中国肉桂与越南清化肉桂叶解剖结构对比分析
2018-11-20梁晓静梁海英梁文汇李开祥
梁晓静 ,梁海英 ,梁文汇 ,李开祥
(1.广西壮族自治区林业科学研究院 a.八角肉桂工程中心;b.广西特色经济林培育与利用重点实验室;c.广西木本香料工程技术研究中心,广西 南宁 530002;2.克莱姆森大学,南卡罗来纳州 克莱姆森 29634)
肉桂C.cassia在我国已有2 000多年的栽培历史,主要分布在北回归线附近地区[1],广西和广东是肉桂的主产区,其中广西的分布面积最广,约占全国的50%以上。此外,福建、海南、云南和四川等省区也有少量栽培[2]。国内种植的当家品种为中国肉桂,经过长期的人工栽培和自然选择,已经形成了不同品种和品系,在广西,肉桂按产地分为“东兴桂”和“西江桂”[3]。西江桂主要分布在藤县、平南、桂平、岑溪、容县、苍梧、梧州等地,而东兴桂主要分布在防城、上思、大新和龙州等地[4]。
清化肉桂C.cassiavar.macrophyll产自越南清化省,亦称西贡桂,清化肉桂桂皮品质好,在国际市场上享有较高荣誉,Dao在过去20多年里研究越南樟属植物,认为所谓的清化桂其实与中国肉桂是同一个种,但是由于收获和加工工艺的不同,越南清化桂和中国肉桂的产品在外表上有着很大的差别[5]。邢继真对清化肉桂和中国肉桂桂皮的显微结构进行了对比分析,结果表明,两者的显微结构部基本一致,但清化肉桂的石细胞和木纤维较少,细胞内含红棕色物质多[6]。傅火带研究比较了清化肉桂和中国肉桂挥发油的差异,认为清化肉桂辣味小,叶油的折光率、含醛量高,成分有所不同,认为是两个不同的种[7]。
植物的叶片是进行光合作用和蒸腾作用的主要器官[8],其形态结构在一定程度上说明其对环境的适应性和进化机制,以及种源地环境对植物形态结构的影响程度[9]。不少学者应用叶片的解剖特征作为分析亲缘关系的佐证,邱芬等研究了不同倍性滇杨叶解剖结构特征,结果表明,对滇杨进行加倍处理之后,其叶片解剖结构与二倍体对照相比较发生了显著变化[10]。招礼军等对5个种源鹅掌楸叶解剖性状进行分析,发现其解剖性状在种间和种内存在着丰富的变异[11]。
目前对于不同品种肉桂的研究主要包括:挥发油成分[12]、叶形态多样性[13]、遗传物质多样性[14]、木材结构特性[15]等方面,而叶片解剖结构比较尚未见报道。本研究通过对不同品种肉桂叶片结构的显微观察,探索相同生态和栽培条件下,不同肉桂品种叶片解剖结构的差异,旨在为品种分类和肉桂资源的研究利用提供相关依据。
1 材料与方法
1.1 试验地概况
位于广西南宁市北郊,108°21′E,22°56′N,海拔95 m,属亚热带季风气候区,光照充足,年均日照时数为1 600 h~1 800 h;雨量丰沛,年降水量1 347.2 mm;空气湿润,年平均相对湿度为80%;气候温和,年均气温在21.6 ℃左右,≥10 ℃年有效积温在7 200 ℃,极端最低温-1.5 ℃,极端最高温度39.4 ℃,一般年份有2~4 d轻微霜冻;土壤是由沙页岩发育而成的赤红壤,pH值为4.5~6.0之间。
1.2 试验材料
供试的肉桂品种为越南清化肉桂和中国西江肉桂的优良单株家系,为V-1,V-2,C-1、2、3、4、5、6、7、8、11号等11个家系,2006年,分别于越南清化省和广西平南县筛选的优良单株并采集成熟种子,进行实生苗培育,2007年春季,将一年生不同优良单株家系苗木定植于广西林科院肉桂资源圃,株行距为2 m×2 m,定植后按常规方法管理。
1.3 测定方法
每个样本选择3株,选取生长一致,健康无病虫害的、向阳的叶片,连中脉一起剪成1.0 cm×0.5 cm的小块,立即放入FAA固定液中,固定24 h以上,再用不同梯度的酒精脱水,二甲苯透明,石蜡渗透包埋,用石蜡切片机切成4~10 μm厚的切片,脱水,番红-固绿染色,中性树胶封片,用Olympus光学显微镜观察。
1.4 数据分析
每个叶片选取3个横切面,每个切面选取3个视野,每个视野测量一组数据,观测指标包括叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度、栅海比、木质部厚度、韧皮部厚度、中脉厚度、中脉突起度。按照以下公式计算栅海比和叶片突起度。栅海比=栅栏组织厚度/海绵组织厚度,叶脉突起度(MPD) = 中脉横切面厚度/叶片横切面厚度。所有数据均用SPSS 13.0进行多重比较、主成分分析以及聚类分析。
2 结果与分析
2.1 叶片横切面解剖结构比较
如图1所示,中国肉桂和越南肉桂的横切结构均为典型的异面叶,由表皮、叶肉和叶脉三部分组成,上表皮细胞近方形,排列紧密,一层,厚度13.56~17.37 μm,下表皮为一层近圆形细胞,排列不规则,体积较上表皮细胞大,厚度为15.49~19.12 μm。肉桂叶肉细胞分化为栅栏组织和海绵组织,栅栏组织由1~2层柱状细胞组成,排列较为疏松,厚度为52.06~56.62 μm,海绵组织细胞分布十分疏松,厚度为122.4~174.95 μm,栅海比为0.30~0.43 μm。叶脉为离基三出脉,中脉较粗,厚度为820.15~1 110.74 μm,其上表皮细胞与维管束之间有3~5层厚角组织,下表皮内有6~8层排列疏松、体积较大的薄壁细胞,维管束为外韧维管束,横切面呈扇形,维管束近轴面为发达的木质部,木质部厚度为136.05~238.36 μm,由多列径向排列的导管和木射线,远轴面为韧皮部,韧皮部厚度为64.47~88.17 μm。韧皮部外为多层薄壁细胞组成的维管束鞘。

图1 不同品种肉桂叶片解剖结构Fig.1 Leaf anatomical structure of different varieties of C.cassia
由表1不同品种肉桂叶横切面解剖特征多重比较可知,清化肉桂2个家系和中国肉桂9个家系的叶片厚度、上表皮厚度、海绵组织厚度、栅海比、木质部厚度、中脉厚度之间存在显著性差异,而下表皮厚度、栅栏组织厚度及中脉突起度之间差异不显著。清化肉桂的叶片厚度、海绵组织厚度、木质部厚度及中脉厚度的平均值分别为251.69、170.09、234.49、1 077.29 μm 显著大于中国肉桂家系,中国肉桂家系叶片厚度、海绵组织厚度、木质部厚度及中脉厚度的平均值分别为219.35、132.46、164.92、868.59 μm。清化肉桂的栅/海为0.31,中国肉桂栅/海为0.41,显著小于中国肉桂。
2.2 肉桂品种各表型性状的相关性分析
肉桂叶解剖结构10个指标的相关性如表2所示,两两指标之间相关性最高的是叶片厚度和海绵组织厚度,相关系数达0.976,其次是海绵组织厚度与中脉厚度,相关系数达0.969,叶片厚度与木质部厚度、中脉厚度之间也呈极显著正相关(均为r=0.947),木质部厚度与海绵组织厚度、中脉厚度之间呈极显著相关性,相关系数分别为0.942和0.930。负相关系数最大的海绵组织与栅/海之间,相关系数为-0.949,其次是中脉厚度与栅/海之间,相关系数为-0.904。

表1 不同品种肉桂叶横切面解剖特征多重比较Table 1 Multiple comparison of anatomical features of transverse section of different varieties of C.cassia

表2 肉桂叶片解剖结构指标的相关性分析Table 2 Correlation analysis of anatomical structure index of leaves
2.3 不同品种肉桂叶片解剖结构特征的主成分分析
肉桂叶解剖结构的主成分分析如表3所示,前2个主成分的贡献率分别为63.290%、17.378%,累积贡献率达80.668%,说明这两个主成分概括了10个解剖结构指标的主要信息。在第一主成分中,叶片厚度、栅栏组织厚度、海绵组织厚度、木质部厚度、中脉厚度以及中脉突起度等指标的特征向量均为正值,其中木质部厚度的载荷量最大,为0.982,其次是叶片厚度、中脉厚度以及海绵组织厚度,载荷量分别为0.979、0.973和0.963。第二主成分中,信息载荷量最大的是下表皮厚度,为0.819,其次是韧皮部厚度,载荷量为0.727,其他指标的信息载荷量均低于0.700。

表3 肉桂叶片解剖结构特征的主成分分析Table 3 Principal component analysis of anatomical features of C.cassia leaves
10个变量集中于两个个主成分中,以第一主成分特征向量为横坐标,第二主成分特征向量为纵坐标,两个个主成分相互垂直,构成一个平面,叶片解剖结构指标形成2个分布群,第一个分布群由木质部厚度、叶片厚度、中脉厚度、海绵组织厚度、中脉突起度等5个指标组成,第二个分布区群由栅栏组织厚度、韧皮部厚度、上表皮厚度、下表皮厚度和栅栏/海绵组织厚度等5个指标组成(图2所示)。

图2 旋转因子载荷分布图像Fig.2 Load distribution diagram of rotation factor
2.4 基于叶片横切面解剖特征的不同品种肉桂聚类分析
根据叶片解剖结构10个指标对肉桂的11个家系进行聚类分析,结果见图3,当截取值约为4时,11个家系分成两组,V-1和V-2聚为一类,而中国肉桂9个家系聚为一大类,其中第二大类又分为两个亚类,第一亚类为C-1和C-5,第二亚类为 C-2、C-3、C-4、C-6、C-7、C-8、C-11, 越 南清化肉桂2个家系与中国肉桂9个家系的遗传距离最大,而中国肉桂的各个家系之间的截取值较小,这说明,越南清化桂与中国肉桂叶片解剖结构之间存在明显的差异。
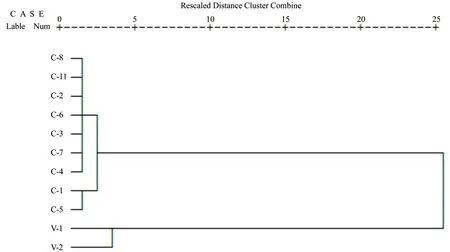
图3 基于叶片横切面解剖特征的不同品种肉桂聚类图Fig.3 Cluster maps based on the anatomical characteristics of the transverse section
3 结论与讨论
越南清化肉桂的桂皮又称南玉桂,品质优于中国肉桂,国际市场上,清化肉桂桂皮价格远远高于中国肉桂[4],享有较高声誉。有些学者认为清化桂是中国肉桂的大叶变种,也有研究认为清化桂和中国肉桂是同一个种,产品和外表上的差别是收获方式和加工工艺不同所造成[5]。
为了探讨中国肉桂和越南清化肉桂的亲缘关系,运用数量分析法对不同品种肉桂家系间叶的解剖结构特征量进行分析,从而将不同种分开。有不少学者运用数量分析法对群体间特定解剖结构特征量进行分析,以将不同的种或品种分开,例如张永福等基于叶片解剖特征分析了3种无患子科果树的亲缘关系,得到三月红是荔枝的一个变种,龙荔是由龙眼进化到荔枝一个中间过渡物种的结论[16]。郑瑞等对61个油茶品种的叶片及中脉横切面解剖特征进行了观察,并对品种间的亲缘关系进行了初步研究并编制了分类检索表,研究结果说明:叶片横切面解剖特征可作为区分油茶品种的依据之一[17]。
植物叶片内部结构相对稳定,各组织厚度及比值等结构特征是植物遗传性的反映,可作为植物分类的依据[18]。叶解剖结构的差异是遗传因子和环境因子共同作用的结果[11],供试的2个肉桂品种11个家系来自同一时期,同一区域,出现的差异是植物叶片内部的遗传物质作用下形成的不同形态结构。中国肉桂和越南清化肉桂在叶的主要特征上表现出较高的一致性比如叶的轮廓,脉序,栅栏组织层和海绵组织层的分化程度,中脉结构的排列等方面。而在叶片厚度、海绵组织厚度、栅海比、木质部厚度、中脉厚度等方面存在显著的差异。
主成分分析中,前两个主成分累积贡献率达到80.668%,说明这两个主成分概括了10个性状指标的主要信息,可以反映不同品种肉桂在叶解剖结构上的差异[19]。第一主成分贡献率为63.290%,木质部厚度的载荷量最大,其次是叶片厚度,中脉厚度,海绵组织厚度,这四个指标信息载荷量最大,都达到0.900以上,是区分肉桂不同品种最主要的解剖结构。聚类分析中,越南清化肉桂2个家系与中国肉桂9个家系各聚为一类,且遗传距离最大,进一步说明清化桂和中国肉桂叶解剖结构之间存在明显的差异,是遗传距离较远的种,这与之前肉桂叶表型性状的研究结果一致[14],认为中国肉桂与越南清化肉桂是两种不同的肉桂品种。
前人对植物亲缘关系的鉴别经历了形态学、孢粉学、同工酶等研究方法,但这些研究方法受主观影响较大[20,21]。除了阐明不同亲缘物种共存的形态学和表型性状外,从植物生理生化、分子、蛋白质组和代谢组学水平上解析植物亲缘识别的机制将是下一阶段研究的重点[22]。肉桂品种分类仍然是按照形态指标为主,存在主观因素的影响,分类结果仍有争议,今后肉桂种质资源亲缘关系的研究将以分子标记以及基因组测序技术、指纹图谱构建、目标基因定位及其连锁图谱绘制等方面为主。为合理利用肉桂资源提供指导。
