三种巨大口蘑菌株贮藏期间褐变及相关酶活性
2018-11-27,,,,,,
,,, , ,,
(华南农业大学食品学院,广东广州 510642)
巨大口蘑(Tricholomagiganteum)是属于口蘑科(Tricholomataceae)口蘑属(Tricholoma)的一种珍稀食药用菌,主要分布在热带及亚热带地区[1]。该菌子实体营养丰富,所含必需氨基酸与非必需氨基酸比值(E/N)达到了WHO提出的理想蛋白质要求[2-3],营养价值优于杏鲍菇[4],且具有降血压、抑制病毒、抗肿瘤等功效[5-8]。
褐变是食用菌商品价值降低的关键因素,会使食用菌子实体色泽变深、软化腐烂和营养流失等[9-10]。酶促褐变是食用菌褐变的主要原因,由多酚氧化酶(PPO)催化酚类物质氧化而引起[11]。根据多酚氧化酶催化的底物不同,将其分为单酚氧化酶[酪氨酸酶(TYR)]、双酚氧化酶(儿茶酚氧化酶)和漆酶,而食用菌中的酶促褐变与PPO、TYR、漆酶活性和总酚含量密切相关[12],且酶活性因品种不同有所差异[13]。巨大口蘑作为一种高温型食用菌在我国南方已广泛栽培,其子实体具有一定的抗褐变能力[14]。但目前国内外对巨大口蘑的研究多集中在野生菌种分离驯化、覆土栽培、营养成分测定、与其他食用菌贮藏品质比较等领域,而对巨大口蘑不同菌株间的贮藏品质研究较少[15-17]。
本研究以巨大口蘑SCAU1、新3和新4三种菌株的子实体为试材,测定贮藏期间褐变度、PPO、TYR、漆酶和总酚含量等,分析褐变及生理指标间的相关性,为巨大口蘑的菌种选育及褐变内在机理的揭示提供依据。
1 材料与方法
1.1 材料与仪器
巨大口蘑SCAU1、新3和新4菌株 由华南农业大学食品学院应用真菌实验室提供;邻苯二酚 分析纯,天津市福晨化学试剂厂;邻联甲苯胺 分析纯,上海晶体试剂有限公司;L-多巴 化学纯,上海源叶生物科技有限公司。
PHSJ-4A型实验室pH计 上海精密仪器仪表有限公司;752型紫外可见分光光度计 上海精密仪器仪表有限公司;CR-410全自动色差计 日本Konica Minolta公司;GL21低温冷冻离心机 德国Eppendorf公司。
1.2 实验方法
1.2.1 样品处理 挑选新鲜采摘、无损伤、大小一致的三种巨大口蘑菌株子实体第一潮菇,用PE气调保鲜袋封口包装,置于12 ℃、相对湿度95%的生化培养箱中贮藏,定期取三种菌株子实体菌盖组织切片,测定各项生理指标,每组处理设3个重复。
1.2.2 测定指标及方法
1.2.2.1 褐变度 参照魏要武等[18]的方法,用标准陶瓷板(X=80.00,Y=83.00,Z=90.00)作为工作指标,测量蘑菇子实体菌盖组织切面的褐变度,用ΔL表示。ΔL值越小,表示颜色越白,褐变程度越轻。计算公式为:
褐变度(%)=(贮藏前L值-贮藏后L值)/贮藏前L值×100
1.2.2.2 PPO活性 参照王治江等[19]的方法,以每分钟OD420减少0.1为一个酶活力单位,结果以U/g FW表示。
1.2.2.3 TYR活性 参照杨春敏等[12]的方法。酶活力单位(U)定义为:20 ℃下每分钟引起OD值变化0.1为1个酶活力单位。
1.2.2.4 漆酶活性 参考潘迎捷等[20]的方法,并加以修改。取1 g子实体菌盖组织切片于研钵,加入5 mL预冷的0.05 mol/L磷酸缓冲液(pH6.8),冰浴研磨,然后在4 ℃,4000 r/min下离心10 min,上清液即为漆酶粗提液。漆酶活性测定体系为:0.5 mL 3.36 mmol/L邻联甲苯胺,3.0 mL 0.1 mol/L醋酸缓冲液(pH4.6)和0.5 mL酶液,25 ℃反应10 min。酶活力以600 nm处吸光率的增值表示。一个酶活单位(U)定义为在上述条件下,每分钟引起OD值改变0.01所需的酶量。
1.2.2.5 总酚含量 参照刘丽丹[21]的方法,以焦性没食子酸为标准品,绘制标准曲线,曲线方程为Y=2.5271X-0.0019,R2=0.9983,根据标准曲线计算样品中总酚的含量。
1.3 数据统计
采用DPS v9.5软件统计数据,方差分析采用ANOVA,多重比较的显著性检验采用Duncan新复极差法;应用SPSS 19.0统计软件进行偏相关分析和逐步回归分析。
2 结果与分析
2.1 不同巨大口蘑菌株的褐变度变化分析
褐变度是浅色食用菌在贮藏期间品质劣变最明显的指标之一,与其衰老程度和经济价值密切相关。如图1所示,三种菌株口蘑子实体的褐变度均随贮藏时间的延长而呈上升趋势。其中在前15 d内,三者间褐变度并无显著差异(p>0.05),随后,新3和新4两菌株口蘑褐变度上升的速率逐渐加快,且显著高于SCAU1菌株的褐变度(p<0.05)。贮藏30 d时,SCAU1、新3和新4三种菌株褐变度分别上升至0.68%、1.23%、1.09%,表明SCAU1菌株在贮藏期间相对不易褐变。随贮藏时间的延长,三种菌株的褐变度均呈不同速率的上升,这可能与贮藏期间酶促反应生成的类黑色素物质在子实体内逐渐积累有关[22]。
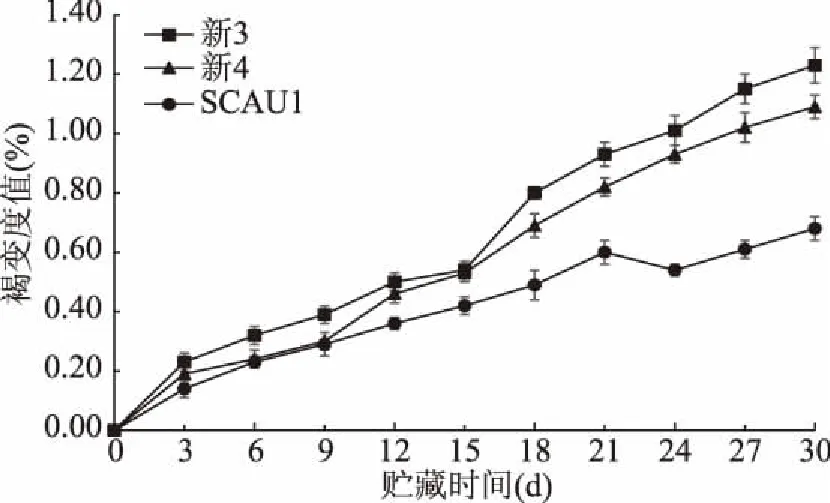
图1 不同巨大口蘑菌株的褐变度变化Fig.1 Changes in browning degree of different Tricholoma giganteum strains
2.2 不同巨大口蘑菌株的PPO、TYR和漆酶活性变化分析
PPO可将食用菌子实体中酚类物质氧化成醌,醌类物质聚合形成褐色物质导致子实体组织变色[23]。如图2所示,采摘时巨大口蘑SCAU1菌株的PPO活性显著低于另外两菌株(p<0.05)。在贮藏期内,三种菌株口蘑PPO活性整体呈下降趋势,可能是由于贮藏期间口蘑子实体自身的代谢作用和品质的劣变影响了酶的活性[24],李南羿等[25]对贮藏期间双孢蘑菇多酚氧化酶活性的研究具有相同趋势。在贮藏期间,新3和新4两菌株间PPO活性差异不显著(p>0.05),且二者PPO活性均显著高于SCAU1菌株(p<0.05),这与王泽生等[26]研究发现双孢蘑菇易褐变菌株的多酚氧化酶活性较高的结果相一致。
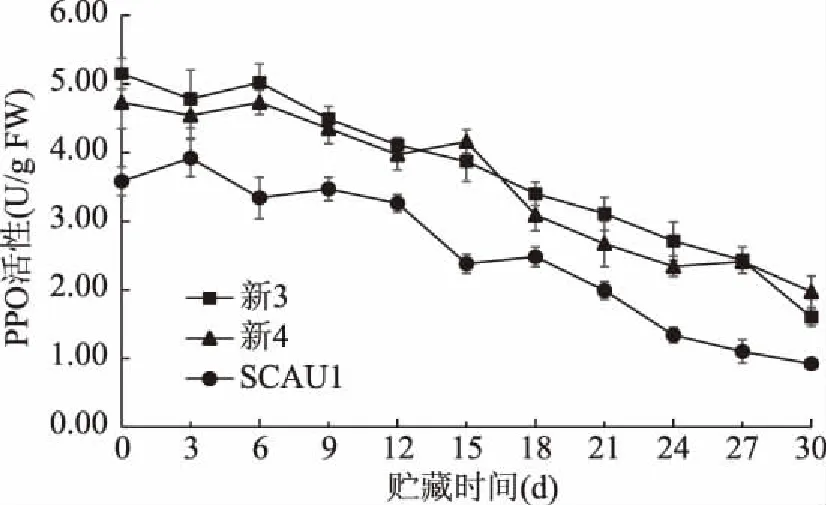
图2 不同巨大口蘑菌株的PPO活性变化Fig.2 Changes in PPO activity of different Tricholoma giganteum strains
TYR主要通过催化酪氨酸生成多巴,多巴进一步反应生成多巴醌,从而调控黑色素在食用菌子实体中的合成[27]。如图3所示,整个贮藏期内,巨大口蘑SCAU1菌株的TYR活性均低于另外两菌株,且与新3菌株TYR活性存在显著差异(p<0.05)。在贮藏期前18 d内,三菌株口蘑TYR活性波动较大,此后均呈不同速率下降;贮藏30 d时,SCAU1、新3和新4三种菌株TYR活性分别降至采收时的54.8%、74.4%和74.1%,表明SCAU1菌株的TYR活性偏低且降幅最大。三种菌株在贮藏期间酪氨酸酶均有先升后降的趋势,可能是采摘和贮藏期间诱导了酪氨酸酶的激活,使其酶活上升,贮藏后期由于子实体自身代谢作用品质劣变,酶活性逐渐下降[28]。在贮藏期间三种菌株的酪氨酸酶活有所差异,这与Moore等[29]对双孢蘑菇不同菌株酪氨酸酶活的研究结果一致。
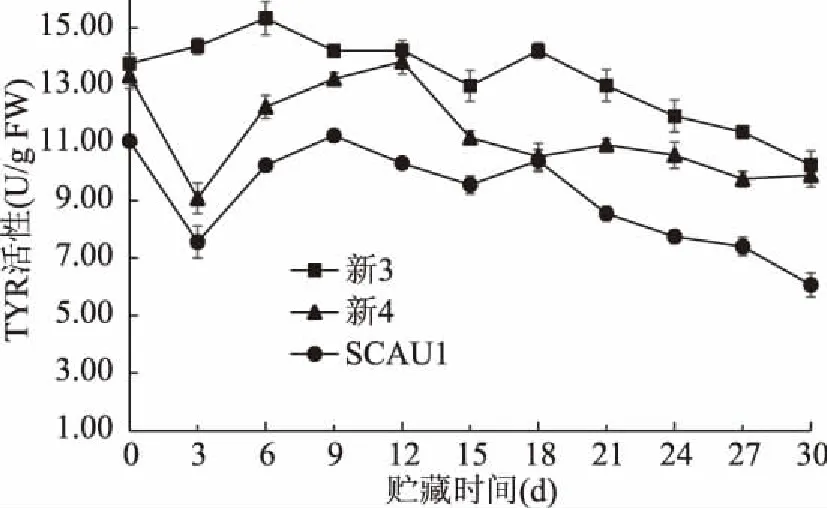
图3 不同巨大口蘑菌株的TYR活性变化Fig.3 Changes in TYR activity of different Tricholoma giganteum strains
漆酶可催化多种酚类和非酚类物质发生氧化或分解,影响食用菌贮藏期间的营养价值和品质[30]。如图4所示,SCAU1和新3两菌株漆酶活性在贮藏前期12 d内有较大波动,此后呈不同速率下降,在贮藏30 d时,分别下降31.2%、34.8%;整个贮藏期间,新4菌株的漆酶活性下降速率缓慢,且在24~30 d范围内,漆酶活性高于SCAU1和新3两菌株,贮藏30 d后,降幅仅为14.7%。这表明贮藏期间SCAU1和新3菌株漆酶活性降幅较大,新4菌株漆酶活性最为稳定。三种菌株在贮藏期间的漆酶活性并不一致,可能是由于不同菌株中的漆酶同工酶不同,催化氧化分解反应的作用存在差异,在白腐菌漆酶的研究中也出现同样的结果[31-32]。
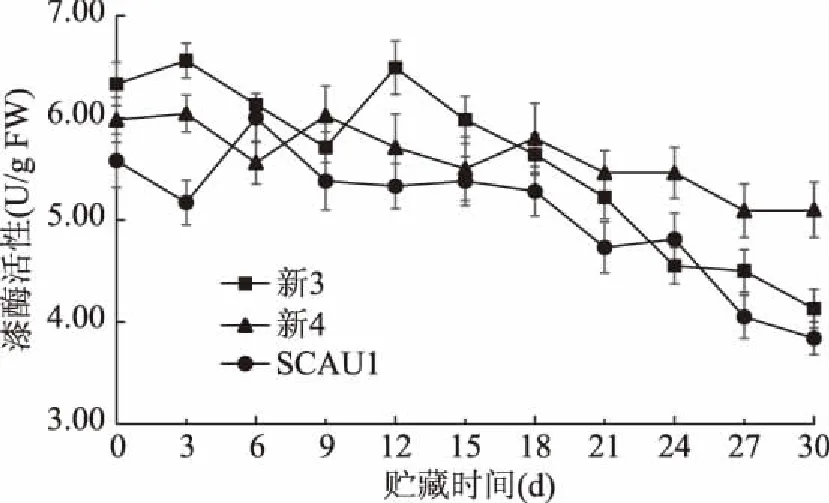
图4 不同巨大口蘑菌株的漆酶活性变化Fig.4 Changes in laccase activity of different Tricholoma giganteum strains
2.3 不同巨大口蘑菌株的总酚含量变化分析
总酚是酶促褐变反应的重要底物,具有很强的抗氧化能力,其含量与果蔬的褐变程度有密切关系[33]。如图5所示,在整个贮藏期间,三种菌株的总酚含量均呈锯齿“M”型下降趋势,并出现两个峰值,刚采摘时,巨大口蘑三种菌株的总酚含量均迅速上升,并在3~6 d内达贮藏期最大值,随后在15~24 d内均出现第二个峰值。在整个贮藏期内,新3菌株的总酚含量相对偏低,三种菌株的总酚含量并未出现显著差异(p>0.05)。贮藏初期,三种菌株的总酚含量逐渐上升,这可能是由于采后低温贮藏时受到逆境胁迫,口蘑子实体细胞产生更多酚类物质,增加的酚类物质可参与组织的伤愈过程[34]。随贮藏时间延长,膜脂过氧化作用加剧,细胞膜逐渐破坏,使原本被隔离的酚类底物与酶类接触发生反应,总酚含量逐渐减少,这与猴头菇和香菇在采后贮藏期间总酚的变化趋势一致[35-36]。
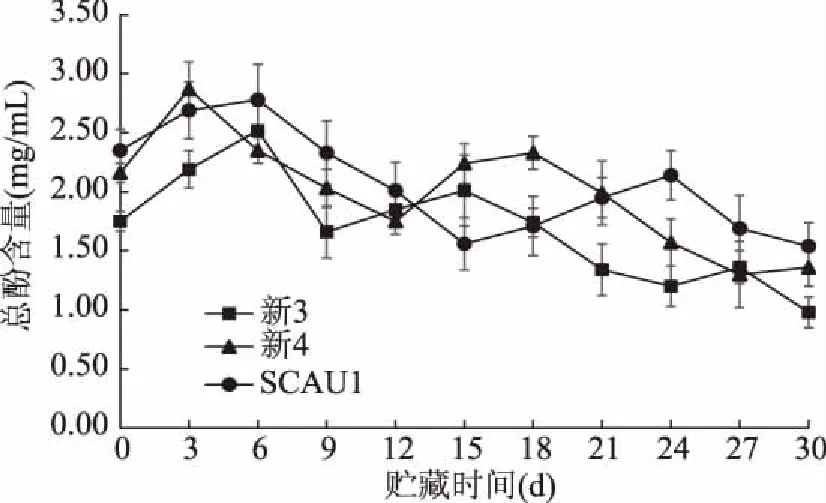
图5 不同巨大口蘑菌株的总酚含量变化Fig.5 Changes in total phenol content of different Tricholoma giganteum strains
2.4 不同巨大口蘑菌株褐变度与各生理指标的相关性研究
偏相关分析是在控制其他变量的线性影响下,分析两变量间的线性关系[37]。如表1所示,将巨大口蘑三个菌株贮藏期间的褐变度,分别与褐变相关酶及总酚含量进行偏相关分析,结果表明,新3菌株褐变度与PPO(p<0.01)和TYR(p<0.05)的活性有不同程度的显著负相关性,新4菌株褐变度与PPO活性呈极显著的负相关(p<0.01),SCAU1菌株褐变度与PPO活性显著负相关(p<0.05),三个菌株的褐变度与漆酶活性和反应底物总酚含量均无显著相关性(p>0.05)。
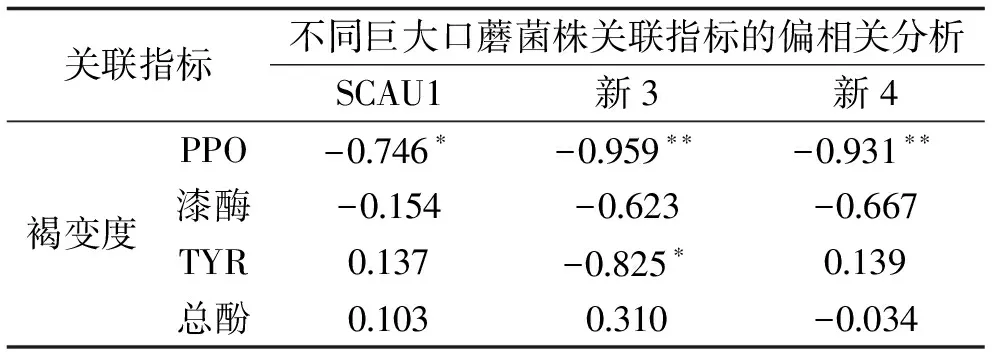
表1 不同巨大口蘑菌株褐变度与各生理指标的偏相关分析Table 1 Correlation analysis between browning degree and various factors in different strains of Tricholoma giganteum
逐步回归分析通过剔除与褐变不相关的因素,对相关因素做量化分析[38],进一步探讨各因素对不同菌株贮藏期间褐变度的影响。如表2所示,分别作出各因素与三个菌株褐变度之间的逐步回归分析,并建立逐步回归方程,对方程进行显著性检验,均达0.05显著水平,表明方程成立;调整决定系数R2均在0.8以上,表明方程的拟合性较好。逐步回归结果表明,三个菌株的褐变度均与PPO活性呈负相关,按照相关性大小排列依次为新3菌株>新4菌株>SCAU1菌株;除此之外,新3菌株褐变度还与其TYR活性呈负相关。
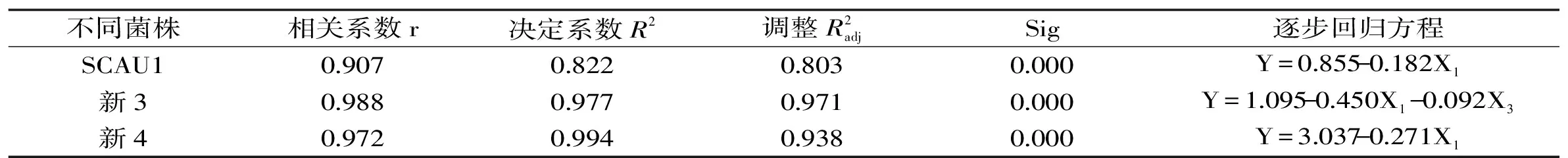
表2 不同巨大口蘑菌株褐变度与各因素的逐步回归分析Table 2 Stepwise regression analysis between browning degree and all factors in different strains of Tricholoma giganteum
3 讨论
巨大口蘑采后品质的劣变主要表现为色泽的加深,其原因与子实体褐变相关酶活性及反应底物密切相关[12]。贮藏期间三种菌株的褐变度均呈上升趋势,PPO和TYR活性总体呈下降趋势,且SCAU1菌株的相应指标均低于另外两菌株,说明低活性的PPO和TYR能降低SCAU1菌株的褐变度,这与王泽生等[26]对双孢菇的研究结果相一致。贮藏期间三种菌株的漆酶活性和总酚含量整体呈下降趋势但菌株间并未出现显著差异,说明漆酶和总酚并不是引起巨大口蘑三种菌株间褐变差异的主要原因,这与王乐芬等[39]研究双孢蘑菇褐变的结果相似。
偏相关性分析和逐步回归分析进一步表明三种菌株的褐变度均与PPO活性呈显著负相关,与贮藏期间双孢蘑菇、紫甘薯和茄子等研究中PPO的表现[26,40-41]基本一致。分析其原因,可能是因为在采前及贮藏初期三种菌株口蘑子实体的组织完好,未受损伤,子实体中PPO及其底物正常存在,由于多酚氧化酶及其底物定位的区域化使得酶促褐变相对不易发生[42],随着时间的延长,酶和底物有机会接触,酶促褐变反应程度逐渐增加,使得褐变度逐渐上升[43];但是,如果进一步延长贮藏时间,采后子实体自身的呼吸作用和能量的消耗以及细胞生理状态的劣变,可能导致PPO的蛋白结构受损或和酶的工作环境不适,进而引起酶活性逐渐下降[25],与褐变负相关,其中的具体机理复杂,还有待进一步的探索和研究。相关性分析中TYR活性仅与新3菌株的褐变度相关性显著,综合实验误差等因素,有待进一步增加试验样本的数量,以明确三种菌株中TYR活性与褐变度的关系。
4 结论
三种巨大口蘑菌株在采后贮藏期间,SCAU1菌株子实体褐变最为缓慢,新4菌株次之,新3菌株子实体褐变速度最快。相关性分析表明,新3、新4菌株的褐变度均与PPO活性呈极显著负相关(p<0.01),SCAU1菌株褐变度与PPO活性呈显著负相关(p<0.05),新3菌株的褐变度还与其TYR活性相关性显著,但各菌株与漆酶和总酚的相关性并不显著。在本研究中,巨大口蘑SCAU1、新3、新4三种菌株子实体在采后贮藏期间表现出不同的贮藏性能,综合各测定指标分析得出SCAU1菌株的贮藏品质最佳,新4菌株次之,新3菌株的贮藏品质最不理想。此研究结果可以为巨大口蘑在推广种植时,菌株品种的选育提供一定的指导。
