是上清中已有少量可溶性的重组蛋白,说明该蛋白质为可溶性蛋白,这与推测氨基酸序列分析结果一致,可进一步改变诱导条件增加上清中可溶性蛋白的表达量。另外,原核表达系统缺乏真核生物中翻译后修饰所需酶类[25],表达出来的可溶性蛋白是否具有生物学活性也需要进一步确认。根据pET32a表达载体的特点,用His标签抗体对诱导表达蛋白进行Western blotting检测,有相应大小的条带出现,表明载体按照正确的翻译框架翻译并表达出带有His标签的目标蛋白。
JH是昆虫特异的激素,研究JH生物合成和代谢途径中的重要酶,对发现更有效的保幼激素类似物和建立利用DNA重组技术的新害虫控制方法有很大帮助。目前,针对烟草天蛾和粉纹夜蛾生物学特性开发的特异性酶抑制剂被认为是潜在的杀虫剂[26-27]。本研究克隆了斜纹夜蛾保幼激素环氧水解酶的基因,构建了重组表达载体pET32a-SlJHEH,并在E.coliBL21(DE3)表达菌株中成功表达,下一步将进行该酶的体外活性研究。
