16个枸杞品种(系)S-RNase基因型鉴定及序列分析
2018-12-11戴国礼张兆萍焦恩宁李彦龙
戴国礼,张兆萍,程 慧,4,张 波,焦恩宁,李彦龙,秦 垦*
(1.宁夏农林科学院枸杞工程技术研究所,宁夏银川 750001;2.国家枸杞工程技术研究中心,宁夏银川 750001;3.甘肃省农业工程技术研究院,甘肃武威 733006;4.西南林业大学林学院,云南昆明 650224)
【研究意义】自交不亲和性是植物预防近亲繁殖和保持遗传变异的一种重要机制,在被子植物中普遍存在[1]。根据植物自交不亲和的遗传控制机理,将植物的自交不亲和性分为孢子体型SSI(Sporophytic self-compatibility,简称 SSI)、和配子体型GSI(Gametophytic self-incompatibility,简称 GSI)[2]。其中,以茄科、蔷薇科和玄参科为代表的配子体自交不亲和性是最常见的类型[3]。【前人研究进展】多年研究发现,GSI植物的自交不亲和性由单一的多态性S-位点控制,该位点为多基因复合体,至少包括2个自交不亲和反应特异性决定因子:花柱S基因与花粉S基因[4]。研究证实,蔷薇科、茄科及玄参科等GSI类植物具有共同的花柱S基因产物,为一种糖蛋白,且发现其具有核糖核酸酶活性,故称为S 核酸酶(S-RNase)[5-6]。S-RNase 是雌蕊-花粉相互识别的关键物质,可分解花粉管RNA,抑制花粉管的伸长,从而导致自交不亲和[7]。继1981年首次从花烟草花柱提取物中分离出与自交不亲和相关的S 糖蛋白,1986 年克隆了 S-cDNA 之后[5-6],已有大量的S-RNase序列得到了克隆。已明确花柱的SRNase在雌蕊识别和拒绝花粉的过程中起着重要作用[8-9]。【本研究切入点】宁夏枸杞(Lycium barbarum L.)在中国分布广泛,为茄科(Solanacease)枸杞属(Lycium L.)落叶灌木,属配子体自交不亲和植物。有学者在田间对宁夏枸杞进行了大量的授粉杂交试验,结果表明,在宁夏枸杞种内存在自交不亲和现象,其繁育系统除麻叶系外都属于专性异交,大多品种具有自交不亲和性,在生产上需配置授粉树[10-11]。育种家已培育出了一定数量的栽培品种与新优系,但目前对各枸杞品种(系)的自交不亲和性及相关基因还缺乏系统深入的研究,使得杂交亲本选配与配置授粉树时具有很大的盲目性,且费时费工。【拟解决的关键问题】本研究以16个国内枸杞主栽品种(系)和地方品种为试材,应用PCR、回收、克隆、测序技术,开展枸杞品种(系)S-RNase基因的分子生物学研究,鉴定参试材料的S-RNase基因型,同时了解宁夏枸杞品种自交不亲和S-RNase基因的多态性。同时,通过田间套袋试验方法,测定亲和指数进行验证。最终确定各品种的自交亲和性及其所含有的S-RNase基因类型,以期为枸杞栽培合理配置授粉树和杂交亲本选配提供理论依据。
1 材料与方法
1.1 材料
供试的16份枸杞品种(系)种植于宁夏农林科学院国家枸杞工程技术研究中心枸杞种质资源圃(38°38'49″N,106°09'10″E),于 2016 年春季采集各品种(系)的嫩叶置于-80℃保存(表1)。
1.2 方法
1.2.1 人工授粉与套袋试验 按照秦 垦等对枸杞属植物自交亲和性的检测和界定方法[10],对参试材料花蕾进行以下2种处理:①同株同花人工授粉后套袋,检测单花自交是否亲和;②同株异花人工授粉后套袋,检测株内自交是否亲和[12]。同一品种(系)每个处理选30个花蕾进行试验,每个实验进行3次重复,35 d后收获果实,统计每个果实种子含量,进而计算自交亲和指数。以自交亲和指数高低判断自交亲和性强弱,亲和类型的划分主要参照方智远、刘后利等的方法进行,自交亲和指数小于5的为自交不亲和,自交亲和指数大于5的为自交亲和。计算见公式(1)。
自交亲和指数=自交结实种子总数/套袋自交花蕾数 (1)
1.2.2 DNA提取及S-RNase基因PCR扩增 采用CTAB法提取各品种(系)基因组DNA。根据茄科S基因一级结构保守区序列合成PCR扩增兼并引物。由上海捷瑞生物工程有限公司合成。引物序列信息为:正向 si200:5’-TCYGTWATGMTKAATAACTGC-3’;反向 si552:5’-AAYRCACTTKAGRCTWGGAAC C-3’(图1)。
摸索扩增条件,建立特异性PCR扩增体系。PCR体系为:依次加入200 ng模板DNA,5 μl 10×Taq Buffer,2.5 nmol dNTPs,正反向引物分别 0.008 nmol,1 U TaqDNA 聚合酶,补充灭菌水至 25 μl,轻弹混匀后加入矿物油,短暂离心。扩增程序:94℃预变性3 min;94℃变性30 s,55℃退火30 s,72℃延伸45 s,35个循环,再72℃延伸10 min;4℃保存。

表1 材料名称及来源Table 1 A list of material name and source

图1 茄科S基因一级结构Fig.1 The basic structure of Solanaceae S-gene
1.2.3 S-RNase基因扩增产物回收、克隆及测序PCR产物经1.2%琼脂糖凝胶电泳检测目的条带的有无。按照AxyPrep DNA凝胶回收试剂盒回收目的片段,将回收产物连接到PMD18-T载体上,转化至DH5α感受态细胞,每个材料挑15~20个阳性单克隆送去测序,确保每个S基因型至少重复测定2次。序列委托上海英潍捷基公司测定。
1.2.4 序列分析与比对 借助EditSeq软件对测序结果进行预处理,在NCBI中利用Blastn检索Gen-Bank中已登陆的S-RNase基因同源序列。用DNAMAN进行多序列比对分析,并推导对应的氨基酸,确定测序结果的S等位基因名称。
2 结果与分析
2.1 S-RNase基因特异性扩增
用1.2%的琼脂糖凝胶电泳检测各供试材料的PCR产物。由图2可知,各供试材料均在约500 bp处有一条明显的亮带。
2.2 枸杞品种(系)S-RNase基因型鉴定

图2 枸杞品种的S基因特异性扩增Fig.2 The specific amplification of S-RNase fragments from Lycium barbarum L.cultivars
分析测序结果,最终获得除去两端载体及引物后长为420 bp的目的片段,分别在NCBI中进行Blast,结合同源比对与序列分析结果,确定均为枸杞属的S基因(图3)。DNA多序列比对结果显示:这17个未知S-RNase基因型的参试材料共涉及9种SRNase基因片段,分别标记为 A、B、C、D、E、F、G、H和I。同时发现,这9种S-RNase基因片段在2个高变区内均具有很高的序列多态性。除‘宁农杞1号’外,其余供试材料均含有其中的2种。依据植物内含子(5GT-3AG)的序列特点,同时参考已登陆的茄科S-RNase基因一级结构特点,确认了位于高变区HVa内一段长为109 bp的内含子序列,进而推导出高变区及其周围核苷酸对应的氨基酸序列(图4)。
将这9条序列分别进行Blastn发现,除G和I两种序列外,其余7种核苷酸序列与GenBank中已登录的枸杞S-RNase等位基因部分编码区序列相似性均在99%以上,分别为:A序列与S-BARB2(GI:323320147)、B序列与S-BARB8(GI:323320159);C序列与S-BARB3(GI:323320149);D序列与SBARB6(GI:323320155);E序列与S-CHIN3(GI:323320165);F序列与S-BARB9(GI:323320161);H序列与S-CHIN6(GI:323320169)。由以上7种类型的S-RNase基因核苷酸序列推导的氨基酸和对应同源性最高的已登录S-RNase等位基因推导的氨基酸序列也完全一致,依据S-RNase基因相同时其DNA序列同源性应为100%的原理,认定为同一种基因。
2.3 2个新S-RNase基因型的序列分析
同源性分析结果显示,G序列与已登录S等位基因 S-BARB6(GI:323320155)一致性最高,达98%,推导的氨基酸序列一致性达95%。分析其氨基酸序列特征,在高变区HVa有2个氨基酸发生变化:由T→A的突变,导致 M(Methionine)→K(Lysine)的变化;由C→G的突变,导致P(Proline)→A(Alanine)的变化。在高变区HVb有1个氨基酸发生变化:由G→C的突变,导致K(Lysine)→N(Asparagine)的变化。在保守区C3、C4之间有2个氨基酸发生变化。分析比较已登录的枸杞属S等位基因,发现S基因的多态性主要是因为高变区(H V)的变异,通常根据高变区的不同来确认不同的S等位基因,由此认定G序列为一个新的S-RNase等位基因,暂将其命名为S-BARB6n。

图3 S-RNase基因序列同源性比较Fig.3 Alignment of nucleotide sequences of S-RNase

图4 9种片段推导的氨基酸序列Fig.4 Putative amino acid sequences of nine fragments

图5 G序列与S-BARB6等位基因DNA序列比对Fig.5 The result of DNA sequences alignment between G and S-BARB6 allele

图6 G序列与S-BARB6等位基因的氨基酸序列比对Fig.6 The result of amino acid sequences alignment between G and S-BARB6 allele
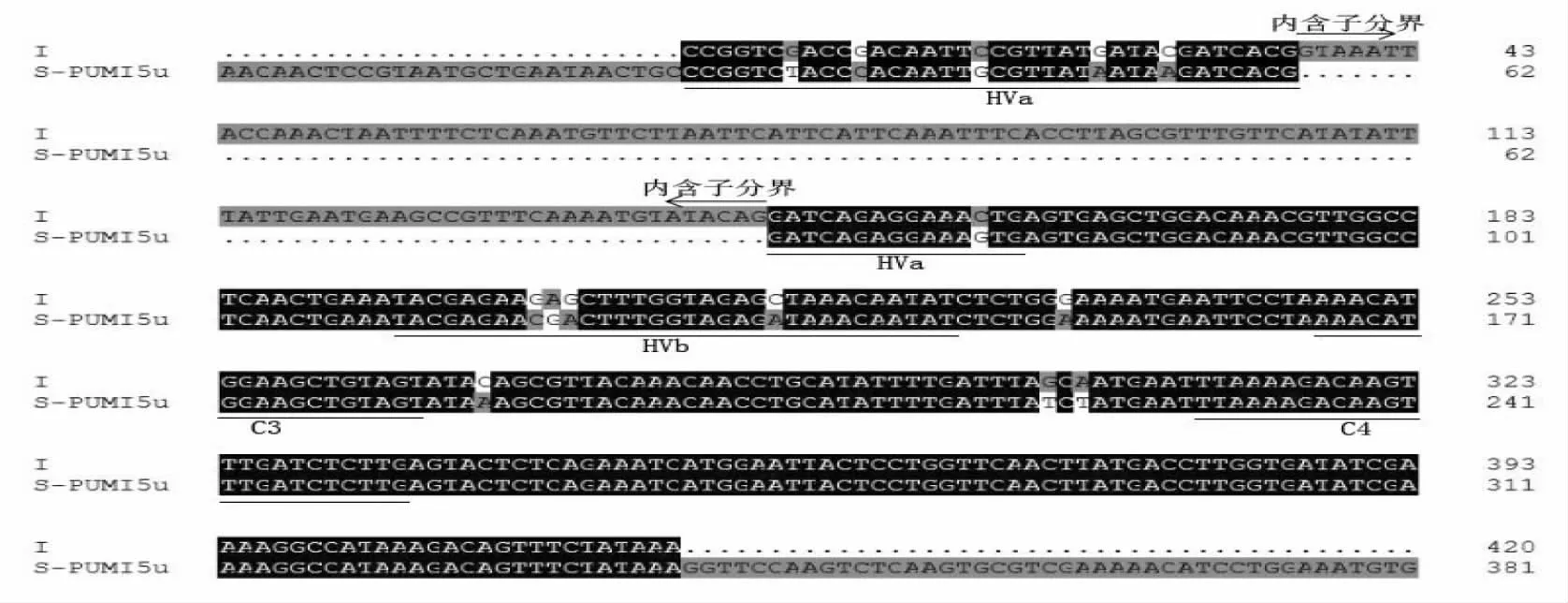
图7 I序列与S-PUMI5u等位基因DNA序列比对Fig.7 The result of DNA sequences alignment between I and S-PUMI5u allele

图8 I序列与S-PUMI5u等位基因的氨基酸序列比对Fig.8 The result of amino acid sequences alignment between I and S-PUMI5u allele
I序列与已登录S等位基因S-PUMI5u(GI:323320217)一致性最高,达96%,推导的氨基酸序列一致性达89%,分析比较氨基酸序列特征,在高变区HVa有6个氨基酸发生变化:由G→T的突变,导致D(Aspartic acid)→Y(Tyrosine)的变化;由G→C的突变,导致R(Arginine)→P(Proline)的变化;由C→G的突变,导致F(Phenylalanine)→L(Leucine)的变化;由G→A的突变,导致D(Aspartic acid)→N(Asparagine)的变化;由C→A的突变,导致T(Threonine)→K(Lysine)的变化;由C→G的突变,导致L(Leucine)→V(Valine)的变化。在高变区HVb有3个氨基酸发生变化:由GAG→CGA的突变,导致K(Lysine)→N(Asparagine)的变化;由C→A的突变,导致S(Serine)→D(Aspartic acid);由C→A的突变,导致A(Alanine)→D(Aspartic acid)。在保守区C3、C4之间有2个氨基酸发生变化。认定I序列也是一个新的S-RNase等位基因,暂命名为S-PUMI5un。
2.4 参试材料自交(不)亲和性的田间验证
通过田间授粉实验,可以发现参试的16个品种(系),有11个属于自交不亲和材料,和同株自花交和同株异花交自交亲和指数小于5的为自交不亲和;5个属于自交亲和材料,其同株自花交自交和同株异花交自交亲和指数大于5的为自交亲。
其中自交亲和品种(系)的基因型为:‘宁杞1号’、‘宁 农杞 3 号’、‘03-08’、(S-BARB2/SBARB8),‘宁杞7号’(S-BARB8/S-BARB6)、‘山东黄果枸杞’(S-BARB2/S-PUMI5un);其中自交不亲和品种(系)的基因型为:‘中国枸杞’(S-BARB2/SCHIN6),‘云南枸杞’(S-BARB3/S-CHIN3),‘黄果枸杞’(S-BARB3/S-BARB6n),‘宁杞 5号’(SBARB2/S-CHIN3),‘宁农杞 1号’(S-BARB2/SBARB2),‘宁农杞 2号’(S-BARB2/S-BARB6n),‘宁农杞9号’(S-BARB2/S-BARB9),‘07-05’(SBARB8/S-BARB6n),‘06-16’(S-BARB2/S-CHIN3),‘07-08’(S-BARB8/S-BARB6n),‘13-01’(S-BARB 2/S-CHIN3)。
3 讨论
3.1 枸杞SI与SC之间S-RNase基因型比较
本研究对11个自交不亲和5个自交亲和品种(系)的 S-RNase基因型进行比较分析发现:SBARB2、S-BARB8等位基因具有很高的基因频率,氨基酸多序列比对结果显示,S-BARB8等位基因在C4保守区后的外显子位置与其他几种等位基因相比有2个位点的氨基酸同时发生变化,即Q/D→L和D/G→H,由此引起的S-RNase基因表达产物的变化,有可能是枸杞自交不亲和向自交亲和突变发生的根本原因。枸杞自交不亲和S-RNase基因的系统进化演变,还需进一步研究。
3.2 枸杞S-RNase的氨基酸序列的特异性
枸杞属(Lycium L.)从经典分类学上属于为茄科(solanacease),茄科中S-RNase有5个保守区域(C1 到 C5)和 2 个高变异区(Hva 和 HVb)[16-17]。并且在茄科的S-RNase中存在的潜在N-糖基化保守位点在C2区,对比其他配子体自交不亲和植物,如蔷薇科的S-RNase的C2区却没有包含N-糖基化位点,同时Ushijima[18]等的研究中,提出蔷薇科的 SRNase不仅有 C1、C2、C3、C4,4 个保守区,而且还有一个RC4区,RC4是蔷薇科所特有的专一性区域,在其他T2/S型的S-RNase中没有发现。并且指出高度变异区RHV定位在与茄科S-RNase的高变异区相似的位置,茄科S-RNase的另一个变异区HVb在蔷薇科的S-RNase中却是保守的[14]。本研究所克隆的枸杞S-RNase基因片段,大小为420 bp,包含了C2、C3、C4保守区,和 2个高变异区(Hva和HVb),并对不同品种(系)的S-RNase基因型进行了鉴定,但是没有得到全长序列,对其结构和特异识别位点的分析存在难度,这也是本研究的不足之处。
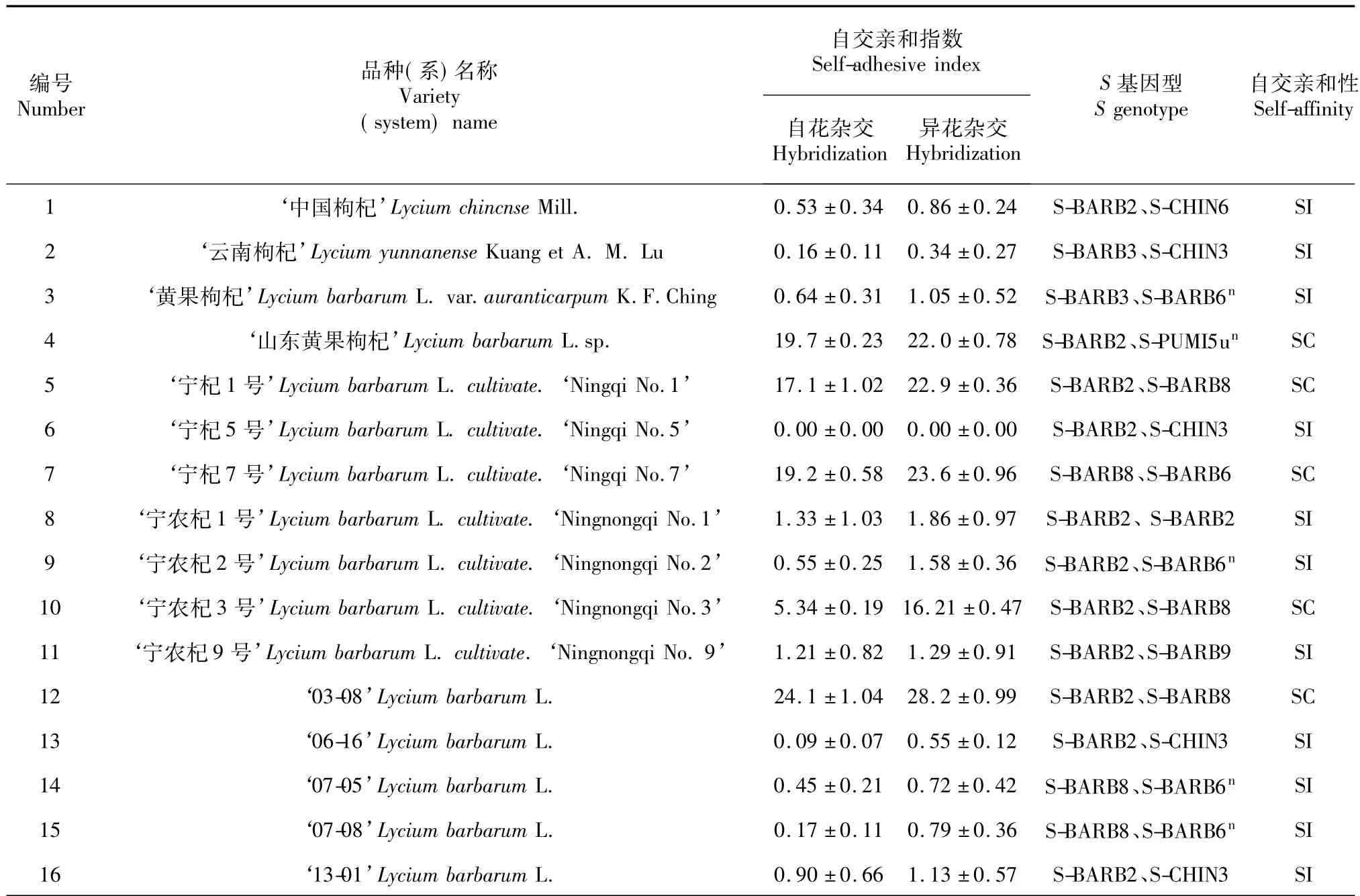
表2 16个枸杞品种(系)的自交亲和性及其S-RNase基因型Table 2 The self-compatibility and S-RNase genotypes of seventeen Lycium L.cultivars
3.3 枸杞S-RNase基因型鉴定的意义
已证实,品种间的亲和性和S-RNase基因型间存在密切联系,若2个品种的S-RNase基因型相同,则相互授粉不亲和,否则表现为亲和。对于鉴定出的11份自交不亲和枸杞品种(系),生产中在配置授粉品种或杂交育种在选配亲本时,品种间的亲和性是首要考虑因素,而本研究S-RNase基因型鉴定结果即可提供有利依据。
本研究结果不仅丰富了枸杞属植物的自交不亲和基因信息,同时能为生产中授粉树的配置,杂交育种亲本选配提供理论依据。继续深入研究枸杞花柱、花粉S-RNase基因及表达产物功能与相互作用机制是未来枸杞自交不亲和性研究领域重要的发展方向。
4 结论
本研究以16个品种(系)为材料,共克隆9个S等位基因,分别为 S-BARB2、S-BARB3、S-BARB6、SBARB8、S-BARB9、S-CHIN3、S-CHIN6,并发现 2 个新的S等位基因,暂分别命名为 S-BARB6n,S-PUMI5un。所克隆获得的9个枸杞S-RNase基因均包含了C2、C3、C4保守区,和2个高变异区(Hva和HVb)。通过田间验证试验证明,11个品种(系)为自交不亲和材料,5个为自交亲和材料。枸杞属植物存在S-RNase等位基因,其基因序列与茄科植物具有一定相似性,枸杞S-RNase基因是控制枸杞自交不亲和性的关键基因,可根据本实验结果进行授粉树的配置和杂交育种亲本选配。
