不同培肥方式对土壤有机碳与微生物群落结构的影响*
2018-11-29冶秀香杨金娟牛红霞
李 倩, 马 琨, 冶秀香, 杨金娟, 牛红霞, 马 玲
不同培肥方式对土壤有机碳与微生物群落结构的影响*
李 倩1,2†, 马 琨2†, 冶秀香1, 杨金娟1, 牛红霞1, 马 玲1
(1. 宁夏大学农学院 银川 750021; 2. 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地 银川 750021)
为揭示旱作区耕地土壤有机碳累积规律及其与土壤微生物群落间的相互作用机制, 试验采用磷脂脂肪酸(PLFA)指纹图谱及土壤腐殖质形态分组的方法, 通过田间定位试验, 研究了马铃薯-马铃薯-油用向日葵-马铃薯-油用向日葵轮作模式下, 有机、无机肥配施(不施肥、单施化肥、化肥配施牛粪、化肥配施羊粪、化肥配施生物有机肥、化肥配施黄腐酸钾)对土壤有机碳累积、土壤腐殖质形态的影响及其与土壤微生物群落结构间的相互关系。结果表明: 在连续培肥5年间, 随培肥时间延长, 土壤有机碳呈波动性上升趋势。与对照相比, 化肥配施牛粪、化肥配施羊粪处理土壤有机碳以年6.61%和8.97%的增长率累积增加, 不同处理外源有机碳含量及有机肥种类的差异影响了土壤有机碳的累积速率。化肥配施高量有机肥(化肥+羊粪、化肥+牛粪)处理显著提高了土壤稳结态、松结态腐殖质含量及松结态/紧结态腐殖质的比例, 且以PLFA表征的土壤细菌、真菌、放线菌、原生动物、土壤微生物群落总生物量与对照处理间均有显著性差异(<0.05)。与对照相比, 各施肥处理的革兰氏阳性菌/革兰氏阴性菌(G+/G-)值均呈降低趋势; 但不同有机无机相结合的土壤培肥方式对土壤G+/G-的比例没有显著差异。多元分析表明, 基于土壤微生物主要类群磷脂脂肪酸含量的排序轴与基于土壤有机碳、腐殖质形态的排序轴之间相关性(1=0.568,2=0.611)较好, 累积变量在98.69%上揭示不同有机无机培肥措施影响下的土壤微生物群落生物量与环境因子间的相互关系。土壤松结态腐殖质含量与土壤G+/G-比值正相关。外源有机碳的施入促进了土壤紧结态腐殖碳向稳结态、松结态腐殖质转化; 较高量外源有机碳施入有助于提升土壤细菌、真菌的生物量。总体而言, 土壤微生物群落结构的变化是受有机无机培肥措施所引起的土壤有机碳含量、腐殖质形态变化驱动; 化肥配施牛粪和化肥配施羊粪有利于土壤有机碳积累和松结态腐殖质的形成, 促进土壤中微生物生物量提高。研究结果可为宁夏中部干旱区土壤合理培肥提供科学依据。
有机无机培肥; 土壤有机碳; 微生物群落; 松结态腐殖质; 紧结态腐殖质; 磷脂脂肪酸
土壤有机质是土壤的重要组成部分, 对土壤生态系统功能至关重要[1]。土壤有机质含量高低反映了有机碳的输入和矿化之间的长期平衡关系, 土壤有机碳的转化对维持土壤质量和生态系统的长期生产力有重要作用[2]。有研究表明: 有机肥与化肥配施是砂姜黑土的最佳培肥模式[3], 是提升红壤水稻土有机质数量、质量的最佳培肥措施[4]。有机、无机肥配施能够降低土壤有机碳的抗氧化能力, 显著增加土壤活性有机碳及腐殖质各组分的含量[5]。肥料配比方式、耕作制度、作物类别的不同都会导致土壤物理、化学和生物学特性的改变而引起培肥效果的差异。土壤有机质数量和质量在很大程度上决定了土壤的肥力和生产力, 有机、无机肥配施是保持土壤可持续生产力的有效途径。
土壤微生物会影响土壤有机质的分解及土壤发育过程[6-7]。相关研究认为, 土壤微生物多样性和活性是可持续农业生态系统的标志, 土壤微生物多样性和丰度是生态系统维护的基础[8-9]。土壤微生物结构和功能的变化很大程度上是由土壤有机质差异造成的。因此, 土壤有机碳含量被认为是决定土壤微生物动力学的主要因素[10]。Wei等[11]通过连续35年不同培肥方式对土壤微生物影响的研究认为, 施肥对微生物群落结构的影响是通过调节钙质潮土中土壤碳和养分的有效性来实现的。施入土壤中外源有机碳及土壤易分解有机碳含量的提高都会影响土壤微生物生物量和微生物活性[12-13]。有机碳的输入增加了土壤细菌和真菌的PLFA生物标记[10]; 与单施化肥相比, 有机肥的施用显著提高了土壤中以PLFA表征的总微生物生物量、细菌、革兰氏阳性菌的生物量, 但革兰氏阴性菌、放线菌、真菌的生物量降低, 革兰氏阳性菌/革兰氏阴性菌(G+/G-)比值增大, 真菌/细菌(F/B)比值降低[11]。然而, 郭芸等[14]发现, 长期有机、无机肥配施不仅显著提高了土壤微生物总PLFA和真菌、细菌、放线菌的生物量, 也提高了F/B的比值; 在短期内, 无机和有机肥料配施对土壤细菌多样性及其活性的影响相对较小[15]。土壤有机碳的积累量及有机碳对微生物群落结构的影响是否会因施入有机肥种类的不同而存在差异?
宁夏中部干旱区土壤有机质含量较低, 有机碳矿化剧烈, 土壤矿物对有机质物理性保护作用较弱。由于自然条件差, 土壤肥力持续下降导致的农作物生产形势比较严峻。水资源短缺也导致当地农业生产灌溉方式发生变化。大面积的滴灌、喷灌方式应用后, 土壤有机碳形态、累积规律及其与土壤微生物群落间的相互关系是否仍然与传统农业管理措施相一致? 我们假设, 在旱作滴灌条件下, 不同类型有机肥料与无机肥料配施, 土壤微生物群落组成的变化仍然是受土壤有机碳差异驱动的; 土壤有机碳、土壤腐殖质主要形态组成与土壤微生物群落组成有正相关关系; 外源高量有机肥的施入对土壤有机碳的积累和微生物群落结构的调节作用最显著。为此, 本文试验比较了化肥配施羊粪、牛粪、黄腐酸钾和生物有机肥处理之间的差异, 旨在进一步探索肥料类型、外源有机碳输入量对土壤有机碳和微生物群落结构影响的作用机理。研究结果将有助于揭示旱作区土壤有机碳与土壤微生物群落组成间的相互关系及其作用机制, 能为宁夏中部干旱区土壤合理培肥提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于宁夏同心县王团镇北村(36°51′6″N, 105°54′7″E), 海拔1 568 m。年降水量272.6 mm, 年蒸发量2 740 mm。土壤类型为灰钙土, 质地为砂质壤土。试验前土壤全氮0.18 g∙kg-1、全磷0.65 g∙kg-1、有机碳3.35 g∙kg-1、碱解氮9.13 mg∙kg-1、速效磷9.32 mg∙kg-1、速效钾97.08 mg∙kg-1、pH为9.20。
1.2 试验设计
试验始于2011年, 采用马铃薯()-马铃薯-油用向日葵()-马铃薯-油用向日葵的轮作模式。2011年、2012年、2014年种植的马铃薯品种为‘冀张薯8号’、‘冀张薯8号’、‘陇薯3号’, 2013年、2015年种植油用向日葵品种为‘S606’。
试验采用单因素随机区组设计, 6个处理, 3次重复, 小区面积6 m×15 m。T1: 不施肥; T2: 单施化肥; T3: 化肥+牛粪, 牛粪施用总量为37 500 kg·hm-2·a-1(有机碳总量为10 110 kg·hm-2·a-1); T4: 化肥+羊粪, 羊粪施用总量为37 500 kg·hm-2·a-1(有机碳总量为12 555 kg·hm-2·a-1); T5:化肥+生物有机肥, 施用总量为1 200 kg·hm-2·a-1(有机碳总量约为210 kg·hm-2·a-1), 生物有机肥由山东泉林嘉有机肥料有限责任公司生产, 其中芽孢杆菌(胶冻样芽孢杆菌+巨大芽孢杆菌)≥0.2亿×g-1, 黄腐酸含量约为195 kg·hm-2·a-1; T6: 化肥+黄腐酸钾, 黄腐酸钾施用总量1 200 kg·hm-2·a-1(有机碳总量约为315 kg·hm-2·a-1), 黄腐酸钾含有机质≥30%、黄腐酸≥18%、K2O≥4%。
马铃薯种植时各处理施肥量中N∶P2O5∶K2O= 12∶6∶7, 折合纯氮390 kg·hm-2·a-1(70%基施, 30%追施)、P2O5195 kg·hm-2·a-1、K2O 210 kg·hm-2·a-1。油葵种植时N∶P2O5∶K2O=12∶6∶4, 折合纯氮390 kg·hm-2·a-1(70%基施, 30%追施)、P2O5195 kg·hm-2·a-1、K2O 120 kg·hm-2·a-1, 有机肥全部基施。作物全生育期滴灌量为120 m3。
每年作物收获时多点取样, 采集各小区0~20 cm土层土壤样本, 低温储存带回实验室, 过1 mm筛, 储存在-40 ℃冰箱, 用于土壤微生物结构(PLFA)测定。部分样品风干, 用于土壤理化性状测定。
1.3 分析方法
土壤总有机碳采用重铬酸钾容量法[16]、腐殖质组成测定采用结合态腐殖质分组法(熊毅—付积平改进法)[17]、土壤微生物群落结构采用PLFA法。称取相当于4 g干土重的新鲜土样, 依照魏常慧等[18]的提取步骤, 利用中国科学院青藏高原研究所高寒生态研究室的Agilent 6850型气相色谱仪, 在进样口为250 ℃、H2流量为40 mL·min-1(0.4 MPa)、氮气(0.4 Mpa)、色谱柱为Agilent HP-5MS, 30 µm×250 µm×0.25 µm, 柱温: 170 ℃下测定, MIDI Sherlock脂肪酸图谱微生物鉴定系统分析待检样品。
1.4 数据分析
DPS 9.50软件进行方差分析和多重比较, Canoco 5.0软件进行多元分析。
2 结果与分析
2.1 无机肥与不同有机肥配施对土壤总有机碳累积的影响
有机无机肥配合施用是实现可持续肥料管理策略的一种有效途径和方法。减少化学肥料用量, 增加有机肥料的施用可以优化土壤微生物所驱动的土壤营养元素的内部循环[19]。由图1可以看出, 随培肥时间的延长, 土壤有机碳呈波动性上升趋势。有机无机肥配施5年后(2015年), 化肥配施牛粪、化肥配施羊粪处理土壤有机碳含量较试验初期有明显累积, 且显著高于其他处理(<0.05)。其中, 2015年时化肥配施牛粪、化肥配施羊粪处理土壤有机碳含量较2011年培肥初期分别增加38.00%和58.98%; 与对照(T1)相比(2015年), 土壤有机碳含量也增加33.05%和44.87%。化肥配施牛粪、化肥配施羊粪处理的土壤有机碳以年6.61%和8.97%的增长率累积增加, 与化肥配施生物有机肥、黄腐酸钾和单施化肥处理相比, 有机碳累积速度提高12.16~16.89倍。然而, 化肥配施生物有机肥、黄腐酸钾及单施化肥下土壤有机碳含量与对照相比无明显差异。可见, 土壤有机碳的累积受施入土壤有机肥中碳含量高低的影响, 较高量有机肥的施入更有利于土壤有机碳的累积。
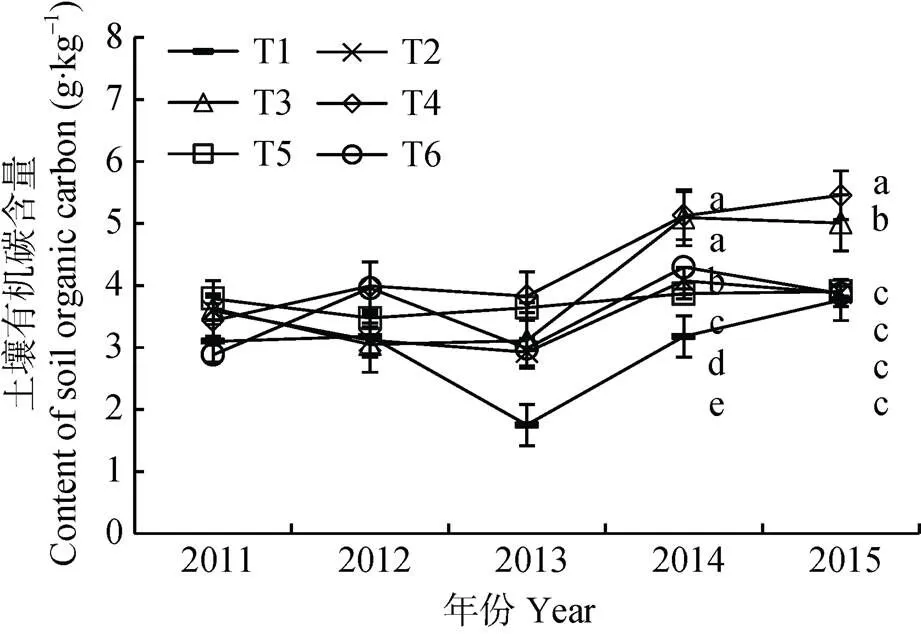
图1 不同有机无机肥配施处理对土壤有机碳的影响
T1、T2、T3、T4、T5、T6分别表示不施肥、单施化肥、化肥+牛粪、化肥+羊粪、化肥+生物有机肥、化肥+黄腐酸钾。不同小写字母表示不同处理间差异显著(<0.05)。T1, T2, T3, T4, T5 and T6 represent blank control, inorganic fertilizer application, combined application of inorganic fertilizer and cow dung, combined application of inorganic fertilizer and sheep manure, combined application of inorganic fertilizer and bioorganic fertilizer, and combined application of inorganic fertilizer and fulvic acid potassium. Different lowercase letters indicate significant differences among different treatments (< 0.05).
2.2 无机肥与不同有机肥配施对土壤腐殖质形态组成的影响
土壤腐殖质是土壤有机质的主要组成部分, 按其结合牢固程度可分为稳结态、紧结态、松结态。紧结态腐殖质对碳的固定和土壤结构有较大的影响, 而松结态腐殖质对土壤养分的释放起着重要作用, 土壤肥力水平主要受松结态腐殖质所占比例的影响[20-21]。
化肥配施不同有机肥, 随有机肥带入土壤有机碳含量的增加, 土壤稳结态腐殖质、松结态腐殖质、紧结态腐殖质含量均显著提高(表1), 不同处理间有显著性差异(<0.05)。与化肥配施生物有机肥、化肥配施黄腐酸钾处理(化肥配施低量有机碳)相比, 化肥配施牛粪、化肥配施羊粪处理(化肥配施高量有机碳)土壤稳结态腐殖质、松结态腐殖质、紧结态腐殖质含量分别提高1.34~1.65倍、1.07~1.13倍、1.36~1.52倍。与对照(T1)相比, 化肥配施高量有机碳对土壤稳结态腐殖质、松结态腐殖质含量提高的幅度明显高于低量有机碳配施化肥处理。此外, 与化肥配施低量有机碳处理相比, 化肥配施高量有机碳处理的土壤松结态/紧结态腐殖质的比例约提高1.27~1.43倍。不施肥及单施化肥处理与其余培肥处理相比, 土壤松结态/紧结态腐殖质比例最低, 仅相当于无机有机配施培肥处理的32.61%~46.88%。土壤紧结态腐殖质含量未受外源有机肥施用带入土壤有机碳数量多少的影响。与对照相比, 单施化肥仅显著提高了土壤稳结态腐殖质含量。持续5年培肥后, 有机无机相结合的培肥方式与对照之间土壤重组腐殖质均有显著性差异(<0.05)。与对照相比, 有机无机结合培肥处理的土壤重组腐殖质含量增幅达8.83%~48.20%; 化肥配施牛粪和化肥配施羊粪处理的土壤重组腐殖质含量较对照提高1.48~1.68倍, 与化肥配施生物有机肥、化肥配施黄腐酸钾处理相比也提高4.10%~33.29%。
总体可见, 在化肥配施有机肥下土壤腐殖质结合形态的变化主要是通过降低土壤腐殖质紧结态比例, 提高土壤稳结态腐殖质、松结态腐殖质含量以及松结态腐殖质/紧结态腐殖质比例实现的。在施用化肥的基础上施入有机肥, 使土壤松结态腐殖质含量提高、土壤松结态/紧结态腐殖质比例提升, 可能是有机无机配施培肥促进土壤养分释放, 提高土壤肥力的主要原因。
2.3 无机肥与不同有机肥配施对土壤微生物群落结构的影响
土壤中总磷脂脂肪酸代表着土壤微生物的总量, 是反映土壤微生物群落结构变化的敏感指标[22]。由表2可以看出, 无机肥与不同有机肥配施处理下以PLFA表征的土壤微生物群落结构间存在较大的差异。化肥配施牛粪和化肥配施羊粪的土壤细菌、真菌、放线菌、原生动物、土壤微生物总磷脂脂肪酸与对照(T1)处理间均有显著性差异(<0.05)。与对照和单施化肥处理相比, 其土壤微生物总磷脂脂肪酸分别提高1.64~3.40倍和1.92~3.99倍。化肥配施牛粪处理土壤细菌、放线菌、真菌的生物量较对照处理分别增加3.42倍、1.90倍和3.90倍, 但以PLFA表征的主要微生物类群生物量增长幅度却低于化肥配施羊粪, 这两种处理除真菌在化肥配施羊粪处理中显著高于化肥配施牛粪处理外, 其余微生物之间差异均不显著。化肥配施生物有机肥和化肥配施黄腐酸钾处理的土壤细菌、真菌、放线菌生物量与对照相比也表现出类似的变化趋势。其中, 化肥配施生物有机肥处理与对照间有显著性差异(<0.05), 但化肥配施黄腐酸钾处理除真菌生物量, 土壤细菌及放线菌与对照间无明显差异。

表1 不同有机无机肥配施处理对土壤腐殖质形态及组分数量的影响(2015年)
T1、T2、T3、T4、T5、T6分别表示不施肥、单施化肥、化肥+牛粪、化肥+羊粪、化肥+生物有机肥、化肥+黄腐酸钾。同列不同小写字母表示处理间差异显著(<0.05)。T1, T2, T3, T4, T5 and T6 represent blank control, inorganic fertilizer application, combined application of inorganic fertilizer and cow dung, combined application of inorganic fertilizer and sheep manure, combined application of inorganic fertilizer and bioorganic fertilizer, and combined application of inorganic fertilizer and fulvic acid potassium. Different lowercase letters in the same column indicate significant differences among treatments at 0.05 level.
与对照相比, 各处理中G+/G-值均呈显著(<0.05)降低趋势, 但不同有机无机结合的土壤培肥方式下, 土壤G+/G-值间无显著差异。与对照相比, 土壤真菌/细菌生物量比值除化肥配施牛粪、羊粪外, 其他处理整体间差异不明显。可见, 在旱作条件下, 有机无机相结合的土壤培肥方式仍可以改变土壤微生物群落结构组成。土壤主要微生物类群生物量及不同微生物类群间比例与培肥方式、肥料类别、外源有机碳数量有明显关系。
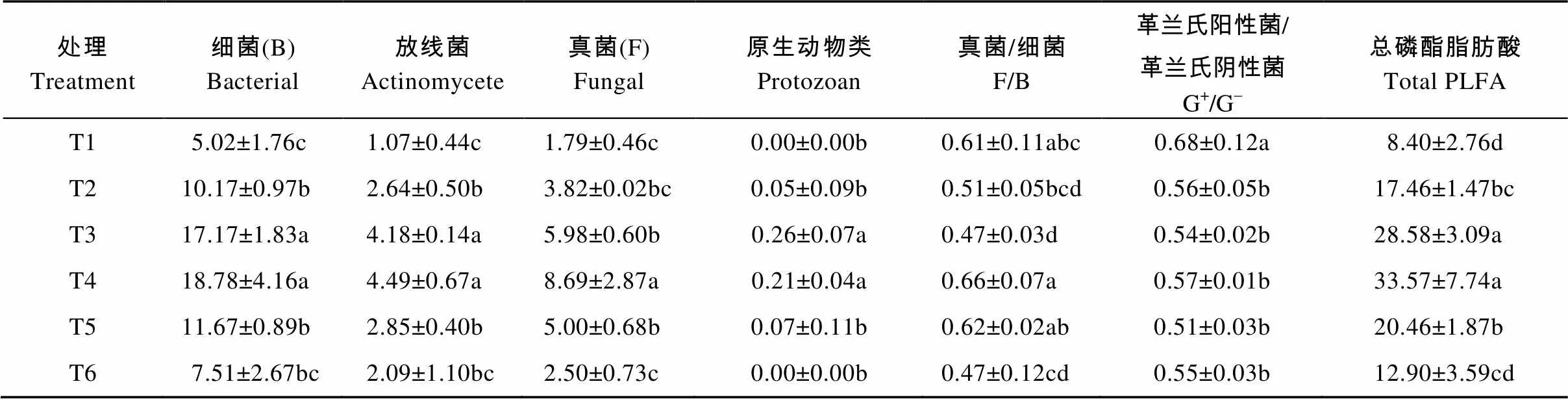
表2 不同有机无机肥配施处理对土壤微生物群落结构组成的影响
T1、T2、T3、T4、T5、T6分别表示不施肥、单施化肥、化肥+牛粪、化肥+羊粪、化肥+生物有机肥、化肥+黄腐酸钾。同列不同小写字母表示处理间差异显著(<0.05)。 T1, T2, T3, T4, T5 and T6 represent blank control, inorganic fertilizer application, combined application of inorganic fertilizer and cow dung, combined application of inorganic fertilizer and sheep manure, combined application of inorganic fertilizer and bioorganic fertilizer, and combined application of inorganic fertilizer and fulvic acid potassium. Different lowercase letters in the same column indicate significant differences among treatments at 0.05 level.
2.4 土壤微生物群落结构组成与土壤有机碳及土壤腐殖质形态间的相互关系
以PLFA表征的土壤主要微生物类群的生物量与土壤有机碳含量均呈正相关(图2a), 主成分分析中第1排序轴(97.16%)、第2排序轴(1.53%)能够在累积变量98.69%水平上揭示不同有机无机培肥影响下的土壤微生物群落变化。化肥配施牛粪和化肥配施羊粪处理的土壤细菌、真菌等主要微生物类群在空间分布上相对聚集, 两处理间的微生物群落结构组成较相似。与对照(T1)相比, 单施化肥、化肥配施生物有机肥和化肥配施黄腐酸钾处理主要土壤微生物类群也分布在不同空间位置上, 说明这些处理的土壤微生物群落结构组成彼此间有差异。此外, 土壤微生物群落组成中G+/G-比值与土壤松结态腐殖质含量间有较好的正相关关系。总体上, 基于土壤微生物主要类群PLFA含量排序轴与基于土壤有机碳及腐殖质形态等环境因子的排序轴之间相关性(1=0.568,2=0.611)较好。
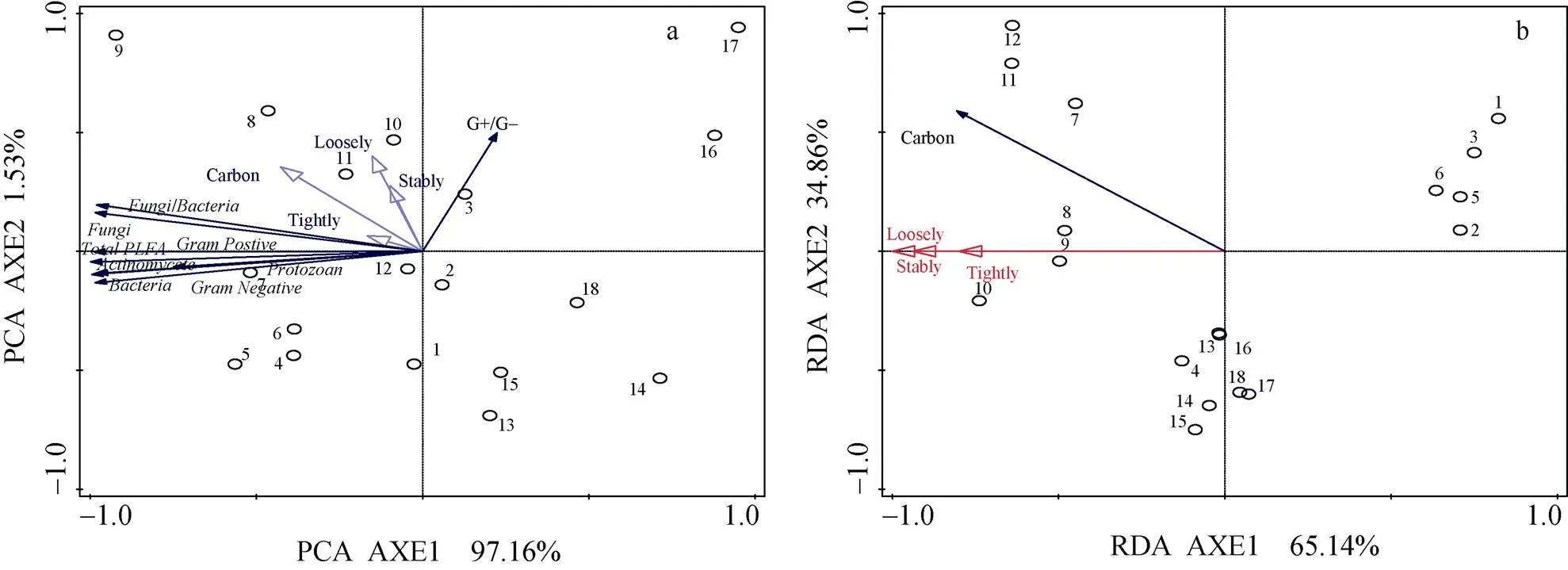
图2 不同有机无机肥配施处理下土壤微生物群落结构组成(a)、有机碳及腐殖质形态(b)的多元分析
图中1、2、3为不施肥处理, 4、5、6为单施化肥处理, 7、8、9为化肥配施牛粪处理, 10、11、12为化肥配施羊粪处理, 13、14、15为化肥配施生物有机肥处理, 16、17、18为化肥配施黄腐酸钾处理。: 细菌;: 真菌;: 放线菌;: 原生动物;: 革兰氏阳性菌;: 革兰氏阴性菌; Tightly: 紧结态腐殖质; Loosely: 松结态腐殖质; Stably: 稳结态腐殖质; Carbon: 土壤有机碳;: 总磷脂脂肪酸。In the figures, 1, 2, 3 are replicates of no fertilization treatments; 4, 5, 6 are replicates of chemical fertilizer treatment; 7, 8, 9 are replicates of treatment of combined application of chemical fertilizer and cow dung; 10, 11, 12 are replicates of treatment of combined application of chemical fertilizer and sheep manure; 13, 14, 15 are replicates of treatment of combined application of chemical fertilizer and bio-organic manure; 16, 17, 18 are replicates of treatment of combined application of chemical fertilizer and fulvic acid potassium.e: gram positive bacteria;: gram negative bacteria; Tightly: tightly combined humus; Loosely: loosely combined humus; Stably:stably combined humus; Carbon: soil organic carbon;: total phospholipid fatty acids.
图2b中, 土壤有机碳含量与松结态、稳结态和紧结态腐殖质含量均正相关。冗余分析(RDA)表明, 第1排序轴、第2排序轴分别解释了土壤有机碳变化的65.14%和34.86%。假设性测验结果表明(pseudo-=8.7,=0.001 7), 所有排序轴都是极显著的; 第1排序轴对有机无机培肥影响下的土壤有机碳累积差异变化解释最多。与对照相比, 土壤有机碳在空间分布上有明显分散。化肥配施生物有机肥和化肥配施黄腐酸钾处理的土壤腐殖质碳的结合形态比较相似; 化肥配施牛粪、化肥配施羊粪处理土壤腐殖质碳结合形态相一致, 单施化肥处理没有明显影响土壤腐殖质碳结合形态的空间分布。
可见, 培肥方式会对土壤腐殖质的形态组成产生影响, 较高量外源有机碳(化肥配施牛粪和化肥配施羊粪处理)的施入会明显影响土壤有机碳的累积。各形态腐殖质含量与土壤有机碳含量都呈正相关, 松结态腐殖质对化肥配施牛粪和化肥配施羊粪处理的微生物群落结构组成影响较大; 配施高量有机肥表现有提高土壤松结态、稳结态腐殖质含量的趋势。
3 讨论
3.1 无机肥与不同有机肥配施对土壤有机碳累积及腐殖质形态组成的影响
有机碳能改善土壤的物理、化学和生物特性, 在植物养分循环中起着重要作用, 是衡量土壤质量的重要指标之一[23]。Guo等[24]发现, 在小麦()-玉米()轮作系统中土壤有机质与施入土壤的牛粪数量有显著正相关, 单施化肥对土壤有机碳累积量没有显著影响。每公顷施用200 m3的罂粟()废弃物能够将土壤有机碳含量从1.24%增加到1.57%, 然而较低用量对土壤有机碳的增加没有明显影响[25]。试验中, 化肥配施牛粪、羊粪处理土壤有机碳含量平均从6.09 g∙kg-1提高到9.02 g∙kg-1。与试验初期相比, 年平均有机碳累积递增幅度可达9.71%, 土壤有机碳累积较明显。然而, 化肥配施生物有机肥和化肥配施黄腐酸钾在培肥5年后, 土壤有机碳的累积量仅比对照增加2.65%~ 3.70%。研究结果与相关报道基本一致。分析认为, 外源有机碳的输入、作物高产导致的残落物中有机物的增加可能是化肥有机肥配施提高土壤有机碳累积的两个主要途径。较高量有机肥的施入有利于土壤有机碳的快速积累, 低量或者不施有机肥对土壤有机碳累积影响不明显。但是, Campbell等[26]认为, 富含有机质的土壤即使持续施肥(包括豆科绿肥或豆科牧草作物还田)也很难增加和改善土壤有机质的含量, 施肥措施并没有影响碳的矿化。
土壤腐殖质是有机质存在的一种特殊形态, 施入土壤中未被分解的有机物通过腐殖化过程转变为腐殖质。由于腐殖质碳对土壤微生物分解有较强的抗性, 因此土壤腐殖质的周转比较缓慢。但是, 土壤腐殖质在提供植物营养物质、矿物分解方面起着重要的作用[27]。有研究指出, 腐殖质的形态与土壤有机碳的储存量和土壤的形成因素有关[28]。施用马粪和马粪配施高量化肥均提高了土壤有机碳的含量, 但对土壤腐殖物质——胡敏酸数量影响不显著。施用马粪下, 土壤2.00~0.25 mm大团聚体中胡敏酸的芳香性增强, 胡敏酸更稳定[29]。长期平衡施肥, 特别是有机肥施用增加了O—烷基、异丙基和N—CH基团的百分比, 降低了腐殖质中芳香族的含量[30]。试验中有机无机结合的培肥方式下, 随进入土壤有机物数量的增加, 土壤腐殖质含量有逐步增加的趋势。其中稳结态、松结态腐殖质含量均显著提高, 紧结态腐殖质没有受到明显影响。尽管各培肥措施都在不同程度上降低了紧结态腐殖质的相对质量分数, 但土壤中腐殖质碳的结合方式仍以紧结态腐殖质为主。紧结态腐殖所占的质量分数较对照处理下降5.84%~25.41%, 而稳结态、松结态腐殖质所占的质量分数则约提高0.55~1.93倍。单施化肥处理对土壤腐殖质形态组成的调节作用较弱, 化肥配施生物有机肥和化肥配施黄腐酸钾处理次之, 化肥配施羊粪和化肥配施牛粪处理作用最强。可见, 随外源有机碳的施入及输入量的增加, 有机碳输入和土壤矿化之间的平衡关系会被打破, 土壤紧结态腐殖碳会向稳结态、松结态碳转化。这与韩晓萍等[31]在有机无机长期配施对小麦产量和土壤有机组分的影响所得的结果基本一致。土壤腐殖质与土壤无机胶体结合的松紧程度不同, 对土壤肥力的贡献亦不相同。松结态/紧结态腐殖质的比例大意味着腐殖质活性较高。试验中, 土壤有机碳含量与松结态、稳结态和紧结态腐殖质含量均呈正相关。单施化肥并没有改变土壤松结态/紧结态的比例, 而配施化肥下高量外源有机碳的输入后土壤松/紧结合态碳的比例明显大于低量有机碳输入。可见, 长期坚持有机无机相结合的培肥方式, 可促进有机质活化, 改善土壤供肥能力。
3.2 土壤有机碳对土壤微生物群落结构的影响及其相互关系
土壤水分、有机质和氮是影响土壤微生物群落组成的最重要环境因素, 在土壤中添加有机碳可以增加土壤微生物的丰富度和活性[32]。在施用有机肥的条件下, 较高土壤有机碳含量与较高的土壤微生物生物量、活性是相互一致的[12,14]。在塔斯马尼亚富铁土壤上使用磷脂脂肪酸甲酯表征的微生物碳与土壤总有机碳之间也有显著正相关关系(=0.57)[25]。水稻()-水稻-休闲种植制度下, 长期施用堆肥会显著增加土壤微生物生物量; 土壤微生物生物量的增加是作物根系生长、根系密度增加所引起的植物对肥料的典型反应[33]。本试验中, 有机无机培肥方式均增加了土壤微生物生物量, 随有机肥带入土壤有机碳数量的增加, 土壤细菌、真菌、放线菌生物量和微生物总生物量显著升高。多元分析表明, 土壤微生物生物量与土壤有机碳含量均呈正相关, 这与Thiele-Bruhn等[34]的研究结果相一致。分析认为, 受施入土壤中有机肥种类、性质和数量的影响, 作物产量增加, 导致根系分泌物和植物残落物的增加, 为土壤微生物提供充足碳源, 相应地提高了微生物生物量; 而有机碳输入和矿化之间新的平衡关系逐渐建立, 导致了土壤有机碳的逐步累积。可见, 微生物群落结构的变化是受有机无机培肥措施所引起的土壤有机碳含量变化驱动的。
土壤细菌群落向革兰氏阴性菌群转变是其对氮肥施用的响应, 是以环丙基脂肪酸(cy17:0)的增加和革兰氏阳性菌与革兰氏阴性菌比例的降低为标志的, 土壤G+/G-的比例在施用化肥下比施有机肥更低[35]。以往的研究认为, 在土壤养分较丰富的条件下, 革兰氏阴性菌比例往往较高[36]。有机物料的添加首先会导致生长速度较快的革兰氏阴性菌快速增殖, 随后革兰氏阴性菌增殖速度下降, 最后才有利于生长速度较慢的革兰氏阳性菌或真菌生长。高量有机肥的施入, 促进土壤微生物对外源有机碳的利用而导致更充分的群落演替效应, 主要是革兰氏阳性菌和真菌对土壤有机质利用的不同步变化所引起的[11]。试验结果显示, 与对照相比, 不同有机无机肥配合施用均显著降低了G+/G-比例, 但G+/G-比例关系却不受有机肥施用带入土壤有机碳数量的的影响。试验结果与Giacometti等[35]的研究结果基本一致。然而, Wei等[11]的研究却表明, 有机肥施用能使G+/G-比例增加12.3%。这些相互矛盾的研究结果可能与不同施肥管理措施下, 土壤有机碳和土壤养分的有效性差异有关。
试验中土壤松结态腐殖质含量与土壤有机碳、土壤G+/G-比例均有显著正相关关系。土壤有机碳是影响微生物群落结构变化的关键因素[37]。试验中有机肥料施入有利于革兰氏阴性菌的生长, 这就意味着土壤革兰氏阴性菌对有机碳的输入反应更敏感, 会在整个土壤细菌群落组成中占据主导地位。土壤有机碳和松结态腐殖质含量应该是土壤革兰氏阴性菌相对丰度变化的主要驱动力。可见, 在较好土壤环境条件下, 土壤革兰氏阴性菌较革兰氏阳性菌应该具有竞争优势。通常含有较多有机碳且肥力较高的土壤中细菌数目多于真菌, 真菌利用难分解有机碳的能力优于细菌, 而细菌则容易利用易分解有机碳[11-12]。Zhong等[36]认为由于细菌比真菌的周转时间短, 所以细菌对土壤肥力的短期变化更为敏感。试验中持续有机无机培肥均明显增加了土壤细菌、真菌的生物量, 整体上较高量的外源有机肥施入有助于提升土壤细菌、真菌的生物量。尽管细菌生物量远高于真菌, 但较对照而言, 真菌生物量增加的幅度明显高于细菌。目前, 有关土壤F/B比率变化都归因于植物根系分泌物数量和质量, 枯落物的质量和数量、有机肥的输入和植物生产力及组成的差异[35]。试验中F/B比例关系比较复杂, 除化肥配施牛粪和化肥配施羊粪处理间有显著差异外, 其他处理也表现出受土壤培肥方式、肥料类型、有机肥数量影响而变化的趋势。外源有机肥输入时, 牛粪和羊粪含水量和各自有机质含量的差异有可能就是导致化肥配施牛粪、化肥配施羊粪处理F/B间差异显著的主要原因。
4 结论
研究结果表明: 旱作节水条件下, 连续有机无机培肥5年后, 土壤有机碳含量逐年上升, 以PLFA表征的土壤细菌、真菌、放线菌、原生动物和总微生物生物量增加, G+/G-比值降低。有机无机肥配施通过提高土壤稳结态腐殖质、松结态腐殖质含量以及松结态腐殖质/紧结态腐殖质的比例来影响土壤有机碳的组成。土壤有机碳含量与土壤主要微生物类群生物量、土壤松结态腐殖质含量与G+/G-比值均有正相关关系。外源有机碳的输入促进了主要微生物类群生物量的增加, 提升了土壤革兰氏阴性菌的竞争优势。
[1] HAI L, LI X G, LI F M, et al. Long-term fertilization and manuring effects on physically-separated soil organic matter pools under a wheat-wheat-maize cropping system in an arid region of China[J]. Soil Biology and Biochemistry, 2010, 42(2): 253–259
[2] MOHARANA P C, SHARMA B M, BISWAS D R, et al. Long-term effect of nutrient management on soil fertility and soil organic carbon pools under a 6-year-old pearl millet-wheat cropping system in an Inceptisol of subtropical India[J]. Field Crops Research, 2012, 136: 32–41
[3] 李玮, 乔玉强, 陈欢, 等. 秸秆还田和施肥对砂姜黑土理化性质及小麦-玉米产量的影响[J]. 生态学报, 2014, 34(17): 5052–5061 LI W, QIAO Y Q, CHEN H, et al. Effects of combined straw and N application on the physicochemical properties of lime concretion black soil and crop yields[J]. Acta Ecologica Sinica, 2014, 34(17): 5052–5061
[4] 邓文悦, 柳开楼, 田静, 等. 长期施肥对水稻土不同功能有机质库碳氮分布的影响[J]. 土壤学报, 2017, 54(2): 468–479 DENG W Y, LIU K L, TIAN J, et al. Effects of long-term fertilization on distribution of carbon and nitrogen in different functional soil organic matter fractions in paddy soil[J]. Acta Pedologica Sinica, 2017, 54(2): 468–479
[5] LIU E K, YAN C R, MEI X R, et al. Long-term effect of manure and fertilizer on soil organic carbon pools in dryland farming in northwest China[J]. PLoS One, 2013, 8(2): e56536
[6] CHEN D M, YUAN L, LIU Y R, et al. Long-term application of manures plus chemical fertilizers sustained high rice yield and improved soil chemical and bacterial properties[J]. European Journal of Agronomy, 2017, 90: 34–42
[7] DING J L, JIANG X, MA M C, et al. Effect of 35 years inorganic fertilizer and manure amendment on structure of bacterial and archaeal communities in black soil of northeast China[J]. Applied Soil Ecology, 2016, 105: 187–195
[8] SU J Q, DING L J, XUE K, et al. Long-term balanced fertilization increases the soil microbial functional diversity in a phosphorus-limited paddy soil[J]. Molecular Ecology, 2015, 24(1): 136–150
[9] LI Y Y, CHEN L Q, WEN H Y, et al. 454 Pyrosequencing analysis of bacterial diversity revealed by a comparative study of soils from mining subsidence and reclamation areas[J]. Journal of Microbiology and Biotechnology, 2014, 24(3): 313–323
[10] ZHANG Q C, SHAMSI I H, XU D T, et al. Chemical fertilizer and organic manure inputs in soil exhibit a vice versa pattern of microbial community structure[J]. Applied Soil Ecology, 2012, 57: 1–8
[11] WEI M, HU G Q, WANG H, et al. 35 years of manure and chemical fertilizer application alters soil microbial community composition in a Fluvo-aquic soil in Northern China[J]. European Journal of Soil Biology, 2017, 82: 27–34
[12] DEMOLING F, FIGUEROA D, BÅÅTH E. Comparison of factors limiting bacterial growth in different soils[J]. Soil Biology and Biochemistry, 2007, 39(10): 2485–2495
[13] BLAGODATSKAYA E, KUZYAKOV Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review[J]. Biology and Fertility of Soils, 2008, 45(2): 115–131
[15] SHEN J P, ZHANG L M, GUO J F, et al. Impact of long-term fertilization practices on the abundance and composition of soil bacterial communities in Northeast China[J]. Applied Soil Ecology, 2010, 46(1): 119–124
[16] 鲍士旦.土壤农化分析[M]. 第3版. 北京: 中国农业出版社, 2000: 30–34 BAO S D. Soil and Agricultural Chemistry Analysis[M]. 3rd ed. Beijing: China Agriculture Press, 2000: 30–34
[17] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000: 121–122 LU R K. Soil Agrochemical Analysis Methods[M]. Beijing: China Agricultural Science and Technology Press, 2000: 121–122
[18] 魏常慧, 刘亚军, 冶秀香, 等. 马铃薯/玉米间作栽培对土壤和作物的影响[J]. 浙江大学学报: 农业与生命科学版, 2017, 43(1): 54–64 WEI C H, LIU Y J, YE X X, et al. Effects of intercropping potato with maize on soil and crop[J]. Journal of Zhejiang University: Agriculture & Life Sciences, 2017, 43(1): 54–64
[19] ZHAO J, NI T, LI J, et al. Effects of organic-inorganic compound fertilizer with reduced chemical fertilizer application on crop yields, soil biological activity and bacterial community structure in a rice–wheat cropping system[J]. Applied Soil Ecology, 2016, 99: 1–12
[20] 张电学, 韩志卿, 王秋兵, 等. 不同施肥制度下褐土结合态腐殖质动态变化[J]. 沈阳农业大学学报, 2006, 37(4): 597–601 ZHANG D X, HAN Z Q, WANG Q B, et al. Dynamic change of soil combined humus in cinnamon soil under different fertilizer regimes[J]. Journal of Shenyang Agricultural University, 2006, 37(4): 597–601
[21] 关文玲, 王旭东, 李利敏, 等. 长期不同施肥条件下土壤腐殖质动态变化及存在状况研究[J]. 干旱地区农业研究, 2002, 20(2): 32–35 GUAN W L, WANG X D, LI L M, et al. Study on the properties and existing state of the soil humus under different long-term fertilizations[J]. Agricultural Research in the Arid Areas, 2002, 20(2): 32–35
[22] BARREIROA, FONTÚRBEL M T, LOMBAO A, et al. Using phospholipid fatty acid and community level physiological profiling techniques to characterize soil microbial communities following an experimental fire and different stabilization treatments[J]. CATENA, 2015, 135: 419–429
[23] KALBITZ K, SOLINGER S, PARK J H, et al. Controls on the dynamics of dissolved organic matter in soils: A review[J]. Soil Science, 2000, 165(4): 277–304
[24] GUO L Y, WU G L, LI Y, et al. Effects of cattle manure compost combined with chemical fertilizer on topsoil organic matter, bulk density and earthworm activity in a wheat-maize rotation system in Eastern China[J]. Soil and Tillage Research, 2016, 156: 140–147
[25] COTCHING W E. Organic matter in the agricultural soils of Tasmania, Australia — A review[J]. Geoderma, 2018, 312: 170–182
[26] CAMPBELL C A, ZENTNER R P, BOWREN K E, et al. Effect of crop rotations and fertilization on soil organic matter and some biochemical properties of a thick Black Chernozem[J]. Canadian Journal of Soil Science, 1991, 71(3): 377–387
[27] 王旭东, 张一平, 姚永斌. 长期不同施肥对土壤腐殖质性质及存在形态的影响[J]. 陕西农业科学, 1997, (2): 6–8 WANG X D, ZHANG Y P, YAO Y B. Effects of long-term different fertilization on soil humus properties and existing forms[J]. Shaanxi Journal of Agricultural Sciences, 1997, (2): 6–8
[28] BONIFACIO E, FALSONE G, PETRILLO M. Humus forms, organic matter stocks and carbon fractions in forest soils of northwestern Italy[J]. Biology and Fertility of Soils, 2011, 47(5): 555–566
[29] 关松, 窦森, 马丽娜, 等. 长施马粪对暗棕壤团聚体腐殖质数量和质量的影响[J]. 土壤学报, 2017, 54(5): 1195–1205 GUAN S, DOU S, MA L N, et al. Effects of long-term application of horse manure on quantity and quality of humic substance in aggregates of dark brown soil[J]. Acta Pedologica Sinica, 2017, 54(5): 1195–1205
[30] XU J S, ZHAO B Z, CHU W Y, et al. Altered humin compositions under organic and inorganic fertilization on an intensively cultivated sandy loam soil[J]. Science of the Total Environment, 2017, 601/602: 356–364
[31] 韩晓萍, 陶云彬, 章哲, 等. 有机无机长期配施对小麦产量和土壤有机组分的影响[J]. 陕西农业科学, 2018, 64(1): 14–17 HAN X P, TAO Y B, ZHANG Z, et al. Effects of long-term organic combined with inorganic fertilization on wheat yield and soil organic components[J]. Shaanxi Journal of Agricultural Sciences, 2018, 64(1): 14–17
[32] YUAN Y, DAI X Q, XU M, et al. Responses of microbial community structure to land-use conversion and fertilization in southern China[J]. European Journal of Soil Biology, 2015, 70: 1–6
[33] NAYAK D R, BABU Y J, ADHYA T K. Long-term application of compost influences microbial biomass and enzyme activities in a tropical Aeric Endoaquept planted to rice under flooded condition[J]. Soil Biology and Biochemistry, 2007, 39(8): 1897–1906
[34] THIELE-BRUHN S, BLOEM J, DE VRIES F T, et al. Linking soil biodiversity and agricultural soil management[J]. Current Opinion in Environmental Sustainability, 2012, 4(5): 523–528
[35] GIACOMETTI C, DEMYAN M S, CAVANI L, et al. Chemical and microbiological soil quality indicators and their potential to differentiate fertilization regimes in temperate agroecosystems[J]. Applied Soil Ecology, 2013, 64: 32–48
[36] ZHONG W H, GU T, WANG W, et al. The effects of mineral fertilizer and organic manure on soil microbial community and diversity[J]. Plant and Soil, 2010, 326(1/2): 511–522
[37] DRENOVSKY R E, VO D, GRAHAM K J, et al. Soil water content and organic carbon availability are major determinants of soil microbial community composition[J]. Microbial Ecology, 2004, 48(3): 424–430
Effect of fertilization managements on soil organic carbon and microbial community structure*
LI Qian1,2†, MA Kun2†, YE Xiuxiang1, YANG Jinjuan1, NIU Hongxia1, MA Ling1
(1. College of Agronomy, Ningxia University, Yinchuan 750021, China; 2. Breeding Base for State Key Laboratory of Land Degradation and Ecological Restoration in Northwest China, Ningxia University, Yinchuan 750021, China)
The Phospholipid Fatty Acids (PLFAs) fingerprint and humus fraction methods were used to determine the accumulation of soil organic carbon in cultivated farmlands and the interaction mechanisms of soil microbial communities with soil organic carbon. The combined effects of organic manure and chemical fertilizer on soil organic carbon accumulation, soil humus forms and their relationships with soil microbial community structures were studied in a 5-crop rotation mode of ‘potato-potato-oil sunflower-potato-oil sunflower’ over a period of five years. The experimental treatments were as follows — no fertilization, inorganic fertilizer application, combined application of inorganic fertilizer and cow dung, combined application of inorganic fertilizer and sheep manure, combined application of inorganic fertilizer and biological organic fertilizer, and then combined application of inorganic fertilizer and fulvic acid potassium. The results showed that soil organic carbon fluctuated with increasing tendency over the five-year period. Compared with the control (no fertilization), soil organic carbon increased at annual average rates of 6.61% and 8.97% under the treatments of combined application chemical fertilizer with cow dung or separately with sheep manure. The rate of accumulation of soil organic carbon was influenced by the amount and type of added exogenous organic matter to the soil. The contents of stable or tightly combined humus and ratio of loosely combined humus to tightly combined humus increased following the addition of high quantities of organic manure (chemical fertilizer plus cow dung or chemical fertilizer plus sheep manure) and inorganic fertilizer. Compared with no fertilization, there were significant differences in soil bacteria, fungi, actinomycetes, protozoa and total microbial biomass, marked by phospholipid fatty acids under combined application of inorganic fertilizer and sheep manure or cow dung. The biomass ratio of gram-positive bacteria to gram-negative bacteria (G+/G-) in the treatments with inorganic fertilizer plus organic fertilizers decreased. There was no obviously difference in the ratio of G+/G-among treatments of combined application of inorganic fertilizer and organic fertilizers. Multivariate analysis showed a good correlation between the first ordination axes based on soil microbial biomass marked by PLFAs and the second ordination based on combined soil organic carbon and humus (1= 0.568,2= 0.611). The relationship between soil microbial biomass and soil environmental factors was explained by the 98.69% cumulative variation in spatial scale. There was a positive correlation between the content of loosely combined humus and G+/G-of soil microbial community. It was concluded that the tightly combined humus fraction gained higher stability than the loosely combined humus when exogenous organic carbon was applied to the soil. The soil bacteria and fungi biomass marked by PLFAs were promoted with increasing amounts of exogenous organic carbon in the soil. The biomass ratio of fungi to bacteria was not influenced by fertilizer management and amount of exogenous organic carbon. Generally, the change in soil microbial community structure was driven by the change in soil organic carbon content and soil humus fraction. Combined inorganic fertilizer with cow manure and inorganic fertilizer with sheep manure supported the accumulation of soil organic carbon and the formation of loose combined humus, but also promoted an increase in soil microbial biomass. The research provided a reliable scientific basis for soil fertilization in semiarid areas in Ningxia.
Combined application of inorganic fertilizer and organic fertilizer; Soil organic carbon; Microbial community; Loosely combined humus; Tightly combined humus; Phospholipid fatty acid (PLFA)
LI Qian, E-mail: 940380226@qq.com; MA Kun, E-mail: makun0411@163.com
Dec. 25, 2017;
Jul. 20, 2018
S182; S154.3
A
1671-3990(2018)12-1866-10
10.13930/j.cnki.cjea.171190
2017-12-25
2018-07-20
* 宁夏科技支撑计划园区专项、国家自然科学基金项目(31660132, 31160104)和宁夏高等学校一流学科建设项目(NXYLX2017B05)资助
*This study was supported by the Technology Park Fund of Ningxia Science and Technology Support Plan, the National Natural Science Foundation of China (31660132, 31160104) and the First-Class Discipline Construction Project of Colleges and Universities in Ningxia (NXYLX2017B05).
† Equal contributors
† 同等贡献者: 李倩, 主要研究方向为农作制度理论与技术, E-mail: 940380226@qq.com; 马琨, 主要研究方向为农业生态学, E-mail: makun0411@163.com
李倩, 马琨, 冶秀香, 杨金娟, 牛红霞, 马玲. 不同培肥方式对土壤有机碳与微生物群落结构的影响[J]. 中国生态农业学报, 2018, 26(12): 1866-1875
LI Q, MA K, YE X X, YANG J J, NIU H X, MA L. Effect of fertilization managements on soil organic carbon and microbial community structure[J]. Chinese Journal of Eco-Agriculture, 2018, 26(12): 1866-1875
