荒漠草原不同土壤生境猪毛蒿个体大小依赖的繁殖分配
2018-12-21,,,,,*,,,,,,
,,,,,*,,,,,,
(1.宁夏大学西北退化生态系统恢复与重建教育部重点实验室,宁夏 银川 750021;2.宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021;3.宁夏大学农学院,宁夏 银川 750021;4.宾夕法尼亚州立大学公共卫生系,宾夕法尼亚 赫尔希 17033)
荒漠草原处于半干旱与干旱区的边缘地带,是亚洲中部特有的、旱生性最强的草原类型[1],是草原向荒漠过渡的一类十分脆弱的草原生态系统[2],一直是生态、地理、环境和经济学者们关注并研究的热点区域。近些年,随着封育禁牧、退耕还林还草、生态移民等政策的实施,宁夏生态环境得到了明显改善,植被覆盖度显著提高。但由于对草地生态系统的结构和功能缺乏了解,不重视草地资源的科学管理和合理利用,宁夏盐池中部荒漠草原多达1/4的草场以猪毛蒿(Artemisiascoparia)为优势种。猪毛蒿为菊科蒿属植物,因具有较强的适应恶劣环境能力而成为主要先锋物种[3]和优势建群物种[4-5],单个样地覆盖度可达50%[6],其含有挥发性物质,气味浓并有苦味,适口性差,其他植物生长受到很大抑制[7],导致草地利用率低[8]。因此,在荒漠草原像猪毛蒿这种非优质牧草,往往容易被忽视,确实是值得引起重视的研究内容。
植物在生长过程中,难免会受到外界环境以及内部资源状况的影响。植物对环境的适应策略是植物在长期的适应环境过程中,调节和改变自身而逐渐形成的一系列适应性特征[9],一直是生态学研究的核心问题[10]。其中,植物繁殖对策是通过调整资源分配模式,以最佳的分配策略适应多变的环境[11],提高植物自身适合度的组织过程[12],是植物生活史过程中最为关键的环节[13],也是生物群落演替和维持生态系统功能的基础,对于揭示植物生长、生殖和生存之间协同进化关系具有重要意义和应用价值。研究植物在不同环境中的繁殖对策,能够反映出植物对环境的适应能力和在该环境中的生殖潜能[14],可为理解该地区物种多样性的形成机制提供有价值的参考[15]。特别是近年来,随着生活史进化理论研究的发展,植物的繁殖对策已成为植物生态学的研究热点,早已引起生态学界的高度重视[16-17]。
植物个体大小与繁殖分配的关系被认为是生活史理论中最基本的研究内容,近些年来,对植物的繁殖分配以及资源权衡已有许多的理论和实验研究[18],大部分学者认为在不同的海拔和密度生境下,随着植株个体增大,繁殖绝对投入会增加,但繁殖相对投入(即繁殖分配比例)则会减小[19-21]。但孟丽华等[22]在海北高寒草甸研究发现,不同海拔梯度圆穗蓼(Polygonummacrophyllum)的繁殖分配与个体大小并不存在依赖效应。以往研究多集中在不同海拔梯度下植物个体大小和繁殖分配的关系上[18],然而,在影响植物繁殖分配的诸多因子中,土壤类型的差异也会对植物的繁殖分配和生活史策略产生重要影响。目前,关于荒漠草原不同土壤类型下猪毛蒿的个体大小与繁殖分配的关系,尚无相关研究报道。因此选择不同土壤类型条件下的猪毛蒿种群作为研究对象,能更准确地了解猪毛蒿种群的繁殖分配策略。
宁夏荒漠草原面积约为1.4×104km2,占全区草地面积的42%[23],地带性土壤为灰钙土,非地带性土壤主要是风沙土[24],裸露的松散基岩风化残积土也普遍分布。在这种异质性生境下,准确掌握草地资源既是合理开发和利用草地的基础,也对生态环境保护具有重要意义。猪毛蒿属广布耐旱植物,近些年来在宁夏荒漠草原迅速扩散,其数量增多是草地退化的标志,有研究结果表明底层土壤质地是决定猪毛蒿群落的构建的主要因子[25]。但由于对其繁殖对策了解甚少,人们尚未找到治理猪毛蒿的有效方法。本研究重点探索以下几方面问题:猪毛蒿的生物量及其分布特征在异质生境下的变化规律;猪毛蒿的个体大小对投入到繁殖的绝对值和繁殖分配的影响;猪毛蒿不同土壤生境下是否均存在繁殖阈值及其繁殖阈值在异质生境下的变化情况,以期对进一步探究猪毛蒿的生态防除提供一些理论依据。
1 材料与方法
1.1 研究区概况
宁夏中部荒漠草原,处于宁夏中部干旱带上,地貌为鄂尔多斯缓坡起伏高原。干旱少雨,蒸发量大,冬春两季风大沙多,属典型的中温带大陆性气候。近50年平均气温为8.5 ℃,年降水量为276.3 mm,年日照时数为2862.6 h。该区域植被主要受水分因素限制[26],物种多样性较少、群落结构简单、沙生特征明显。地表水与地下水资源匮乏。20世纪60年代以来,由于人类活动加剧,土地沙化面积仍不断扩大。
1.2 研究材料
猪毛蒿为菊科蒿属植物,又名滨蒿、东北茵陈蒿、土茵陈,一年生、二年生或多年生草本,广布于欧、亚大陆温带至亚热带地区。在半干旱或半温润地区的山坡、林缘、路旁、草原、黄土高原、荒漠边缘地区都有,局部地区构成植物群落的优势种。主根单一,狭纺锤形、垂直,半木质或木质化,根状茎粗短。茎稀2~3枚,红褐色或褐色,有纵纹;常自下部开始分枝,下部分枝开展,上部枝多斜上展,枝上密生叶,茎、枝幼时被灰白色或灰黄色绢质柔毛,以后脱落。叶近椭圆形、长卵形,二至三回羽状全裂,具长柄,花期叶凋谢;茎下部叶初时两面密被灰白色或灰黄色略带绢质的短柔毛,后毛脱落。头状花序近球形,稀近卵球形,极多数,直径1~2 mm。7-8月开花,9-10月结实。瘦果倒卵形或长圆形,褐色。初秋种子萌发以幼苗越冬,来年逢雨迅速生长[27-28]。本试验中研究对象为当年春萌苗生长的植株。
1.3 取样及测定方法
在宁夏中部荒漠草原区应用随机取样的方法,选取39个典型的猪毛蒿群落样地,样地基本信息见表1。所有的猪毛蒿植株样本于2017年10月4-8日进行采集,先用GPS定位并记录采样点的经纬度和海拔信息,然后在每个样地设置3~5个样方,随机选取生长健康、没有人为干扰和病虫害影响的猪毛蒿个体,用卷尺测量地上部分高度和冠幅后挖取整株,根部清理干净后,分株装于信封袋带回实验室。基岩风化残积土、风沙土和灰钙土生境下总取样量分别为20、105和65株。土壤硬度采用数显式土壤紧实度仪(USA,SC-900)进行测量;土壤水分采用便携式土壤水分测量仪(ML3,England)进行测量;盖度测量选择常用的目测估算法;密度为实际测量值。测量猪毛蒿植株特征前再次清理,去除杂草、小石块等杂质,把植株分为头状花序、根、茎和叶等部分,分别装入信封并置于65 ℃烘箱中烘至恒重,然后用万分之一电子天平进行称重。
1.4 数据分析
对个体大小应该用营养器官生物量还是地上部分总生物量来衡量并不统一,本研究中猪毛蒿个体大小以地上部分生物量干重为准[16];繁殖器官大小以头状花序干重为准;营养器官是根、茎和叶的干重之和;株高为主茎最上端与地面间的自然高度;冠幅为东西向长度和南北向长度的乘积;繁殖分配为繁殖器官干重占植物个体总生物量干重的比例。在位置效应分析中,将植株按株高平均分为上部、中部和下部[29],分别统计单株猪毛蒿各部位的生物量、繁殖体数量、繁殖体重量,并计算单个繁殖体重量,单个繁殖体重量为单株繁殖体重量与单株繁殖体数量的比值。采用经典的异速方程:Y=βXα进行异速增长关系分析,个体大小(V)与繁殖器官(R)的数据进行对数转换以去除变量偏差对回归系数的影响,然后进行统计分析相关性[30]。
采用Excel 2016和SPSS 21.0软件进行统计、计算和制图,采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)分析不同生境各特性、猪毛蒿生长指标、生物量等方面的差异。
2 结果与分析
2.1 异质生境
猪毛蒿生长的3种土壤生境下0~20 cm土壤水分存在明显的差异(表2),65个灰钙土测量点的平均含水率极显著高于风沙土(105个测量点)和基岩风化残积土(20个测量点)(P<0.01)。0~20 cm平均土壤硬度也存在显著差异(P<0.05),灰钙土平均硬度为1779.01 kPa,基岩风化残积土则为1412.56 kPa,风沙土最小,仅为506.18 kPa。调查样地中植被的总盖度无显著差异(P>0.05),但基岩风化残积土中猪毛蒿的盖度显著低于其他两种生境(P<0.05)。春萌猪毛蒿的密度间无显著差异(P>0.05),但灰钙土中的秋萌苗达到了227.23株·m-2,显著高于风沙土(135.69株·m-2)和基岩风化残积土(88.47株·m-2)(P<0.05)。
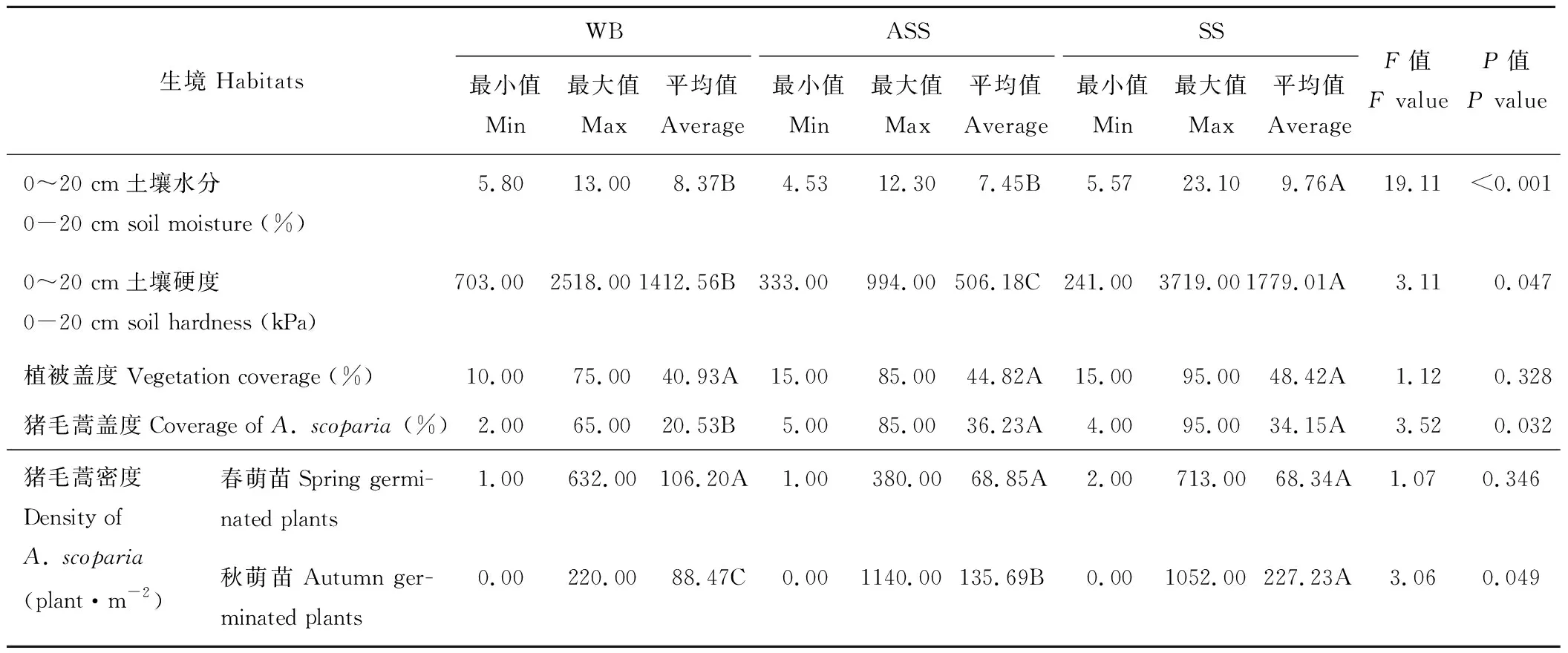
表 2 猪毛蒿不同生境特性的比较Table 2 Comparisons of the heterogeneous habitats properties of A. scoparia
注:同行不同字母表示不同生境间差异显著(P<0.05)。
Note: Different letters in the same row indicate the significant difference in the heterogeneous habitats (P<0.05).
2.2 生长指标及生物量特征
分析结果显示(图1),灰钙土生境下猪毛蒿平均株高(28.1 cm)略大于风沙土(27.9 cm),但差异未达到显著水平(P>0.05),与基岩风化残积土生境下(19.1 cm)差异显著(P<0.05)。灰钙土和风沙土中猪毛蒿平均冠幅高出基岩风化残积土的63.71%和42.48%,但差异未达到显著水平(P>0.05)。而灰钙土中茎粗平均值为3.84 mm,风沙土为2.95 mm,分别显著高出基岩风化残积土中猪毛蒿平均茎粗的1.46和0.89倍(P<0.05)。平均单株生物量差别较大,灰钙土中高达10.60 g,风沙土中为7.07 g,基岩风化残积土仅为1.28 g,但经方差分析,各生境下单株生物量未达到显著差异(P>0.05)。
2.3 生物量间的相关关系
所调查的猪毛蒿个体中营养器官平均重4.52 g,繁殖器官为3.73 g(图2)。其中,基岩风化残积土生境下营养器官平均重仅为0.69 g,繁殖器官为0.59 g;风沙土中猪毛蒿营养器官平均重3.81 g,繁殖器官为3.25 g;而灰钙土生境下猪毛蒿的营养器官平均重5.78 g,繁殖器官则为4.82 g。基岩风化残积土中单株猪毛蒿的生物量较小,营养器官和繁殖器官生物量也较小,风沙土和灰钙土多集中在0~20 g。总体来看,荒漠草原中猪毛蒿的繁殖器官和营养器官生物量间呈线性正相关关系(R2=0.972,P<0.001)。
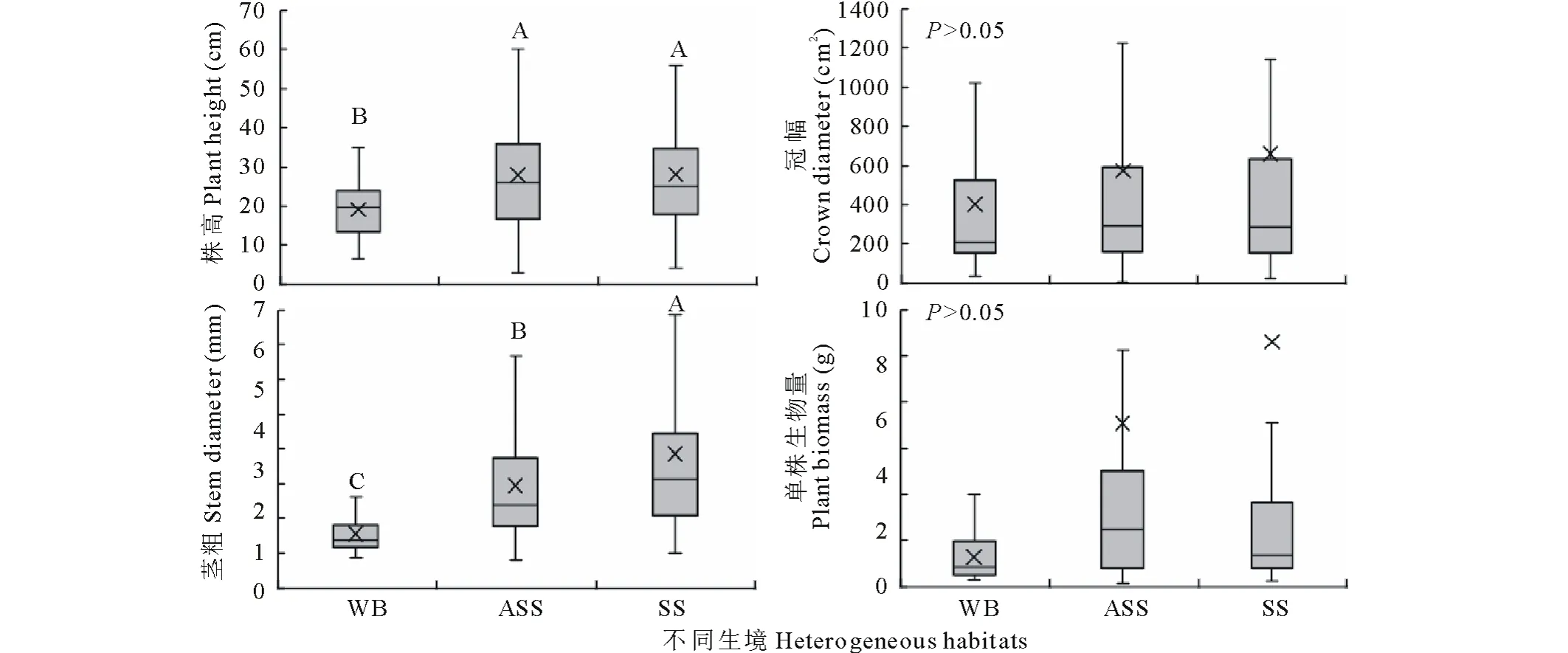
图1 不同生境下的猪毛蒿生长指标及生物量特征Fig.1 The characteristics of growth and biomass in heterogeneous habitats 符号“×”表示平均值,不同大写字母表示不同土壤类型间差异显著(P<0.05),下同。The symbol “×” indicate the mean value in the graph, the different capital letters indicate significant differences at the 0.05 level, the same below.
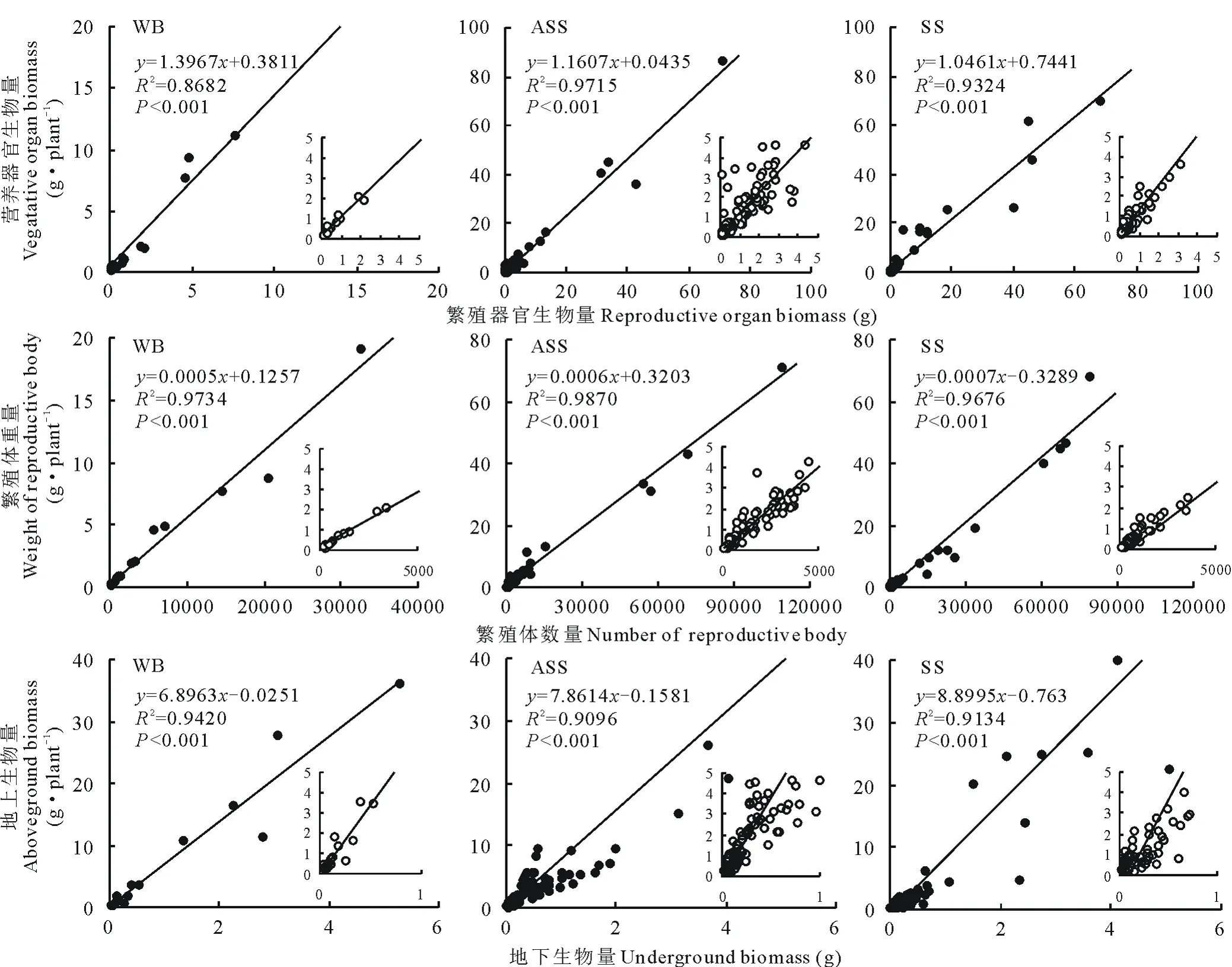
图2 猪毛蒿各组生物量之间的线性回归关系Fig.2 Linear regression relationship between each biomass group of data of A. scoparia
从图2中可以看出,不同生境下猪毛蒿的繁殖体数量主要分布在0~30000个,单株繁殖体重量多集中在0~20 g。其中,基岩风化残积土生境下单株猪毛蒿的繁殖体平均个数仅为941.47个,平均繁殖体重量也仅为0.59 g;风沙土中单株猪毛蒿繁殖体为4700.94个,重量平均为3.25 g;而灰钙土中繁殖体个数高达7283.31个,繁殖体平均重4.82 g。单株猪毛蒿繁殖体数量越多,繁殖体的重量也就越多,存在着显著正相关关系(R2=0.986,P<0.001),说明猪毛蒿的繁殖体数量和质量间也存在着正向权衡。
而单株猪毛蒿地上生物量多集中在0~10 g,平均值为7.32 g,地下生物量则在0~2 g,平均值为0.94 g。基岩风化残积土中猪毛蒿平均地上生物量为5.96 g,地下生物量则为0.87 g;风沙土中地上生物量平均值为6.26 g,地下生物量则为0.82 g;灰钙土中地上生物量平均值为9.45 g,地下生物量则为1.15 g。可以看出,随着地下生物量的增大,地上生物量也呈线性增加,分析表明单株猪毛蒿地上生物量和地下生物量间存在着极显著正相关关系(R2=0.953,P<0.001),同样说明猪毛蒿的地上和地下生物量间也存在着一定的权衡。
2.4 位置效应
猪毛蒿不同部位的地上生物量、繁殖体数量、繁殖体重量和单个繁殖体重量变化情况见图3。单株地上生物量平均重大小为:风沙土>灰钙土>基岩风化残积土,但3种生境下差异不显著(P>0.05);风沙土和灰钙土生境下地上生物量不同部位平均重表现出下部>中部>上部的位置格局,而基岩风化残积土中则表现出中部>下部>上部,但各部位间未能达到显著差异(P>0.05)。繁殖体重量和单株繁殖体数量也表现出相同的趋势,同样经过方差分析,不同生境下和各部位间均差异不显著(P>0.05)。繁殖体重量中,基岩风化残积土、风沙土和灰钙土生境间分别为1.87、2.52和2.18 mg,无显著差异(P>0.05);基岩风化残积土和灰钙土生境下猪毛蒿上部、中部、下部单个繁殖体重量分别为0.62、0.64、0.60 mg和0.75、0.76、0.67 mg,均表现出中部>上部>下部,而风沙土中上部平均重为0.89 mg,高于中部(0.85 mg)和下部(0.78 mg),但不同生境下猪毛蒿各部位单个繁殖体重量间均差异不显著(P>0.05)。
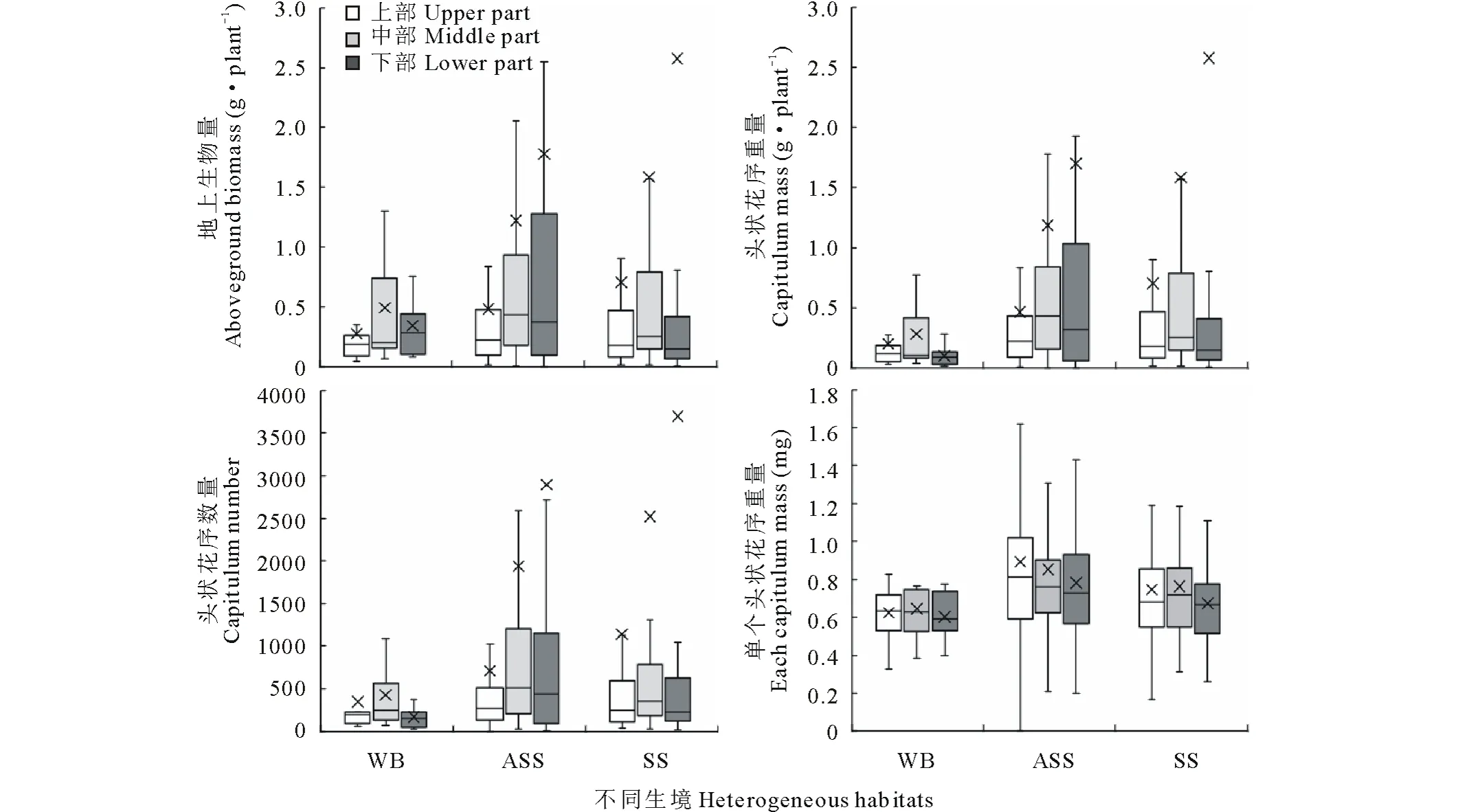
图3 不同生境下的猪毛蒿个体大小和繁殖器官格局特征Fig.3 The structure characteristics of aboveground biomass and reproductive body of A. scoparia in heterogeneous habitats
2.5 个体大小依赖的繁殖器官绝对投入量
根据繁殖器官生物量与个体大小存在的异速增长效应[20, 31],按照侯勤正等[18]的分析方法,对繁殖器官生物量与个体大小进行对数转换后,讨论两者间的线性关系(图4)。结果显示,猪毛蒿的繁殖器官绝对投入量(lgR)在3种生境下均随着个体大小(lgV)的增大而增加,且呈现出极显著的正相关关系(P<0.001)。当y轴为0时,基岩风化残积土、风沙土和灰钙土生境下x轴的截距(图中虚线)分别为0.316、0.292和0.299,表明猪毛蒿繁殖需要一个最小的个体大小,即存在繁殖阈值,所对应繁殖阈值的个体大小分别为2.07、1.96和1.99 g。
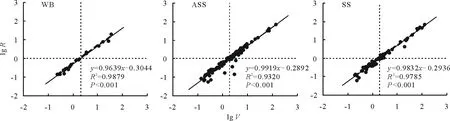
图4 不同生境下猪毛蒿繁殖器官生物量(lg R)和个体大小(lg V)的相关性Fig.4 Correlation analysis between aboveground biomass (lg V) and reproductive biomass (lg R) of A. scoparia in heterogeneous habitats
2.6 个体大小依赖的繁殖分配
整体来看(图5),基岩风化残积土生境下猪毛蒿的繁殖分配(0.494±0.082)略低于风沙土(0.536±0.129)和灰钙土(0.516±0.105),但3者间均未达到显著差异(P>0.05)。从猪毛蒿繁殖分配和个体大小的关系来看,3种生境下均呈现出负相关关系,经Pearson相关性分析,基岩风化残积土、风沙土和灰钙土生境下的相关系数分别为-0.327、-0.045和-0.024,但均未达到显著相关水平(P>0.05)。
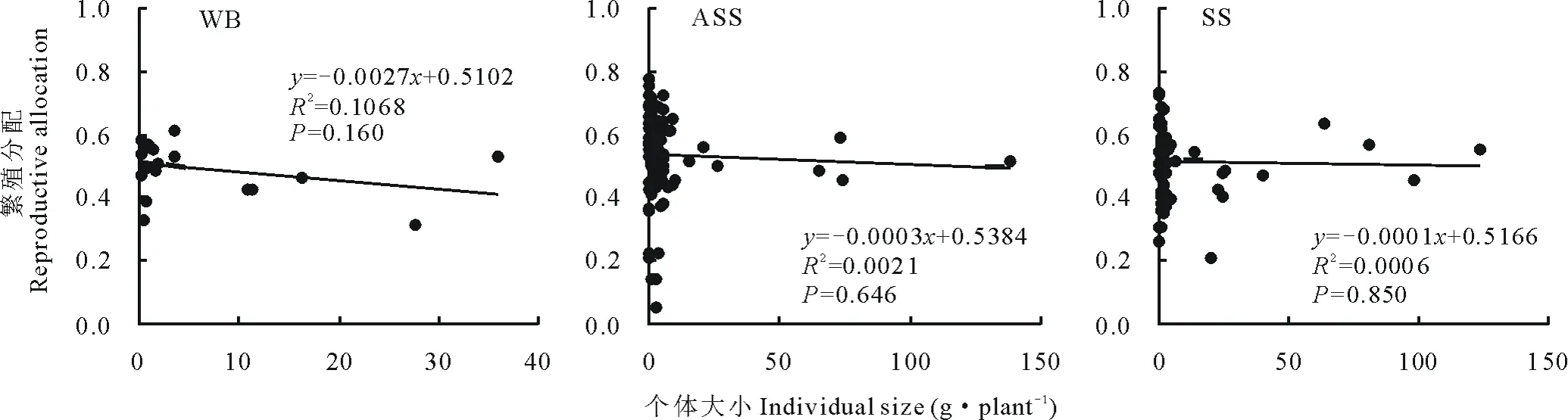
图 5 异质生境下猪毛蒿植株个体大小与繁殖分配的关系Fig.5 Correlation analysis between individual size and reproductive allocation of A. scoparia in heterogeneous habitats
2.7 生物量的权衡
通过图 6可以发现,不同土壤生境下单个头状花序重量大小排序为:风沙土(0.812 mg)>灰钙土(0.722 mg)>基岩风化残积土(0.614 mg)。不同生境下猪毛蒿单个头状花序重量和个体大小、单个头状花序重量和每株头状花序数量间呈负相关关系。虽然大量研究已经发现,花的数量和大小之间存在权衡关系[32],但也有学者研究发现多年生禾本科植物Amphibromusscabrivalvis的这种权衡仍未被有力证明[33],这与本研究对猪毛蒿的研究结果相一致。
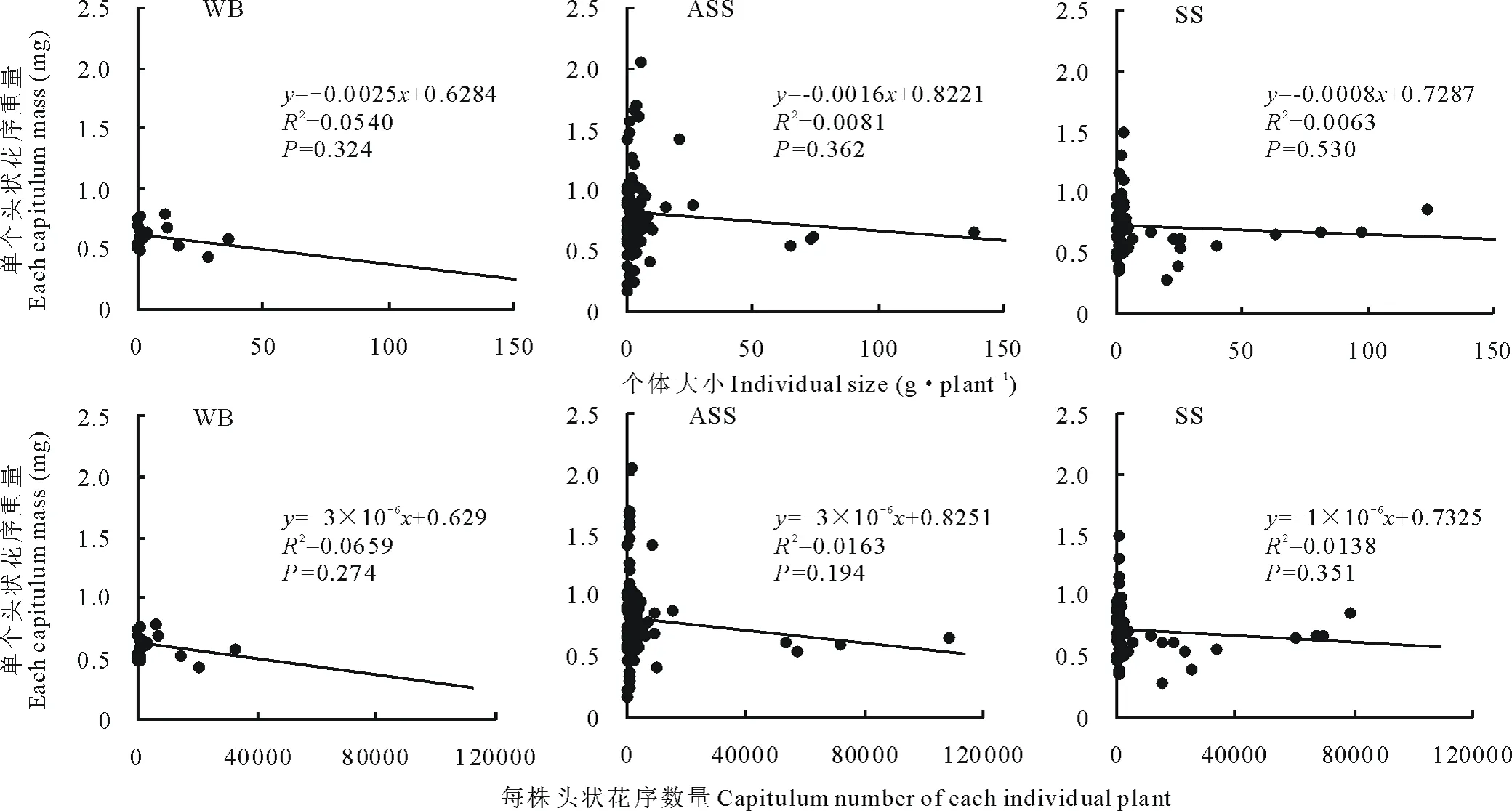
图6 单个头状花序重量与个体大小、每株头状花序数量的线性回归关系Fig.6 Liner regression relationship of capitulum mass with individual size and capitulum number
3 结论与讨论
3.1 不同生境下的个体大小及其位置效应
植物在其生长过程中植株个体之间总要产生大小差异,个体大小的表型可塑性是植物适应异质环境的重要方式,可使植物适应不确定环境或持续变化的环境[32,34]。由于较大的个体面临的生存危险远大于个体较小者,小个体更适宜于恶劣的生态环境,从而在竞争中取得优势,这也是植物在长期进化过程中形成的适应环境的生活史对策[35]。荒漠草原的生境高度异质性,生态因子的波动不稳定性,都会对植物的生长繁殖造成胁迫。通过对不同土壤生境特性的实地调查,发现基岩风化残积土由于底层(>20 cm)是较为坚硬的母岩,猪毛蒿根系延伸和生长繁殖受到了一定的限制,因此,其地上生物量和地下生物量、繁殖器官和生殖器官、繁殖体数量和重量等方面均小于风沙土和灰钙土生境下。
通常情况下,植株不同部位甚至同一花序不同位置的小花、果实和种子的数量、大小、形态及繁殖效率可能不同[36],存在明显的位置效应[21]。多数研究表明种子的大小或数量沿花序由下而上或先开的花到晚开的花递减,这种差异是植株个体水平上提高整体适合度的繁殖对策[37],在繁殖资源受限时植物会通过权衡繁殖资源分配比而形成差异化的分配格局[29],也有研究发现植株中部或基部晚开的花有较高的果实、种子形成率[38]。猪毛蒿的头状花序近球形,并排成复总状或复穗状花序,在茎上再组成大型、开展的圆锥花序,研究其繁殖体的位置效应,并比较植株不同部位的繁殖资源分配的差异,对其种群适应机制的研究具有重要意义。本研究中,不同生境下猪毛蒿未表现出资源投入的差异性,其单株不同部位的地上生物量、繁殖体数量、繁殖体重量和单个繁殖体重量均无显著差异(P>0.05),这与Zimmerman等[39]研究发现一种新热带兰花(neotropical orchid)果实和种子的形成是不受位置效应影响相一致的。目前解释植物资源投入差异主要有资源竞争、结构效应和花粉限制3种假说[37],但Brunet[40]对广泛分布在北美洲西部落基山脉南部和中部的多年生毛茛科草本植物Aquilegiacaerulea研究认为,以上3种假说并不能全部解释基部花较低的坐果率和结实率,而将这一现象更多地归因于基部花的花粉产量和传播在形态学和功能上的偏雄性特化。而猪毛蒿对生境资源的高效利用,不同位置的花获得资源与花序轴维管组织的发育状况及大小没有显著差异,以及不同部位的花由于自交和异交在数量和质量上是否有显著差异,是否是猪毛蒿头状花序并未表现出显著位置效应的原因,这种生理学机制及其与生境的关系还有待进一步研究证实。此外,菊科植物种子多有异型性[41],猪毛蒿异型种子质量如何,萌发和生长是否具有差异,是否存在位置效应,也是今后值得研究的内容。
3.2 个体大小依赖的繁殖分配
由于自然种群中植物存在大小等级性[30],对繁殖投入与个体大小的关系又成为繁殖分配研究中生态学家关注的焦点[42],分析个体大小依赖的繁殖策略是理解植物繁殖资源分配的关键[20]。而异速模型通常被认为是植物繁殖绝对投入和个体大小的最适关系[43]。荒漠草原猪毛蒿的繁殖器官生物量的绝对投入(lgR)都随着个体大小(lgV)的增大而增加,表现出极显著正相关关系(P<0.001)。说明猪毛蒿能同时增加对营养生长和繁殖的资源投入,不同生境主要作用于它的个体大小,并不直接作用于繁殖器官的大小,这和侯勤正等[18]对青藏高原湿生扁蕾(Gentianopsispaludosa)的研究结果相一致。
以往较多的研究多集中在海拔因子,但海拔并非是影响繁殖分配模式的唯一因子,环境中的其他因子同样影响植物的繁殖分配模式[22]。针对不同土壤生境条件下,植物繁殖分配的相关研究还较少。由于植物存在个体大小依赖的繁殖分配的前提假设是植物可用资源的有限性[44],而本研究地点以基岩风化残积土、风沙土和灰钙土为主,土壤发育年轻,薄层性和粗骨性强,养分含量差别相对较小,因此未表现出显著的差异。这表明在荒漠草原,猪毛蒿的繁殖分配没有受到土壤类型条件的明显限制,具有较广的适应能力,既能生活在结构松散的风沙土上,也能适应硬度较大的灰钙土和基岩风化残积土生境条件。但该区域植物生长受限的主要因子是水分,今后应当对不同土壤水分条件下猪毛蒿个体大小依赖的繁殖分配进行相关研究。因此,不同土壤水分条件和资源利用效率对荒漠草原猪毛蒿的繁殖分配的影响将是未来研究的重要方面。
进行繁殖的植株个体大小存在一个阈值[45],这已经在许多案例中被证实[43,46-47]。但也存在例外,赵方等[48]对不同海拔居群中华山蓼(Oxyriasinensis)、赵志刚等[30]对5种毛茛科植物的研究,并没有发现繁殖阈值的存在。猪毛蒿繁殖器官生物量极显著地随着地上个体大小的增大而增大,这与前人的研究结果相一致[16],且繁殖阈值较低。Fan等[49]对珠芽蓼(Polygonumviviparum)的研究结果表明,在恶劣的环境中植物的繁殖比营养生长具有更为重要的地位。本研究同样也表明猪毛蒿在较少的营养积累下就可以达到开花繁殖状态,说明其繁殖比营养生长具有更为重要的作用和意义。这也证明了植物亲代个体对子代的贡献就是尽可能获得高的繁殖输出,使得个体的适合度也就越大[42]。
而不同生境同种植物其繁殖阈值通常存在差异,表明繁殖阈值与环境因子密切相关[22]。湿生扁蕾(G.paludosa)在生境从嵩草(Kobresiasp.)草甸经交错带到金露梅(Potentillafruticosa)灌丛的过渡中,湿生扁蕾所获得的有效资源逐步减少的,其繁殖所需的个体大小的阈值也呈现出递减趋势,这表明植物繁殖所需的个体大小阈值会随其生境可获取的有效资源量的增加而变大[18]。岩白菜(Bergeniapurpurascens)个体大小繁殖阈值在林线以下会随海拔升高而增加,而在林线以上则差异不显著,推测阈值的变化与植株可获得的资源量有关,生境稳定则繁殖阈值相对稳定[43]。本研究中,荒漠草原不同生境下猪毛蒿繁殖阈值差异不大,说明它在不同生境下均可以完成后代繁殖,可能是由于调查土壤类型生境的差异性较小,不足以使得它的繁殖阈值发生变化。
3.3 生物量间的权衡关系
植物在不同的选择压力下,通常以权衡(trade-offs)的方式将有限的资源合理地分配给不同的功能或器官[50]。植株体内资源分配的权衡关系是植物生态学策略的一个普遍特征[51]。自然环境中可供植物利用的资源是有限的,当一定的资源分配给某个器官或行使某种功能时,就不能被其他的器官或功能所利用[52],而植物的每个生活史性状在有限生态资源条件下都达到最佳是无法实现的,植物必须权衡这些性状间的资源分配,才能更好地适应复杂多变的环境[32]。有研究表明,在植物生长的后期,植物将会选择增加繁殖投资来确保有性生殖的顺利进行[32],采取将资源尽可能地集中到繁殖体,以确保后代的延续,而且有些植物会选择产生更少而更大的种子来确保每粒种子的生存能力[53],本研究中,大小依赖的繁殖分配、头状花序重量以及单株头状花序数量间的关系均属于猪毛蒿对这些性状的权衡,表现出了一定的趋势,但猪毛蒿的这些权衡间没有显著的分配关系,也可能是邻体效应与繁殖分配的关系在个体水平上仍不明了的原因,也就是说邻体效应(如本研究区内广泛分布的人工柠条林)是否会影响植物(猪毛蒿)个体的繁殖分配,以及植物在不同邻体效应下将采取何种繁殖对策,目前尚无明确的结论[16]。同时,不排除和本研究的取样量有关,个体大小>20 g的样本数相对较少,会导致拟合方程和显著性检验的结果未能很好地表现,今后将加大样本数量进行综合分析。
此外,根据野外观察发现,猪毛蒿的大量结实和多次萌发特性的叠加使种群增加了适应荒漠草原恶劣多变环境的途径,这可能是猪毛蒿长期对选择压力作出积极反应,也是对生境长期适应的结果,使得其在荒漠草原中具有适应优势,保证种群的延续。
