祁连山排露沟流域青海云杉种群结构与空间分布格局
2018-11-20赵维俊王顺利孟好军
赵维俊 ,王顺利 ,孟好军 ,马 剑 ,牛 赟
(1.甘肃省祁连山水源涵养林研究院,甘肃 张掖 734000;2.甘肃省森林生态与冻土水文水资源重点实验室,甘肃 张掖 734000;3.甘肃张掖生态科学研究院,甘肃 张掖734000;4.淮阴师范学院 城市与环境学院,江苏 淮安 223300)
种群垂直结构和径级结构反应了种群数量动态及其发展趋势,在一定程度上反映了种群与环境间的相互作用及其在群落的作用和地位[1]。种群空间分布格局描述了种群个体的空间分布特征,即使是同一种群在不同发育阶段、不同的生境条件也有明显的差别[2]。青海云杉Picea crassifolia林是我国寒区旱区典型的地带性植被类型,其生态功能特别是水源涵养功能对水资源短缺的西北内陆来说乃是重中之重,受气候变化特别是人为干扰的作用,祁连山生态环境受到了严重的破坏。近几年来,祁连山生态环境问题受到了党中央、国务院、环保部等的密切关注,并以坚决有力的举措扎实推进祁连山生态环境整治保护恢复工作。科学解决森林植被恢复与水资源间的矛盾,需要深入了解植被结构和空间格局对水文的影响,并能定量评价和准确区分植被的水文效应[3]。因此,作为森林生态恢复和生态水文研究的重要组成部分,其林木种群基础研究不可缺少。
目前,对祁连山青海云杉种群结构和空间分布格局已有一些专家和学者进行了研究[4-6],但对流域尺度海拔梯度的青海云杉种群特征研究较为少见,青海云杉林生长具有明显的海拔梯度变化特征,研究祁连山建群种或优势种青海云杉种群结构和空间分布格局,有助于了解当下青海云杉林海拔梯度上的变化特征和生长现状,获得其可能的发展趋势。另外,祁连山排露沟流域作为国家重点野外科学观测试验站,在该流域进行了大量的生态水文过程试验和研究[7-9],但多数研究还仅仅是在单一尺度或者均一地表覆被条件下进行研究,对主导过程的尺度演进关系还缺乏深入研究,譬如如何在流域海拔梯度进行生态水文过程综合试验研究,种群结构和分布格局可以为其植被水文试验研究提供依据。可见,本研究有望为祁连山青海云杉林生态系统的可持续经营、定量评价植被对水文的影响等方面提供依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于祁连山中段西水林区的排露沟流域(38°24′N,100°17′E),流域总面积 2.85 km2,长4.25 km,纵坡比降1∶4.2,海拔2 600~3 800 m。年平均气温-0.6~2.0 ℃,年蒸发量1 052 mm,年日照时数1 893 h,日辐射总量110.28 kW·m-2,年平均相对湿度为60%[10-11],属高寒半干旱山地森林草原气候。流域自然条件复杂,水热条件差异大,形成了多种具有明显垂直梯度的植被类型和土壤类型,海拔从低到高,植被类型依次为山地草原植被、山地森林草原植被、亚高山灌丛草甸植被、高山冰雪植被;土壤类型依次为山地栗钙土、山地灰褐土、亚高山灌丛草甸土、高山寒漠土。在各类土壤中山地灰褐土和亚高山灌丛草甸土是生长森林的土壤,山地灰褐土分布在海拔2 600~3 300 m地带,是乔木林的主要分布带;亚高山灌丛草甸土分布在海拔3 300~3 800 m 亚高山地带,是湿性灌木林的主要分布带。建群种青海云杉呈斑块状或条状分布在实验区海拔2 600~3 300 m 阴坡和半阴坡地带,与阳坡草地犬齿交错分布;灌木优势种有金露梅Potentilla fruticasa、鬼箭锦鸡儿Caragana jubata和吉拉柳Salix gilashanica等,草本主要有珠牙蓼Polygonum viviparum、黑穗苔Carex atrata和针茅Stipaspp.等。流域所在的祁连山西水林区曾在1935年发生过大规模的森林采伐事件[12]。
1.2 样地布设与植被调查
2017年7月份在祁连山排露沟流域青海云杉林分布的海拔范围2 700~3 300 m内沿海拔梯度选择生长状况良好,未受明显干扰的典型林分设置固定样地进行青海云杉林调查和取样,样地大小为20 m×20 m,将每个20 m×20 m样方分隔为16个5 m×5 m的样格,然后按“N”字字形顺序调查。调查对象为样方内所有胸径(DBH)≥ 1 cm的活立木木本植物,对每个个体悬挂铝制标牌进行标记,以待再次复查。DBH≥5 cm的个体用胸径尺测量其胸径,1 cm≤DBH<5 cm的个体则用数显游标卡尺测量,测量时随机选择相互垂直的两个方向测量两次,取平均值。调查内容包括活立木木本植物的胸径、树高、冠幅、枝下高、个体相对坐标(x,y),样地内物种组成、生长状况等,同时记录DBH<1.0 cm的活立木木本植物株数和高度及草本植物物种组成、生长状况等。结合张雷等人在该流域对青海云杉中龄林结构和生物量随海拔的变化特征[13-14],本研究采用以空间代替时间的研究方法,将排露沟流域青海云杉林分布划分为4个海拔区段,分别是海拔2 700~2 800 m(H1)、2 800~2 900 m(H2)、2 900~3 100 m(H3)和3 100~3 300 m(H4),调查样地基本信息见表1。根据青海云杉林的树高特征,划分为:下层木,H≤6 m,中层木,6 m<H≤12 m,上层木,H>12 m[14]。根据胸径将青海云杉非更新苗划分为:小树,5.0 cm<DBH≤12.5 cm,中树,12.5 cm<DBH≤22.5 cm,大树,DBH>22.5 cm[15]。
1.3 数据处理
测定种群的分布格局类型和集聚强度的数学模型较多,在实际应用中,各模型均有其适应性和优缺点。为避免不同模型的片面性,本研究采用如下多个指标进行测度,包括Caasier模型中的扩散系数、Davidh & Moore模型中的丛生指标、Kunou模型中的CA指标、Waters模型中的K指标、Morisita模型中的扩散指数、Lloyd模型中的拥挤度(M*)和M*/M指标、La模型中的La/M指标和Z-V模型中的Z/V指标,所有指标参数采用IBM SPSS Statistics 21软件统计分析,各模型的具体计算公式和判断标准见下[16-18]:

表1 不同海拔区段青海云杉林样地基本信息Table 1 The basic information in different elevations of Picea crassifolia forest
(1)Caasier模型的扩散系数C:
C=S2/M,其中:S2为样本方差,M为样本平均值,下同。当C=1时,种群为随机分布;C>1时,种群为聚集分布;C<1时,种群为均匀分布。
(2)Davidh & Moore模型中的丛生指标I:
I=S2/M-1,当I=0时,种群为随机分布;I>0时,种群为聚集分布;I<0时,种群为均匀分布。
(3)Kunou模型中的CA指标:
CA=(S2-M)/M2,当CA=0时,种群为随机分布;CA>0时,种群为聚集分布;CA<0时,种群为均匀分布。
(4)Waters模型中的K指标:
K=M2/(S2-M),K值越小,种群聚集度越大,K>8时,种群则接近Poisson分布。
(5)Morisita模型中的扩散指数I&:
I&=nΣ(fx2-N)/N*(N-1),其中:n为抽样数,N为总虫数。当I&=1时,种群为随机分布;I&>1时,种群为聚集分布;I&<1时,种群为均匀分布。
(6)Lloyd模型中的拥挤度M*和M*/M指标:
M*=M+S2/M-1,当M*/M=1时,种群为随机分布;M*/M>1时,种群为聚集分布M*/M<1时,种群为均匀分布。
(7)La模型中的La/M指标:
La=M-M/S2+1,当La/M=1时,种群为随机分布;La/M>1时,种群为聚集分布;La/M<1时,种群为均匀分布。
(8)Z-V模型中的Z/S2指标:
Z=S2/M-1+S2,当Z/S2=1时,种群为随机分布;Z/S2>1时,种群为聚集分布;Z/S2<1时,种群为均匀分布。
2 结果与分析
2.1 不同海拔区段青海云杉种群的垂直结构
分布在祁连山排露沟流域的唯一树种青海云杉林,其在不同海拔区段H1、H2、H3和H4青海云杉林分的郁闭度分别为0.66~0.69、0.58~0.87、0.50~0.83、0.35~0.66,海拔区段H1的郁闭度最大,H4的郁闭度最小,其他海拔区段林分郁闭度居中,这与样地林分密度直接相关(见表1)。不同海拔区段青海云杉种群高度级频率分布图见图1,从图1可以看出,不同海拔区段青海云杉种群高度级频率分布各不相同,海拔区段H1、H2、H3和H4青海云杉林分的高度级频率属于中层木所占的比例分别为:60.7%、45.2%、57.0%、52.5%,属于上层木和下层木所占的比例分别为:14.6%、11.2%、28.9%、32.5%和24.7%、43.6%、14.1%、15.0%,不同海拔区段林分中层木所占的比例均最大,需要指出的海拔区段H2下层木所占的比例接近中层木,海拔区段H3和H4下层木所占的比例最小、海拔区段H1上层木所占的比例最小。
2.2 不同海拔区段青海云杉种群的径级结构
采用空间代替时间的方法,即以胸径级代替年龄进行分析[19]。通过统计不同海拔区段青海云杉林种群内个体是小树以上的所有存活个体的胸径级频率分布,可以看出不同海拔区段青海云杉种群的胸径级频率分布存在着明显的差异(见图2),在所有存活个体当中,海拔区段从低到高的小树占各自的总体比例分别为44.8%、41.0%、52.1%和37.5%,中树占各自的总体比例分别为46.0%、50.5%、29.7%和38.8%,具有较多的小树和中树,二者所占总体比例均达到76.3%以上,大树个体较少;需要指出的是海拔区段H1和海拔H2区段大树所占的比例仅为9.2%和8.5%,海拔区段H3和海拔区段H4大径级个体所占的比例较海拔区段H1和海拔区段H2大树所占的比例大,分别为18.2%和23.7%,青海云杉林生长特别缓慢,低海拔处缺少大径级青海云杉个体,不排除历史上人为砍伐的原因。
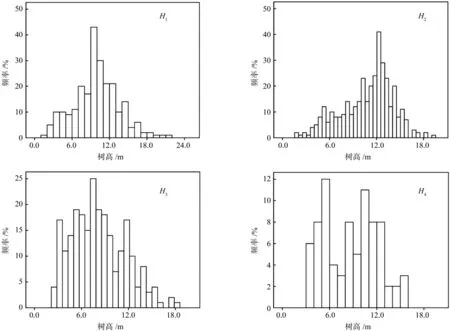
图1 不同海拔区段青海云杉种群样地高度级频率分布Fig.1 The height class frequency distribution in different elevations of Picea crassifolia population

图2 不同海拔区段青海云杉种群样地胸径级频率分布Fig.2 The DBH class frequency distribution in different elevations of Picea crassifolia population
2.3 不同海拔区段青海云杉种群的空间分布格局
用扩散系数(C)、丛生指标(I)、CA指标、K指标、扩散指数(I&)、拥挤度(M*)、聚集度(M*/M)、La/M指标和Z/S2等指标对不同海拔区段青海云杉种群空间分布格局进行了判定(见表2),从表2可以看出,不同指标均判定海拔区段H1、H3和H4青海云杉种群均呈聚集分布格局。当K指标趋于无穷大时(一般为8以上),则接近泊松分布,因此海拔区段H2青海云杉种群呈泊松分布格局,即:低海拔和高海拔青海云杉种群表现为聚集分布格局,中海拔青海云杉种群表现为泊松分布格局。不同海拔区段的拥挤强度(M*)指标大小表现为H4>H3>H1>H2,聚集强度(M*/M)指标大小表现为H3>H4>H1>H2,拥挤强度和聚集强度指标均表明海拔区段H1、H3、H4青海云杉种群的聚集分布特性和海拔区段H2青海云杉种群的泊松分布特性。因此,可能是该流域低海拔H1的水分限制和高海拔H3、H4的热量限制造成了青海云杉种群的聚集分布格局,其格局有利于抵抗不良环境的生态适应对策,中海拔H2的水分和温度的相对适宜造成了青海云杉林的泊松分布格局,因对资源需求及种内竞争程度减弱造成聚集强度的减弱。

表2 不同海拔区段青海云杉种群空间分布格局Table 2 Distribution pattern of Picea crassifolia population in different elevations
3 讨 论
(1)祁连山排露沟流域不同海拔区段的乔木唯一优势种是青海云杉林,这是由干旱和冷湿的环境决定的,其种群中层木所占的比例居中,可能是青海云杉种群林木中等高度、强烈保守的生活史策略导致该高度级生长慢、死亡率低的缘故[20]。海拔区段H2青海云杉种群下层木比例接近上层木所占的比例,林木更新较好。海拔区段H3和H4上层木所占的比例和海拔区段H1下木层所占比例均居中,原因是高海拔林木密度较低,竞争能力弱,加上处于林缘地段,光照条件较好,上层木生长得到一定程度的释放,而低海拔主要是人为干扰所致,当然加上此海拔段的水分匮乏的缘故。
(2)祁连山排露沟流域不同海拔区段的青海云杉种群结构试验表明,青海云杉种群小树和中树后续个体较多,大树较少,因是采伐后形成的天然次生林,种群处于恢复期阶段,虽然小径级个体较多,林木更新较好,但是基于样地尺度调查的基础上分析出的结论。目前,青海云杉林分布呈斑块化、片段化分布,林地生产能力和生态功能低下,森林天然更新恢复尤为困难,而且土地利用率不高[21-22],受多种因素的影响,祁连山的建群种青海云杉次生林天然更新能力差、更新苗难以成树,严重限制着其生态系统及多种服务功能的稳定性,青海云杉林生态系统一旦受到破坏,由于极少有其他树种能够适应这里的较差条件而更新起来,破坏的林分有可能向灌木化、草本化等方向发展。人工更新给造林恢复也带来很大的困难,如成活困难,造林成本高,造林后的结构和质量不好等问题,而且树木更新存在着树种间及地域间的巨大差异,以往的研究成果并不能被照搬应用到祁连山的青海云杉林管理中。因此,急需进行青海云杉次生林天然更新机理研究,以提高林分的生产能力和生态功能为目的,紧密结合天然林保护工程,提出有效的更新恢复技术措施,为营建生态效益好、社会效益高的林分结构模式提供坚实的理论支持,为改善祁连山生态环境、促进该区社会经济的可持续发展具有十分重要的意义。
(3)祁连山排露沟不同海拔区段的青海云杉种群空间分布主要符合聚集分布格局和泊松分布格局。对山地森林而言,海拔梯度的变化直接影响了水热组合,进而影响了林木在不同海拔的空间分布格局。研究表明,该流域降水随海拔梯度(2 600~3 300 m)的增加呈递增趋势,海拔每升高100 m,其降水平均递增率为11.09%;土壤表层温度随海拔梯度(2 700~3 100 m)的增加呈递减趋势,每升高100 m,其温度降低0.39 ℃[23],另外该流域青海云杉种群人为干扰很小。可见,影响海拔H1、H3和H4的聚集分布格局主要因子气候因子,而影响海拔H2的泊松分布格局主要因子可能是树木自身生物学特性决定的。不同林木种群分布格局其影响因子是有区别的,诸如恩施天然红椿种群的聚集分布格局主要来源于人为干扰[24]。关于林木种群格局研究,如果能结合林木径向生长,则有利于深入理解当地林木种群分布与外部环境和自身遗传因素的相互作用[25]。另外,格局强度指数难以反映格局类型随尺度变化的依赖关系,因此在后期的研究中沿海拔梯度建立样带进行青海云杉种群空间分布格局随尺度变化的空间格局及空间关联性分析,深入地认识青海云杉种群空间格局形成的起因与过程,更好地为当地森林经营提供依据。
4 结 论
(1)祁连山排露沟流域青海云杉种群垂直结构表明,不同海拔区段青海云杉种群中层木最多,海拔2 900~3 100 m和3 100~3 300 m上层木居中,海拔2 700~2 800 m下层木居中。海拔2 700~2 800 m和3 100~3 300 m灌木盖度和高度明显大于其他海拔,其草地盖度也明显大于其他海拔,草地高度在不同海拔区段间的变化不明显。
(2)祁连山排露沟流域不同海拔青海云杉种群径级结构表明,试验样地的青海云杉种群处于恢复期阶段。目前,整体而言,祁连山青海云杉林生态系统需要加强青海云杉林的管护和经营,特别是林木更新研究。
(3)祁连山排露沟流域海拔2 700~2 800 m、2 900~3 100 m和3 100~3 300 m的青海云杉种群呈聚集分布格局,其分布格局可能受制于气候因子;而海拔2 800~2 900 m的青海云杉种群呈泊松分布格局,其分布格局可能受制于自身的生物学特性。
