榆林市2种引栽绿化树种的抗寒性分析
2018-12-11李鲜花赵欢蕊
刘 娟,李鲜花,赵欢蕊
(榆林学院生命科学学院,陕西榆林719000)
在低温条件下,植物体内会发生一系列错综复杂的反应,导致植物体内的生理指标发生变化,因此,可以通过对其生理指标的测定来反映植物的抗寒性。有研究表明,植物对低温的反应与植物体内的游离脯氨酸含量、丙二醛含量和过氧化物酶活性密切相关。
在正常情况下,脯氨酸含量并不高,然而在植物遭受低温影响时,脯氨酸含量增多,其能够作为抗寒性的生理指标[1]。吕优伟等[2]对早熟禾在低温胁迫下的生理响应及苗期抗寒性研究中认为,游离脯氨酸含量随着温度的降低而逐渐上升。郭黄萍等[3]在山西梨部分栽培品种抗寒性研究初报中得出,梨枝条中游离脯氨酸含量随着外界温度的降低而升高。黄浩等[4]在黄梁木种源的抗寒性综合评价中得出,在低温胁迫条件下,不同黄梁木种源的枝芽存活率与游离脯氨酸含量呈极显著正相关,说明生理指标的变化与枝芽的抗寒性密切相关。
丙二醛含量在抗性生理研究中是一种常用的指标,它能间接地反映植物对低温胁迫的抵抗和忍耐能力,可以利用其作为脂质过氧化指标来表示植物本身对逆境条件的反应[5]。王庆军等[6]在3个石榴品种的抗寒性分析中指出,植物的抗寒性与丙二醛含量呈负相关关系。赵欢蕊等[7]在对辽东栎在低温胁迫下的生理响应研究中指出,丙二醛含量随着自然温度的降低逐渐上升。何威等[8]在山茶花耐寒性研究中也指出,丙二醛含量在自然降温过程中随着温度的降低逐渐增加,其可作为鉴定山茶花品种耐寒性强弱的生理指标。
过氧化物酶(POD)活性的变化在一定程度上可以反映植物的抗寒性,当植物感受到外界温度降低时,过氧化物酶通过酶促降解H2O2,避免自由基对细胞膜的伤害,保护植物细胞[9]。王勇等[10]在核桃优良品种枝条抗寒性生理生化指标分析中指出,随着核桃生育进程的继续以及气温的变化,POD酶活性在核桃枝条内含量越高,抗寒性越强。艾鹏飞等[11]在研究仁用杏抗寒性生理指标评价中得出,过氧化物酶活性在胁迫降温过程中呈现出上升趋势,且与仁用杏枝条的萌芽率相关性较好。因此,过氧化物酶活性的变化在一定程度上可以反映植物的抗寒性。
合欢和紫荆是榆林市近年来引进的新树种,在已有的研究中,关于合欢和紫荆对低温的反应,尤其是抗寒生理的研究较少。榆林地处陕西省最北部,冬季寒冷,而这2种植物皆喜温暖湿润气候,低温是限制其引种的主要环境因子[12]。
本试验在低温条件下对合欢和紫荆的3种抗寒性生理指标进行比较,旨在探索其在自然降温过程中生理变化的规律,为进一步扩大其引种范围提供理论支持。
1 材料和方法
1.1 试验地概况
试验地位于陕西省榆林市榆林学院校园内,位于东经 108°58′~110°24′,北纬 37°49′~38°58′。属于温带半干旱大陆季风气候,一年中以12月至次年2月日照差异最大,四季温差明显,干湿不同。年平均气温为8.3℃,冬季寒冷,平均气温在-7.8~4.1℃,极端最低温曾达到-32.7℃。试验期间(2016年11月至2017年3月)榆林地区气温变化情况如图1所示。
1.2 试验材料
以合欢和紫荆的一年生休眠枝条为试验材料。
1.3 试验方法
从2016年11月1日至2017年3月1日,每隔10 d左右取样一次,取样时间分别为11月1日、11月11日、11月21日、12月1日、12月11日、12月21日、1月1日、1月11日、1月21日、2月1日、2月11日、2月21日、3月1日,共13次,采集植物时取上部生长良好的一年生休眠枝条,放入保鲜袋中带回实验室作为试验材料备用。
1.4 测定项目及方法
游离脯氨酸含量的测定采用茚三酮法[6],丙二醛含量的测定采用巴比妥酸法[8],POD活性的测定采用愈创木酚法[6]。
1.5 数据分析
采用SPSS 22.0软件进行数据分析,采用Excel 2010进行图表处理。
2 结果与分析
2.1 自然降温对合欢和紫荆游离脯氨酸含量的影响
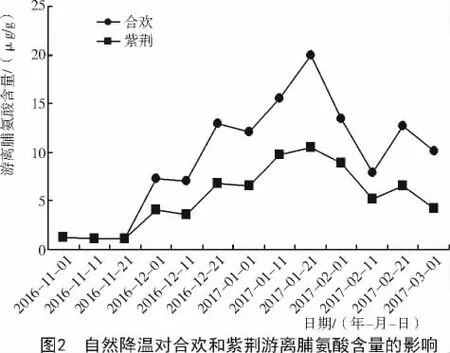
由图2可知,在试验期间,合欢枝条中游离脯氨酸含量随着温度的降低而升高,随着温度的升高而降低,与温度呈负相关。当温度在0℃以上时(2016-11-01—11-21),随着温度的升高,游离脯氨酸含量缓慢减少;在12月1日当温度降至0℃以下时,再随着温度的升降,游离脯氨酸含量呈波形变化,共出现4个波峰,分别出现在温度骤降的12月1日(-8℃),12月21日(-8℃),1月21日(-20℃)和2月21日(-8℃),各波峰对应的游离脯氨酸含量分别为 7.23,12.99,19.99,12.64 μg/g。整体上,游离脯氨酸含量呈先上升后下降的趋势。
紫荆植物体内脯氨酸含量的变化,与合欢基本相同。当温度在0℃以上升温时,脯氨酸含量缓慢减少,在12月1日温度降至0℃以下时,随着温度的变化,脯氨酸含量呈波形变化,共出现4个波峰,各波峰对应的脯氨酸含量分别为4.01,6.83,10.41,6.57 μg/g。与合欢相比,均有所下降,分别下降了3.22,6.16,9.58,6.07 μg/g。此后,随着温度的上升,至试验末的3月1日(-5℃),合欢脯氨酸含量下降至 10.03 μg/g,紫荆下降至 4.12 μg/g。在同一时期内,合欢枝条中的脯氨酸含量均高于紫荆。
2.2 自然降温对合欢和紫荆丙二醛含量的影响
由图3可知,在试验期间,合欢枝条中丙二醛含量随着温度的降低而升高,随着温度的升高而降低,与温度呈负相关。当温度在0℃以上时(2016-11-01—11-21),随着温度的升高,丙二醛含量缓慢减少;在12月1日当温度降至0℃以下时,随着温度的升降,脯氨酸含量呈波形变化,共出现4个波峰,分别出现在温度骤降的12月1日(-8℃),12月21日(-8℃),1月21日(-20℃)和2月21日(-8℃),各波峰对应的丙二醛含量分别为8.02,9.06,13.46,7.83 μmol/g。整体上,丙二醛含量呈现出先上升后下降的趋势。
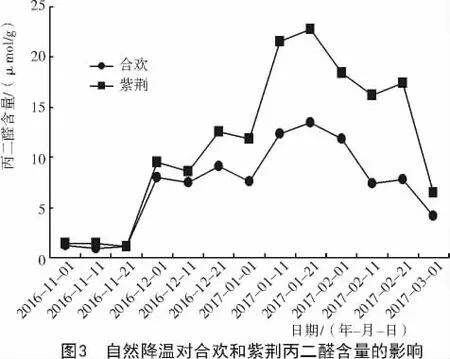
紫荆植物体内丙二醛含量的变化,与合欢基本相同。在试验初期温度在0℃以上时,丙二醛含量变化较小,当12月1日温度降至0℃以下时,随着温度的升降,丙二醛含量呈波形变化,共出现4个波峰,各波峰对应的丙二醛含量分别为9.56,12.53,22.73,17.38 μmol/g,与合欢相比,均有所上升,分别上升了 1.54,3.47,9.27,9.55 μmol/g。此后,随着温度的上升,至试验末的3月1日(-5℃),合欢丙二醛酸含量下降至4.11 μmol/g,紫荆下降至6.53 μmol/g。在同一时期内,合欢枝条中的丙二醛含量均比紫荆低。
2.3 自然降温对合欢和紫荆过氧化物酶(POD)活性的影响
由图4可知,在试验期间,合欢枝条中过氧化物酶活性随着温度的降低而升高,随着温度的升高而降低,与温度呈负相关。当温度在0℃以上时(2016-11-01—11-21),随着温度的逐渐升高,过氧化物酶活性缓慢降低;在12月1日当温度降至0℃以下时,随着温度的变化,过氧化物酶活性呈波形变化,共出现4个波峰,分别出现在温度骤降的12月1日(-8℃),12月21日(-8℃),1月21日(-20℃)和2月21日(-8℃),各波峰对应的POD活性分别为 11.19,15.89,30.01,15.26 U/(g·min)。整体上,POD活性呈现出先升高后降低的趋势。
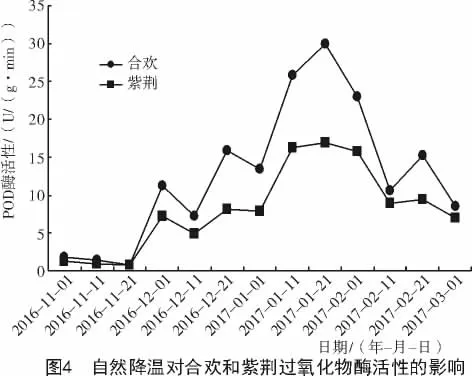
紫荆植物体内POD活性的变化,与合欢基本相同。当温度在0℃以上升温时,POD活性缓慢降低,当12月1日温度降至0℃以下时,随着温度的变化,POD活性呈现出波形变化,共出现4个波峰,各波峰对应的POD活性分别为7.29,8.12,16.88,9.37 U/(g·min)。与合欢相比,均有所降低,分别降低了3.9,7.77,13.13,5.89U/(g·min)。此后,随着温度的上升,至试验末的3月1日(-5℃),合欢POD活性下降至8.59 U/(g·min),紫荆下降至7.02 U/(g·min)。在同一时期内,合欢枝条中的POD活性均高于紫荆。
3 讨论
本试验表明,在自然降温过程中,合欢和紫荆中的脯氨酸含量、丙二醛含量、过氧化物酶活性均呈波形变化,随着温度的降低而增加,随着温度的升高而减少,与温度呈负相关关系。这分别与李丽等[13]对淡竹、张艳侠等[14]对石榴、鲁金星等[15]对砧木和酿酒葡萄枝条中的研究结果一致。可能是因为植物在低温条件下,会分泌植物蛋白,以增加细胞液的浓度来维持植物的生命[16],如脯氨酸能够维持细胞结构、细胞运输和调节渗透压等,使植物具有一定的抗性和保护作用[17]。而当植物在低温刺激后产生POD对H2O2有促降解的作用,得以提高植物的抗寒性,同时温度的降低使植物细胞过氧化,从而产生大量的丙二醛。
王小媚等[18]研究表明,在低温胁迫过程中,杨桃的抗寒性指标POD活性呈现出先升后降的变化趋势,呈上升趋势可能是因为温度过低,超出了植物的耐寒范围。而本试验中,POD活性与温度呈负相关关系,可能是因为自然降温过程中出现的最低温度还没有达到2种树种所能忍受的临界低温,说明POD活性在不同的作物中抗寒性存在一定的差别,这与石如意等[19]、王勇等[20]的研究结果一致。
4 结论
本试验通过对游离脯氨酸含量、丙二醛含量、过氧化物酶活性3个指标的测定来进行合欢和紫荆2种植物的抗寒性比较分析。综合3种指标得出,合欢的抗寒性大于紫荆。这一结论对于扩大植物引种、丰富榆林本地的园林绿地生物多样性指数、进一步改善生态坏境具有重大意义。此外,由于植物在抵御低温的过程中,会产生一系列错综复杂的生理变化,且易受多种外界因素的影响,考虑今后可进一步通过对半致死温度、电阻率、可溶性蛋白等生理指标进行综合评定,从而加强对植物抗寒性鉴定的准确性和可靠性。
