大肠杆菌ABC转运体研究进展
2018-12-08,,
,,
ABC转运体是所有转运体家族中最大的一类转运体,广泛分布于从原核生物到人类的几乎所有的物种中。大部分的ABC家族转运体都是跨膜蛋白质,通过偶联ATP的水解释放的能量转移底物。这些蛋白质组成了古老的转运体家族,其进化时间超过30亿年。系统发生方面的证据支持ABC家族的分歧时间早于细菌、古细菌和真核生物的分歧时间,许多人类的ABC转运体与某些细菌如大肠杆菌的ABC同源。
不同类型的ABC转运体转运不同的底物,从小分子如离子、糖或氨基酸到大分子如抗体、药物、磷脂和寡肽等。在细菌中ABC转运体参与营养物质的摄取或细菌毒素的分泌以及将抗生素泵出细胞使细菌具有抗药性;在植物中,ABC家族转运体参与对除草剂的抗性[1];在人类中,ABC家族基因突变会导致功能改变,与一些遗传病如囊性纤维化有关,另外ABC家族转运体的上调表达与肿瘤细胞的耐药性有关。ABC家族的蛋白质不仅参与物质的跨膜运输,也参与除转运以外的其它功能如mRNA转录及DNA损伤修复等[2-3]。
1 大肠杆菌的ABC转运体
大肠杆菌是革兰氏阴性菌,是包括人类在内许多动物的肠道正常菌群。大肠杆菌K-12菌株,是一个非毒力缺乏O-抗原和K-抗原并在实验室广泛使用的菌株,常被认为是大肠杆菌培养的标准菌株。1997年大肠杆菌K-12菌株完成全基因组测序,在其基因组中最大的蛋白质家族是ABC转运体家族,发现了至少72个编码ABC家族蛋白的基因,占整个基因组的5%[3]。这个庞大的转运体对于维持大肠杆菌正常的营养吸收及代谢产物的排出是非常重要的。而在人类基因组中只有48个ABC转运体,金黄色葡萄球菌、果蝇、拟南芥分别有28、56、113个ABC转运体系统。除植物外,真核生物较原核生物含有较少的ABC系统。如果基因组中ABC系统数量多,说明这个生物有更多的底物被运进或运出。因此在真核生物中,能运动的生物体如动物较不能运动的生物如植物需要较少的ABC转运体。因为能运动的生物含有较丰富的主动运输系统保证营养物质的供给和代谢产物的排出。
2 大肠杆菌ABC转运体分类
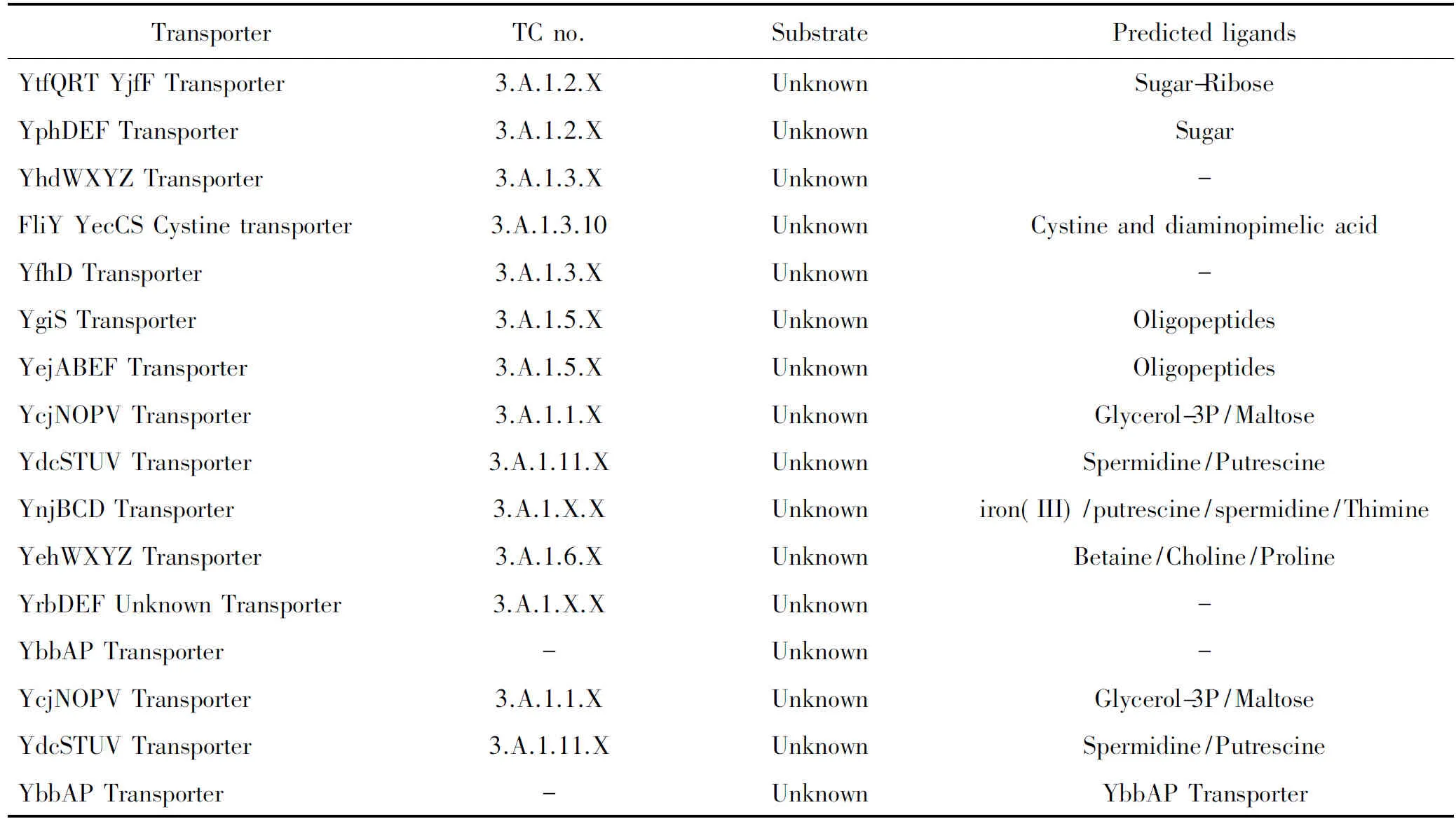
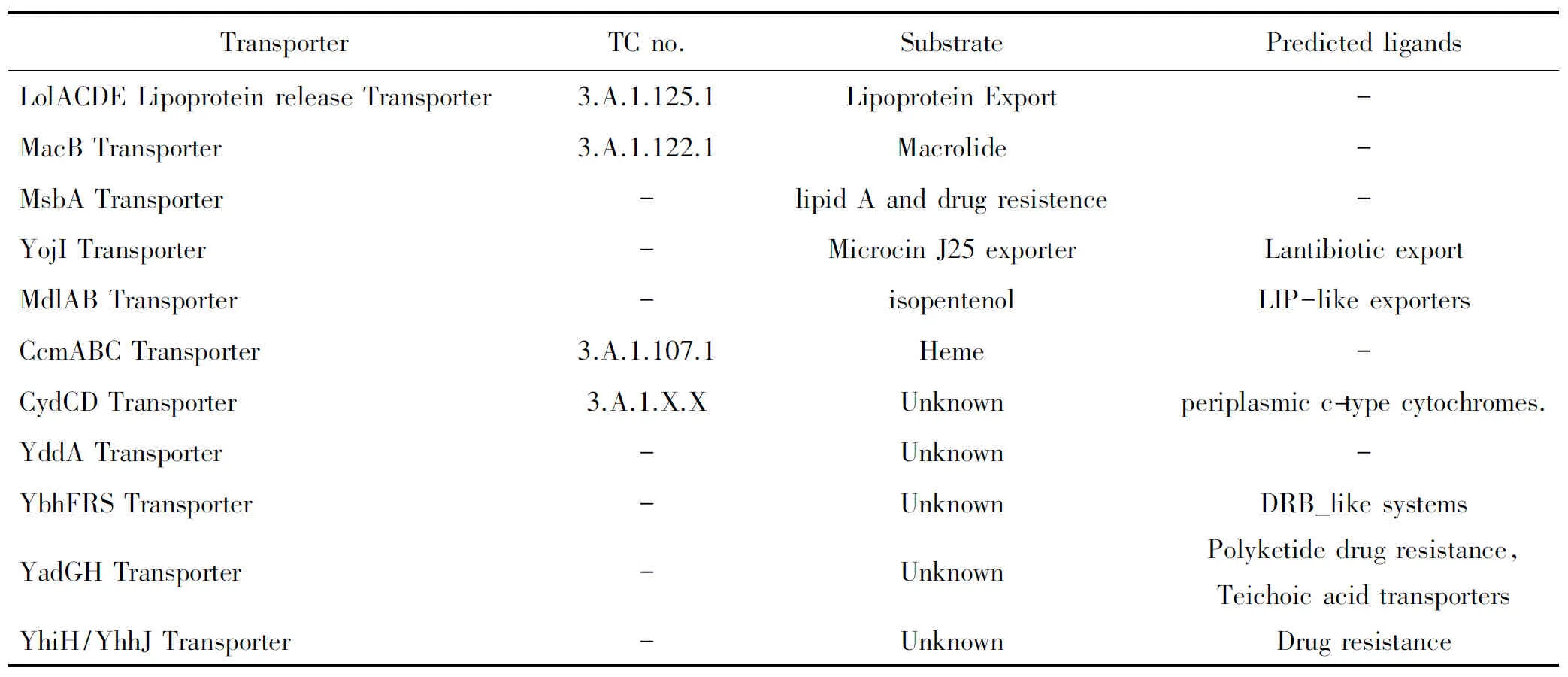
目前大肠杆菌ABC超家族根据研究进展及功能分为3类。第一类为摄取系统,该类转运体将胞外的物质运输到细胞内,是大肠杆菌获取营养的主要途径,该类转运体包括已经被实验证实的摄取系统共41个(见表1),其中研究比较清楚的包括大肠杆菌VB12摄取系统BtuCDF转运体,HisJMPQ组氨酸转运体,OppABCDF寡肽转运体等,还有16个预测的ABC摄取系统,即这16个摄取系统目前为止功能还不清楚(见表2)[4]。第二类为输出系统,这类转运体的底物主要包括细胞内合成的需要或能够运输到胞外执行功能的蛋白质,例如胞外的外周蛋白,细胞壁的主要成分磷脂及脂多糖等都是依赖ABC转运体外排到胞外的,外排的底物还包括重金属、大肠杆菌产生的溶血素、小菌肽、大肠杆菌素及抗生素等,大肠杆菌的外排转运体有11个(见表3)[3],其中有一部分是根据结构域预测的。这些ABC外排泵对于大肠杆菌适应不良环境起非常重要作用。大肠杆菌外排系统研究较为清楚的包括LolCED负责脂蛋白在内外膜间的翻转[5];MsbA在相关功能研究之前预测为与细菌抗药性有关的外排系统,但是经功能研究之后发现该系统不但与多药抗性有关,还负责将细胞质合成的类脂A(lipid A)转运到膜周间隙中[6];脂多糖的外排系统LptAB[7];溶血素外排系统HlyAB[8];大肠杆菌素外排系统CvaAB[9];亚铁血红素外排系统CcmAB[10];YojI 转运体负责小抗菌肽J25的外排[11];以及与大环内酯类药物外排有关的macAB等[12]。输出系统的大部分成员功能还未知;第3类为与物质运输无关的系统(见表4)。参与mRNA转录、DNA损伤的修复等,因此有人将ABC超家族翻译成ABC系统。因为并不是所有的ABC家族成员都参与一定底物的转运。大肠杆菌的非转运体预测的有4个,这类转运体典型的代表为UvrA,是目前为止已知的唯一一个与转运没有关系的ABC家族成员。UvrA系统在紫外线诱导的DNA损伤中起到修复DNA的作用[13]。
3 大肠杆菌ABC转运体的结构
所有的ABC转运体无论是摄取或输出转运体都有相似的结构,即包含整合膜蛋白构成的2个跨膜结构域(MD)和2个核苷酸结合结构域(NBD),在许多细菌中,这4个结构域分别由不同的基因编码,如大肠杆菌的麦芽糖转运体复合物MalEFGK[14]和核糖转运体复合物RbsBCD[15],而在有些细菌和真核生物中这4个结构域可能由一个基因编码,同时包含1个NBD结构域和1个MD结构域,2个这样的蛋白形成二聚体发挥功能,这样的ABC转运体称为半转运体如大肠杆菌的磷脂A和多药外排转运体MsbA[7],也有些ABC家族转运体由1个基因编码同时含有2个NBD结构域和2个MD结构域,这样的转运体称为全转运体,如人类的肿瘤抗药性转运体P-型糖蛋白就是1个典型的ABC家族的全转运体[16]。在过去的10年里来自于不同细菌的45个不同的ABC转运系统有91个ABC晶体结构被报道,其中35个是SBPs结构,另外56个晶体结构是NBD结构和全长的转运体结构。后56个包含5个来自于4个不同的转运体和51个NBD结构域,其中NBD结构域中13个是以二聚体形式存在,另外的38个都是单体[17-27]。
3.1核苷酸结合结构域 ABC转运体的NBD结构域保守性较强,在几乎所有的ATP结合蛋白中都包括保守的Walker A和Walker B结构域,而信号结构域(LSGGQ)只出现在ABC转运体家族中。NBD结构域是ABC转运体的马达结构域,能结合并水解ATP产生的能量诱导NBD构象改变,迫使NBD结构域相互靠近并与ATP形成三明治结构,这些改变传递给MD结构域,引起其构象改变,使其向内或向外开放,从而使底物输入到胞外或胞内。
基于已经获得的晶体结构,一般的NBD结构域包含3个部分:α螺旋结构域,1个催化结构域和1个调节结构域。而α螺旋结构域和催化结构域在所有的NBD结构域中都存在,只有在CysA,GlcV和MalK转运体中存在额外的调节结构域。所有的ABC转运体的NBD结构域包含高度保守的结构域如Walker A (GXXGXGKS/T with X 代表如何氨基酸) 和 Walker B (ΦΦΦΦD with Φ 代表输水氨基酸残基),这2个motif包含了ATP识别和结合的位点[28],而信号结构域(signature motif LSGGQ,也叫做C-loop或LSGGQ motif)只出现在ABC转运体中是ABC转运体的标志性结构域,在NBD形成的二聚体中顺向NBD的Walker A motif与反向NBD的signature motif 与ATP的□-磷酸结合形成催化结构域。

表1 大肠杆菌中已经被证实的输入系统(41个)Tab.1 Experimentally confirmed import system

NBD结构域还包含A-环(A-loop),Q-环(Q-loop),D-环(D-loop)和H-环(H-loop也叫做switch loop)仅由一个保守的氨基酸残基组成。组成A-环的氨基酸残基为芳香族的氨基酸残基,通常是酪氨酸,对于ATP与NBD结构域的结合起辅助作用[29-30];组成Q-环的氨基酸残基为谷氨酰胺,与ATP’s的γ-磷酸结合;组成Q-环的氨基酸残基为组氨酸,与ATP’s的γ-磷酸直接结合。在全长的ABC转运体中Q-环即与MD结构域结合又与ATP结合,在NBD和MD结构域的构象偶联中起重要作用[26,31]。
ATP与一个核苷酸结合位点水解引起NBD的二聚化,但是2个位置的水解需要ATP与NBD形成三明治样结构[32]。ATP的结合、水解及ADP的释放诱导NBD结构域构象的变化,NBD构象的变化诱导MD构象也发生变化从而完成底物的转运。
3.2ABC转运体的跨膜结构域 与保守的NBD结构域相比,跨膜结构域(MD)的保守性较差[33],这与MD结构域参与不同底物转移的功能有关。在ABC外排转运体中,全长的转运体含有2个MD结构域形成1个二聚体结构,包含12个跨膜的α螺旋,而ABC摄取转运体中,跨膜α螺旋数量不同,10~12个跨膜螺旋可能是底物跨膜转移的最小数量。许多ABC外排泵的MDs都有2个位于细胞外的环和2个位于细胞质基质的环。在金黄色葡萄球菌多药外排转运体Sav1866 同源二聚体结构中,位于细胞质基质内有2个短偶联螺旋形成的环(ICL1和ICL2)与细胞膜平行并与NBD结构域相互作用[34]。第一个偶联螺旋(ICL1)可以同时与2个NBD结构域结合,而第2个偶联螺旋(位于ICL2) 专一的与半转运体对面的NBD结构域结合[35]。在NBD结构域中Q-环提供了与偶联螺旋相互作用的氨基酸残基。NBD结构域中有1个保守的位于X-环(X-loop)谷氨酸盐可以同时与两个偶联螺旋结合被认为在NBDs与MDs的相互作用中起重要作用[35]。NBD和MD偶联的确切机理还不清楚。
3.3底物识别结构域 摄取系统而不是输出系统通常包含可以与底物识别和结合的底物结合受体(solute-binding receptors,SBP),一个系统通常含有一个或多个底物结合受体,在革兰氏阴性菌中底物结合受体位于细胞周质(periplasm),革兰氏阴性菌如大肠杆菌的原核型ABC转运体的基质结合蛋白和自由状态的基质结合蛋白在胞外空间含量较高。其比例远高于可能与他们结合的ABC转运体。可能额外的SBPs能够结合可能结合的任何底物,经SBPs将底物交给附近的ABC输入泵完成底物转运。在革兰氏阴性菌的外排泵中需要确保ABC转运体与外膜蛋白结合便于穿过膜周间隙的转运,如大肠杆菌外排溶血素系统HlyB[8]。HlyB转运体转移溶血素时与外膜蛋白TolC相互作用,在外膜与膜周间隙形成一个跨膜转移溶血素的外排系统。
在革兰氏阳性菌中缺乏外膜的SBPs暴露在细胞表面,因此底物结合受体既可以是脂蛋白也可以位于细胞质膜外,或细胞表面相关蛋白,底物结合蛋白分为6个亚家族[36]。SBP与MT和NBD形成的同源或异源二聚体的N端或C端结合,因此2个或更多的SBP可以与二聚体结合可以促进转运效率。而有些特殊的高度亲和的ABC家族的摄取系统如维生素、矿物质和其他小分子摄取系统叫做ECF系统,缺乏细胞外的受体[37]。
大肠杆菌是最简单的模式生物,对其它ABC家族转运体系统进行研究不仅可以了解该物质ABC家族转运系统的转运机制,同时也可以为其他物种的相关研究提供理论参考,大肠杆菌的外排泵家族中有一部分与其抗药性有关,对这些外排泵的深入研究对于解读人类的抗肿瘤药物的外排有一定的帮助。
